
Abdullah A. Mahmod
Department of Biochemistry, King Saudi University P.O. Box 2455, Riyadh 1 1451, Saudi Arabia.
Journal of Medical Sciences
Year: 2001 | Volume: 1 | Issue: 4 | Page No.: 251-254







ABSTRACT
The Effect of mercury chloride (HgCl2) on the human plasma cholinesterase has been investigated. Hg+2 was very effective inhibitor producing 50% inhibition at 1.2 x 10‾6 M and nearly 100% inhibition at 0.05 x 10‾6 M. The study of initial velocity indicates a noncompetitive mechanism of inhibition by Hg+2 when butrylthiocholine (BTCh) is substrate. Inhibition shows a Ki of 0.25�M. But when ATCh is substrate the inhibition was uncompetitive. The secondary plots of inhibition with respect to ATCh indicates a dissociation constant (Ki) of 0.12�M. The inhibition was dependent on the substrate employed during the activity assay. In conclusion, the distinct behavior of butrylcholinesterase(BChE) for Hg+2 may reflect the Subtle differences in the amino acids which provide the environment when the ‘catalytic tried’ is located.
PDF Abstract XML References Citation
How to cite this article
Abdullah A. Mahmod, 2001. Investigation of the Reversible Inhibition of Butrylcholinesterase by Mercury
Chloride. Journal of Medical Sciences, 1: 251-254.
DOI: 10.3923/jms.2001.251.254
URL: https://scialert.net/abstract/?doi=jms.2001.251.254
DOI: 10.3923/jms.2001.251.254
URL: https://scialert.net/abstract/?doi=jms.2001.251.254
INTRODUCTION
Human serum butrylcholinesterase (BChE), is a tetrameric glycoprotein of identical sub-units has 53% sequence identity to acetylcholinesterase (AChE) and shows similar responses to classical cholinesterase inhibitors (Lockridge, 1988).
Unlike AChE, the physiological significant of BChE is not fully understood. Role of BChE in cell differentiation, mitosis (Masson et al., 1996) and in detoxification (Chatonnet and Lockridge, 1989) have been proposed. BChE is studied because of its clinical usefulness in predicting prolonged apnea in response to the muscle relaxant succinylcholine. People with genetic variants of cholinesterase have an abnormal response to succinylcholine, (Kalow and Gunn, 1957) being unable to breath four hours after receiving a dose of succinylcholine that produce 3-10 min of apnea in the majority of the population. A second clinical use of BChE is in diagnosis of poisoning by organophosphate esters and carbamate of the type used in insecticides, in certain therapeutic drugs, and in nerve gas (Mounter et al., 1957).
Mercury is one of the most harmful metals found in the environmental contaminations and the most severe and unfortunate example is the so called minamata disease (Yamazaki et al., 1978). The toxicity of Hg+2 is assumed to be involved with its high affinity to–SH groups of protein or amino acid in living organisms (Baes and Mesmer, 1976 and Kamikashi et al., 1974). Effect of Hg+2 on BChE activity have been reported, but the details are unclear (Abdelsalam, 1987 and Hill and Soares, 1989)
It has been shown that BChE is inhibited by Mg+2 (Kambam, 1988), Cd+2 (Abdullah, 2000) and Cu+2 (Abdullah, 2001). Therefore, The research work was conducted to investigate the inhibition of human BChE by Hg+2, moreover, may provide some insight in the nature of the catalytic mechanism of BChE and the kinetic actions of this cation on the enzyme.
MATERIALS AND METHODS
Butrylthiocholine iodide, acetylthiocholine iodide and (5,5- dithiobis-2-nitrobenzoic acid) [DTNB] were obtained from Sigma Chemical Co., (St. Luis, MO, USA ) and Mercury chloride (HgCl2) was from Merk (Germany). The other reagents used in this study were of analytical grade.
The 10ml venous samples were collected from the antecubital vein, using heparin as the anticoagulant, 5ml was used for routine hematology and the other 5ml for cholinesterase estimations. The latter was centrifuged for separation of plasma from red cells and then stored at – 5° C until analysis. (Lepage et al., 1985).
Kinetic procedure: BChE and Hg+2 were incubated at 25 °C in 100mM phosphate buffer, pH 7.4 containing 4ml DTNB in final volume of 5.5ml. The substrates (butrylthiocholine or acetylthiocholine) were used in small volumes to give a final concentration of 0.06mM and the absorption increase at 412nm was recorded with Pharmecia Spectophotomater LKB Ultra Spec III, according to the methods of Garry and routh, (1964) which is based upon the technique of Ellman et al. (1961). Assays were carried out in duplicate and BChE activity was expressed as micro moles of sulfhydryl groups liberated in 1min. from 1ml of plasma. Mercury chloride in concentrations from 0.006mM to 1mM. The reaction was started by addition of the substrate.
In the measurement, a blank tube containing all the compound except enzyme was used. So the spontaneous hydrolysis of BTCh or ATCh has been eliminated. The effect of Hg+2 on enzyme activity was determined, the kinetic data were analyzed graphically and the line of the best fit for each of double reciprocal plots and kinetics constants and their standard deviation were obtained using the GraFit computer program (Leatherbarrow and Grafit, 1992).
RESULTS AND DISCUSSION
The inhibitory effect of very low concentration of Hg+2 on BChE and the IC50 (inhibitor concentration that inhibit 50% of enzyme activity) was shown in Fig. 1.
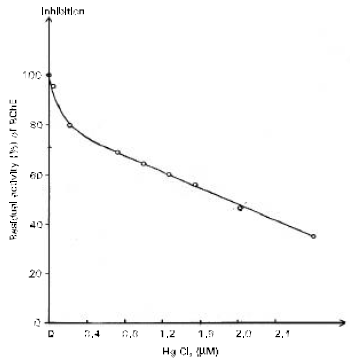 | |
| Fig. 1: | Mercury chloride (0.02-2.5μM) reversibly inhibited the BChE activity (27-80%) in a concentration dependent manner. The IC 50 [inhibitor concentration that inhibits 50% of enzyme activity] being about 0.88 μM. |
The results plotted according to the method of Lineweaver-Burk are shown in Fig. 2a. The inhibition was noncompetitive and Ki was 0.25FM. According to the description of this type of inhibition, an increase in substrate concentration causes both velocity(v) and initial velocity(vi) to increase by the same factor (Segel, 1975a). The net effect of a noncompetitive inhibitor is to make it seem as if less enzyme was present. The purely non competitive nature of the inhibition was confirmed by the secondary plot of Fig. 2, (i.e Fig. 2b). The slope of the reciprocal in the presence of a pure noncompetitive inhibitor is a linear function of [I]. In this inhibition the enzyme-substrate-inhibitor complex (ESI) may break down at a different velocity from the enzyme-substrate complex (ES), and the velocity is the sum of the two reactions (Segel, 1975a).
Results illustrated in (Fig. 3a) correspond to the double reciprocal plots according to the method of LineweaverBurk, of the values found when ATCh is the substrate. The inhibition was uncompetitive. The Ki, determined by reploting the slopes of the primary Lineweaver-Burk plot versus Hg+2 concentration (Fig. 3b) was 0.12FM.
A classical uncompetitive inhibitor is a compound that binds reversibly to the enzyme –substrate (ES) complex yielding an inactive ESI complex (Segel, 1975b). The inhibitor does not bind to the free enzyme. Consequently, we can predict that Maximum velocity (Vmax) in the presence of an uncompetitive inhibitor (Vmaxi) will be lower than the Vmax in the absence of inhibitor.
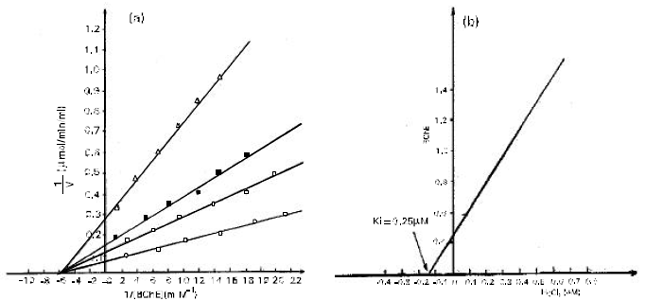 | |
| Fig. 2: | (a) Lineweaver-Burk plot of vs BTCh in the absence (○) and presence of 0.1 mM (⊗) 0.24mM (■) and 0.5 mM (∆)HgCl2). The lines were plotted from a regression analysis programme. Each point is the mean of value of four separate determination (b) Inset shows a secondary plot of the slopes derived from the primary linewever burk plot versus HgCl2 concentration. |
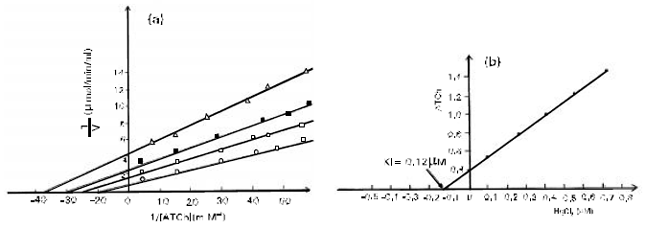 | |
| Fig. 3: | (a) Reciprocal plots in the absence of Hg (○) in the presence of 0.1 mM (⊗) 0.24mM (■) and 0.5 mM (∆) mercury chloride. The lines were plotted from a regression analysis programme. Each point is the mean of value of four separate determination (b) The inset shows a secondary plot of the slopes derived form the primary Linewver Burk plot versus HgCl2 concentration |
Unlike non-competitive inhibition, however, the apparent Michaelis-Menton constant (Km) value will decrease (Segel, 1975c). The type of inhibition appeared to be different for two substrates, there was a difference in affinity: the dissociation constant (Ki) for BTCh was about two times higher than for ATCh. This indicated a much higher affinity for the ATCh to both the peripheral and the active site. Though the underlying mechanism for the difference between the two substrates has not yet been clarified, the positional relation between the inhibitor site and substrate sites on the BChE might be different.
The calculated V max value was used to obtain nH (Hill coefficient from the Hill plot (Fig. 4) because the Hill plot deviates from linearity at low and high substrate concentrations, nH value was determined from the slope of the Hill plot at v =0.5 Vmax and was found to be as 1.1.
While some tight-binding inhibitors cause irreversible inhibition of an enzyme as a consequence of the formation of a covalently bonded enzyme-inhibitor complex (Morrison, 1969), it is not necessarily true that all tight-binding inhibitors must exert their effects in this manner. Reversible inhibition remains a possibility, and in this paper consideration is given to what kinetic effects might be with Hgcl2 that combine strongly with BChE in a reversible manner.
The possible mechanism is that Hg+2 binds with and peripheral anionic binding site of the free BChE in such a way that some conformed changes takes place which may influence the catalytic potency of BChE by the proper positioning of the esteratic and anionic subsites of the catalytic center for a cylation with ATCh (Balasubramanian and Bhanumathy, 1993; Bhanumathy and Balasubramanian, 1998; Sussman et al., 1991).
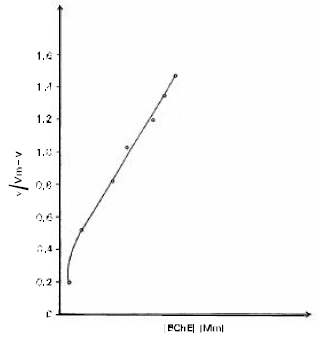 | |
| Fig. 4: | Hill plot for BChE nH values has been determined from the slop of plot at v=0.5 V max and found to be as 1.1. |
This possibility is similar to a non competitive type of inhibitory behavior of Hg+2.
The observation that catalytic activity of BChE can be modified by the binding of multivalent inorganic cations to site other than the catalytic site suggested a possible mechanism of Hg+2 neurotoxiaty (Marquis and Lerrick, 1982).
This data reported on the nature of the inhibition of Hg+2 on BChE and the IC50 showed that the mechanism of the inhibition differs according to the source of HgCl2. Indeed, we have observed that inhibition constants determined using old HgCl2 solution were significantly different from those with freshly prepared solution. We took the precaution of only reporting results determined using freshly prepared HgCl2 solution. The exact nature of inhibitory species in old HgCl2 solution as well as the mechanism of inhibition remain to be determined.
ACKNOWLEDGMENTS
The author is grate to H. A. Gambo for his technical assistance and the authority of King Khalid University Hospital for providing the blood. This work was supported by grand No. Bio/1416/03 from the research Center of the College of Science at King Saud University.
REFERENCES
- Abdelsalam, E.B., 1987. Comparative effect of certain organophosphorus compounds and other chemicals on whole blood, plasma and tissue cholinesterase activity in goats. Vet. Hum. Toxicol., 29: 146-148.
PubMedDirect Link - Balasubramanian, A.S. and C.D. Bhanumathy, 1993. Noncholinergic functions of cholinesterases. FASEB J., 7: 1354-1358.
Direct Link - Bhanumathy, C.D. and A.S. Balasubramanian, 1998. Selective inactivation of butyrylcholinesterase with metal chelators suggests there is more than one metal binding site. Int. J. Biochem. Cell. Biol., 30: 695-705.
CrossRefPubMedDirect Link - Chatonnet, A. and O. Lockridge, 1989. Comparison of butyrylcholinesterase and acetylcholinesterase. Biochem. J., 260: 625-634.
CrossRefPubMedDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Garry, P.J. and J.I. Routh, 1965. A micro method for serum cholinesterase. Clin. Chem., 11: 91-96.
PubMedDirect Link - Hill, E.F. and J.H. Soares Jr., 1987. Oral and intramuscular toxicity of inorganic and organic mercury chloride to growing quail. J. Toxicol. Environ. Health, 20: 105-116.
CrossRefPubMedDirect Link - Kalow, W. and D.R. Gunn, 1957. The relation between dose of succinylcholine and duration of apnea in man. J. Pharmacol. Exp. Ther., 120: 203-214.
PubMedDirect Link - Kamikashi, T., K. Kizaki, K. Murakami and S. Ishibashi, 1974. Difference in kinetic properties between hexokinase type I isoenzymes from various rat tissues with reference to the effect of a thiol inhibitor. Biochem. J., 137: 139-142.
Direct Link - Kambam, J.R., S.M. Perry, S. Enthman and B.E. Smith, 1988. Effect of magnesium on plasma cholinesterase activity. Am. J. Obstet. Gynecol., 159: 309-311.
PubMedDirect Link - Lepage, L., F. Schiele, R. Gueguen and G. Siest, 1985. Total cholinesterase in Plasma: Biological variations and reference limits. Clin. Chem., 31: 546-550.
PubMedDirect Link - Lockridge, O., 1988. Structure of human serum cholinesterase. Bioessays, 9: 125-128.
CrossRefPubMedDirect Link - Mounter, L.A., H.C. Alexander, K.D. Tuck and L.T.H. Dien, 1957. The pH dependence and dissociation constants of esterases and proteases treated with diisopropyl fluorophosphates. J. Biol. Chem., 226: 867-872.
Direct Link - Marquis, J.K. and A.J. Lerrick, 1982. Noncompetitive inhibition by aluminum, scandium and yttrium of acetylcholinesterase from Electrophorus electricus. Biochem. Pharmacol., 31: 1437-1440.
CrossRefPubMedDirect Link - Masson, P., M.T. Froment, C.F. Bartels and O. Lockridge, 1996. Asp70 in the peripheral anionic site of human butyrylcholinesterase. Eur. J. Biochem., 235: 36-48.
CrossRefDirect Link - Sussman, J.L., M. Harel, F. Frolow. C. Oefner, A. Tokler and I. Silman, 1991. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholine-binding protein. Science, 253: 872-879.
CrossRefPubMedDirect Link - Yamazaki, S., Y. Dokia, T. Watanabe and K. Fuwa, 1978. Problems in mercury determination in environmental samples. Ecotoxicol. Environ. Safety, 2: 1-8.
CrossRefDirect Link - Lineweaver, H. and D. Burk, 1934. The determination of enzyme dissociation constants. J. Am. Chem. Soc., 56: 658-666.
CrossRefDirect Link