
The Duc Nguyen
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Beatriz Estela Casareto
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
LiveDNA: 54.24973
Chitra Ramphul
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Keita Toyoda
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Toshiyuki Suzuki
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Taketomo Fujiwara
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Yoshimi Suzuki
Environment and Energy Systems, Graduate Schools of Science and Technology, Shizuoka University, Shizuoka, 422-8529, Japan
Research Journal of Microbiology
Year: 2018 | Volume: 13 | Issue: 3 | Page No.: 127-137







ABSTRACT
Background and Objective: Little is known about glycerol as a mediator between zooxanthellae and coral bacterial communities. This study evaluated the amount of glycerol in the coral holobiont of M. digitata and its effects as growth enhancer and antimicrobial properties of two Vibrio bacteria isolated from M. digitata. Materials and Methods: The amount of glycerol in tissue of M. digitata was determined after four days exposure to temperature (27, 32°C) and an extended period of light (12 and 18 h per day). Next, culture-independent and agar diffusion techniques were used to assess the antagonistic/antimicrobial properties of potential Vibrio bacteria isolated from M. digitata. Effect of glycerol on growth and antimicrobial properties of two selected Vibrio bacteria were examined by liquid culture assays. Results: Seventy-one isolated colonies from M. digitata were tested against the pathogen V. coralliilyticus SWA07, only six (8.5%) showed antimicrobial activity. Results indicated that isolates MH66 and MB3 displayed antibacterial and antagonistic activity against a wide range of target bacteria. So these were selected for further study. Two coral bacterial isolates MH66 and MB3 were closely related to Vibrio sp. displayed potent antagonistic/antimicrobial activity against target bacteria. Glycerol concentrations (from 0.5 to 10 mg mL–1) significantly enhanced the growth and antimicrobial properties of both MH66 and MB3. Conclusion: Glycerol concentration increased when M. digitata was exposed to light and temperature stresses and it enhanced the growth and antimicrobial properties of Vibrio MH66 and MB3.
PDF Abstract XML References Citation
How to cite this article
The Duc Nguyen, Beatriz Estela Casareto, Chitra Ramphul, Keita Toyoda, Toshiyuki Suzuki, Taketomo Fujiwara and Yoshimi Suzuki, 2018. Glycerol Enhances Growth and Antimicrobial Properties of Selected Vibrio Bacteria Associated with the Coral Montipora digitata. Research Journal of Microbiology, 13: 127-137.
URL: https://scialert.net/abstract/?doi=jm.2018.127.137
URL: https://scialert.net/abstract/?doi=jm.2018.127.137
INTRODUCTION
The coral holobiont is the association of the coral with several micro-organisms: the symbiotic algae zooxanthellae, bacteria, archaea, cyanobacteria, fungi, viruses and endolithic algae1-3. Zooxanthellae play important roles in the growth and survival of reef-building corals by providing substantial amount of carbon and energy to the corals4. Zooxanthellae synthesize organic carbon, such as glycerol and glucose, which are usually translocated up to 70% to the coral tissue5,6. Glycerol has historically been considered to be the most significant component of the translocated carbon from zooxanthellae7,8. However, recently there is increasing evidence that glycerol is not the main translocated carbon in intact symbiosis under the normal environment conditions9,10. Several previous studies have suggested that increasing the synthesis of glycerol from zooxanthellae is mainly related to an osmotic stress and photo-protective mechanism11-13. When corals were exposed to changes in environmental conditions (such as elevated seawater temperature, increased irradiation, changes in salinity and pollution), zooxanthellae might face hyper osmotic stress, which therefore could cause a higher concentration of glycerol in the coral tissues12. However, the roles of glycerol in coral holobiont are poorly understood. Coral-associated bacterial communities are the key components of the coral holobiont due to their crucial roles in the nutrient cycles and protective/defensive mechansism1,2,14. The complex antimicrobial and/or antagonistic properties of certain coral-associated bacteria could play vital roles in the regulation and structuring of coral-associated bacterial communities and hence could affect the coral health state1,15-18. Some studies have found that when corals are exposed to stressful conditions, such as high temperature, some changes in abundance and diversity of mainly Vibrio group in coral-associated bacteria communities occurred19,20. Many Vibrio are known as opportunistic coral pathogens under certain environmental conditions and their virulence depends on environmental stress conditions such as high temperature21-23. On the other hand, some culturable Vibrio bacteria isolated from coral colonies displayed some type of antimicrobial properties against certain pathogens15,16,18, whereas others act as antagonists to a range of other coral-associated bacteria16,17. These characteristics may assist the bacteria cells in competition for nutrients, space and survival24. Since the antimicrobial properties of some Vibrio associated-coral bacteria could promote competition with other bacteria in a certain way, under favorable conditions, these might increase their abundance24. Evidence from the current literature suggests that coral-associated Vibrio bacteria may act as important biological indicators under certain environmental changes20,23. To date, most of the published research on the antimicrobial or antagonistic activity of some coral-associated bacteria are focussed on its abundance and taxa15-18,25. Since glycerol could be one of the preferred carbon sources of bacteria, some bacteria might consume glycerol for their growth through different metabolic pathways26-29. In addition, glycerol may promote the synthesis of antibiotic compounds of some bacteria through secondary metabolic activity30,31. As a result, glycerol may affect the growth and metabolism of certain coral-associated bacteria. It is likely that the elevated concentration of glycerol in corals could expand the activities of certain Vibrio, which may trigger an increase in the abundance of Vibrio-associated coral. However, there are a few reports on the effects of glycerol on the growth and secondary metabolism of Vibrio associated-coral bacteria. As a result, the aim of this study was to determine the concentration of glycerol in tissue of the coral M. digitata under stressful conditions (high temperature; extended lighting period) and to detect certain Vibrio bacteria isolated from M. digitata which displayed potent antimicrobial and antagonistic activities. The final objective was to assess the effect of glycerol on the growth and antimicrobial properties of certain Vibrio bacteria isolated from M. digitata.
MATERIALS AND METHODS
Collection and treatment of samples: Healthy and bleached coral M. digitata (x3), six individuals of the Drupella spp. (Drupella) and one crown-of-thorns starfish (COTS) were collected in September 2017 at Sesoko reef (26°39 N, 127°51 E), Okinawa, Japan. Samples were rapidly prepared for isolation of bacteria by washing them twice with filtered autoclaved seawater (FASW). For incubation experiment, twelve branches of healthy M.digitata were kept under running natural seawater in aquaria (Tropical Biosphere Research Center, University of the Ryukyus, Sesoko Island, Okinawa) during 4 days for adaptation process. The collection of samples was conducted according to permission No. 28-6 granted from Okinawa Prefectural Government.
Determination of glycerol concentration in tissue of M. digitata: normal vs. stressful conditions of high temperature and extended lighting exposure Incubation design: The incubation experiment was set according to Suzuki et al.32. Three branches (~5 cm in length) were placed in each of four glass bottles containing 800 mL of 2μm filtered seawater (Advantec, CA, USA). The control/normal condition was set at 27°C and 12 h of lighting period/day; thermal stress was at 32°C, 12 h of lighting period/day; light stress at 27°C, 18 h of lighting period/day; thermal combined light stresses were set at 32°C and 18 h of lighting period/day.
Treatment of samples and measurements of glycerol: Coral samples (~1 cm in length) from the middle part of coral branches were collected (x3) before the start of the incubation (initial) and after 4 days of incubation. Their surface areas were determined using the aluminum foil method32. Coral samples were mashed in 3 mL buffer (50 mM Tris-HCl, pH 7.5) in a mortar, then centrifuged at 3.000 g for 2 min, the supernatants were then centrifuged at 10.000 g for 10 min. The extracts were stored at -80°C until futher analysis. Glycerol was measured according to the Glycerol Colorimetric Assay Kit manufacturer’s protocol (Cayman Chemical Ltd., Michigan, USA). The concentrations were normalized to the surface area of the coral pieces and were expressed in μg cm–2.
Isolation and growth of bacteria: Small portions (~1×1 mm) of tissue from the mouth of coral predators (Drupella and COTS) were cut by using sterile scissors and washed twice with FASW. Coral tissue was extracted according to Shiroyama et al.25. The resulting tissue was placed directly onto marine agar (MA) plates according to Ramphul et al.33. All the plates were incubated at 27°C for 48 h. Bacterial colonies were screened according to their size, shape and pigmentation34. A single colony of each pure bacterial isolates was transferred to 2 mL of liquid marine broth tryptone33 (MBT). The bacteria were grown for 18 h with shaking (100 rpm) at 27°C and then stored at -80°C with 20% of sterile glycerol (300μL) for subsequent studies.
Identification of bacteria through 16S rRNA sequencing analysis: One milliliter of bacterial culture (18 h) was centrifuged (10.000 g, 2 min, 20°C). The pellets were used for DNA extraction using a modified protocol from Dilhari et al.35. Polymerase Chain Reaction (PCR), according to the manufacturer’s protocol (No. R010A, TaKaRa Bio Inc., Shiga, Japan), together with the primers fD1 (5-AGAGTTTGATCCTGGCTCAG-3) and rP2 (5-ACGGCTACCTTGTTACGACTT-3)36 were used to amplify each sample. The size of the PCR products was verified on 1% agarose gel and was purified using a NucleoSpin Gel and PCR clean-up kit (Macherey-Nagel, Duren, Germany). 16S rRNA sequencing was carried out at the Research Institute of Green Science and Technology, Shizuoka University, Japan. Bacteria species were identified through the Basic Local Alignment Search Tool (BLAST) using DNA Data Bank of Japan (http://blast.ddbj.nig.ac.jp). Potential coral pathogens were then identified and used as target bacteria for screening antimicrobial properties of coral bacterial isolates.
Antimicrobial properties of coral-associated bacteria by agar-disk diffusion method: Coral bacterial isolates were initially screened for their antibacterial properties against the bacterium V. coralliilyticus SWA07 (AB 490821)33. Those isolates which inhibited the growth of V. coralliilyticus SWA07 (an inhibition zone >10 mm) were retained for the second screening. The latter consisted of using the coral bacterial isolates (positive results to the first screening) as test samples/bacteria against various target bacteria isolated from Drupella and COTS. Antagonistic activity of coral bacterial isolates (positive results to the first screening) were screened against other coral bacterial isolates. Amodified agar diffusion assay was employed according to Rypien et al.17. After 10 min, sterilized paper discs (ø = 8 mm, thick type, Advantec, Tokyo, Japan) were placed onto the soft agar lawn (9 paper discs/plate) and 20 μL of 18 h coral bacterial isolates were dripped onto the paper discs.Marine broth (20 μL) was used as negative control. All the plates were incubated at 27°Cfor 24 h. The diameter of the inhibition zone was measured. All the assays were repeated twice.
Effect of glycerol and its concentrations on the growth of coral bacterial isolates MH66 and MB3: Bacteria cultures (24-h)were diluted (OD600 = 0.02) and 100 μL were inoculated into the wells of sterile 96-well micro plates. Afterwards,100 μL of MBT medium containing various concentrations of sterile glycerol (Wako, Osaka, Japan) was added to obtain final glycerol concentrations of 0.05, 0.1, 0.25, 0.5, 1, 2.5, 5 or 10 mg mL–1. Medium without glycerol was used as control. Micro plates were incubated at 27°C with shaking (100 rpm) for 60 h. The OD600 was measured every hour for the first 24 h and then every 6 h until the end of incubation (60 h) using a BioTek SynergyTMHT Multi-Detection Microplate Reader. The growth curves of MH66 and MB3 were deduced both in the presence and absence of glycerol37. The mean of OD600 value at the stationary phase growth (24 h) in the presence of glycerol was compared to control. The experiment was repeated three times.
Preparation cell-free supernatant of coral bacterial isolates - MH66 and MB3: The cell-free supernatant (CFS)of MH66 and MB3 were collected under two different treatments (optimum glycerol concentration and optimum growth phase) and prepared according to Shnit-Orland et al.38. The first group of CFS was extracted through enrichment culture of MH66 and MB3 in MBT containing various glycerol concentrations (0, 0.05, 0.1, 0.25, 0.5, 1, 2.5, 5, 10 mg mL–1) at 27°C and shaken at 100 rpm and harvested after 48 h of incubation (representing the stationary phase). The second group of CFS was prepared through enrichment culture of MH66 and MB3 in MBT containing 1 mg mL–1 of glycerol (selected from the first assay) and without glycerol and harvested at different growth phases at 6, 12, 24, 36, 48, 60 h.
Antimicrobial assay using the CFS of MH66 and MB3: Antimicrobial assay of CFS was conducted in a 96-well plate according to Shnit-Orland et al.38. The first group of CFS was tested against the growth of three target bacteria (D11, D12 and V. coralliilyticus SWA07). The second CFS group was used to test growth inhibition of strain V. coralliilyticus SWA07 only. Briefly, a 1:1 ratio between the levels of treatment (100 μL of CFSs) and target strain cultures (100 μL diluted to OD600 = 0.02) were added in each well in triplicates. Filtered MBT was used as blanks (200 μL) and negative control (100 μL). Plates were incubated at 27°C and shaken at 100 rpm in a micro plate shaker. The OD600 value was read every hour for 12 h of incubation using a micro plate reader, the assay was repeated twice. The OD600 values at initial and final time points of log phase of target bacteria we reused to calculate the growth rate over a given time according to Stasiak-Rozanska et al.37. Inhibition percentage values were determined for each different CFS as described by Couch et al.39 following the equation:
where, I is the inhibition percentage, BC is the mean growth rate of target bacteria in the negative control and BE is the mean growth rate of target strain with treatment CFSs of MH66 and MB3.
Data analysis: One-way ANOVA and Tukey tests were used to determine the stress conditions on glycerol concentration in coral tissues and the effects of glycerol concentrations on the growth and antimicrobial properties of Vibrio bacteria. The relationship between glycerol concentrations (transformed into log form) and growth of MH66 and MB3 was detected using Pearson's correlation. All the statistical analyses were performed using MINITAB software version 14 and a p-value less than 0.05 was set as statistically significant to test for all the hypotheses: (I) healthy coral tissues from M. digitata will contain a higher concentration of glycerol under temperature and light stresses, (II) Vibrio bacteria isolated from M. digitata will show antimicrobial properties against certain possible coral pathogens and antagonism against other coral bacterial isolates, (III) glycerol will enhance the growth of certain Vibrio bacteria and (IV) glycerol will enhance the antimicrobial properties of certain Vibrio bacteria.
RESULTS
Glycerol concentration in coral tissues: The glycerol concentration in tissue of the healthy coral M. digitata after four days incubation varied according to different stress conditions as shown in Fig. 1 (One-way ANOVA, F = 56.64, p<0.0001). In thermal combined light stresses, glycerol concentration increased by 1.9 times (21.0±1.5 μg cm–2) with respect to the initial (10.9±0.6 μg cm–2, p<0.0001), also 1.2 times higher compared to light stress treatment (p<0.01). In light stress, glycerol concentration was 1.5 times higher (17.2±0.7 μg cm–2) than the initial (p<0.001). No significant difference was noted among control, thermal stress and initial (p>0.05). The hypothesis I was supported by the above results, which denoted an increase in temperature 32°C and 18 h extended light exposure increased the glycerol concentration in coral tissue of M.digitata.
Antimicrobial, antagonistic activity and identification of bacterial isolates: The bacterial isolates from Drupella, COTS and coral M. digitata which displayed antibacterial/ antagonistic effects were identified through 16S rRNA gene analysis and exhibited in Table 1.
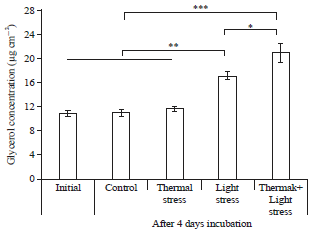 | |
| Fig. 1: | Glycerol concentration in healthy M. digitata tissues (μg cm–2) at initial and after 4 days exposed to thermal (32°C) and light stresses (18 h of illuminated period/day), values are the Mean±SD, n = 3. Significant differences among conditions were determined by Tukey HSD test (*p<0.01, **p<0.001 and ***p<0.0001) |
| Table 1: | 16S rRNA gene analysis of bacterial isolates from Drupella spp., COTS and coral M. digitata |
 | |
| #Sequences (>400 base pairs) were compared with existing sequences in the GenBank using DDBJ, *Potential coral pathogens from Drupella (D) and COTS (C), M. digitata healthy (MH), M. digitata bleached (MB) | |
| Table 2: | Antimicrobial properties of bacteria isolated from M. digitata |
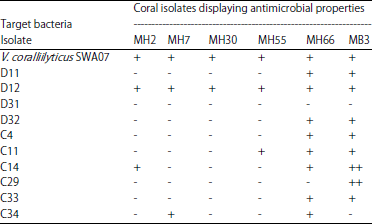 | |
| Bacterial isolates from Drupella (D) and COTS (C); M. digitata healthy (MH); M. digitata bleached (MB); +: Presence of antibacterial activity (inhibition zone diameter 10-12 mm); ++: Strong presence of antibacterial activity (inhibition zone diameter >12 mm), -: Absence of antibacterial activity or inhibition zone diameter <10 mm | |
Seventy-one isolated colonies from M. digitata were tested against the pathogen V. coralliilyticus SWA07 through the agar diffusion assay, only six (8.5%) showed antimicrobial activity (Table 2). The spectrum of the antibacterial activity of these isolates was then tested against other target bacteria isolated from Drupella and COTS was represented in Table 2. As shown in Table 3, antagonistic activity of six coral bacterial isolates was studied individually against several other coral bacterial isolates.
| Table 3: | Antagonistic activity among bacteria isolated from M. digitata |
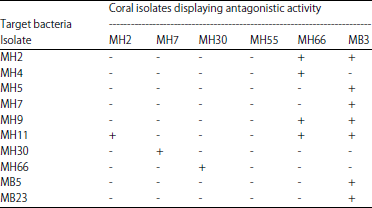 | |
| Bacterial isolates from M. digitata healthy (MH); M. digitata bleached (MB); +: Presence of antagonistic activity (inhibition zone diameter: 10-12 mm); -: absence of antagonistic activity | |
Results indicated that isolates MH66 and MB3 displayed antibacterial and antagonistic activity against a wide range of target bacteria. Isolates MH66 and MB3 identified as Vibrio bacteria were selected for further study (Table 1).
Effect of different glycerol concentrations on the growth of Vibrio MH66 and MB3: The isolates MH66 and MB3 showed slightly different growth patterns in various glycerol concentrations (0, 0.1, 1, 2.5 mg mL–1, Fig. 2). The lag phase, over a short period of 1 h was similar for MH66, MB3, control and different concentrations of glycerol. However, there was a considerable increase in the growth (exponential phase) for both isolates when grown at concentrations of 0.1, 1 and 2.5 mg mL–1.
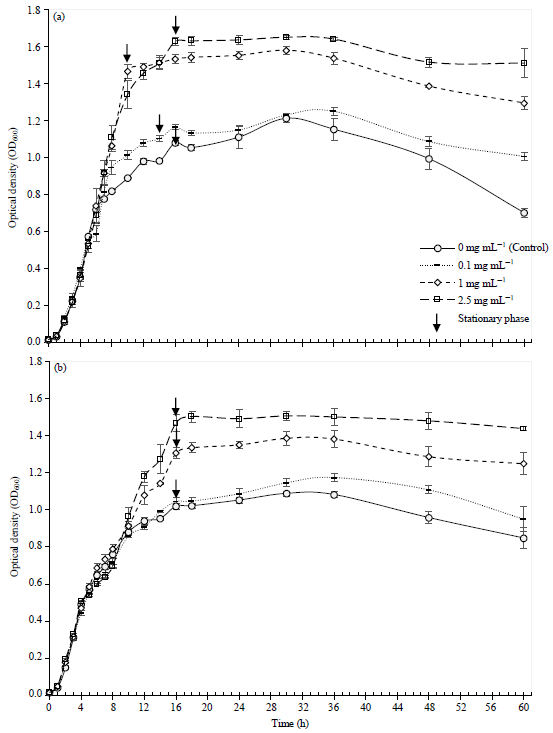 | |
| Fig. 2: | Effect of various glycerol concentrations (0, 0.1, 1 and 2.5 mg mL–1) on the growth of isolates (a) MH66 and (b) MB3 |
| Values are the Mean±SD, n = 3 | |
MH66 grown in concentrations of 2.5 mg mL–1 reached stationary phase at 16 h, cultures grown in glycerol concentrations of 1, 0.1 mg mL–1 and control reached stationary phases at 10, 14 and 16 h, respectively (Fig. 2a). However, MB3 grown in all glycerol concentrations (2.5, 1, 0.1, 0 mg mL–1) reached stationary phase at 16 h (Fig. 2b). A positive significant correlation between glycerol concentrations (0.05, 0.1, 0.25, 0.5, 1, 2.5, 5 and 10 mg mL–1) and growth of MH66 (r = 0.749, p<0.001) and MB3 (r = 0.962, p<0.001) was noted (Table 4). These results abided to the third hypothesis: glycerol enhanced the growth of Vibrio bacteria, MH66 and MB3, isolated from M. digitata.
Effect of glycerol on the antimicrobial properties of Vibrio MH66 and MB3: Effect of different glycerol concentrations at the stationary phase. The growth inhibition (%) against the three target bacteria (D11, D12 and V. coralliilyticus SWA07) varied significantly by enriched CFSs obtained from different concentrations of glycerol as shown in Fig. 3.
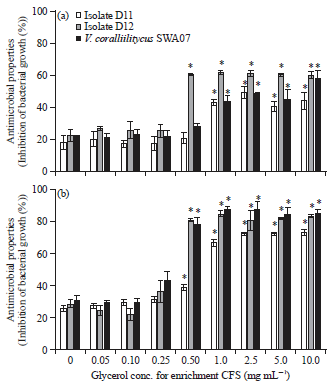 | |
| Fig. 3(a-b): | Growth inhibition (%, Mean±SD, n = 3) of three target bacteria (D11, D12 and V. coralliilitycus SWA07) by CFSs harvested at 48 h cultured (a) MH66 and (b) MB3 Tukey HSD test (*p<0.01) |
| Table 4: | The percentage growth of isolates MH66 and MB3 grown in various glycerol concentrations |
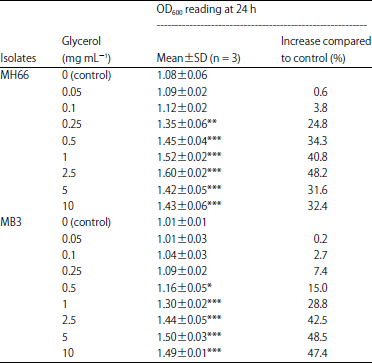 | |
| A positive significant effect of glycerol concentrations was obtained by Tukey HSD test (*p<0.01, **p<0.001 and ***p<0.0001) when compared to control (0 mg mL–1) | |
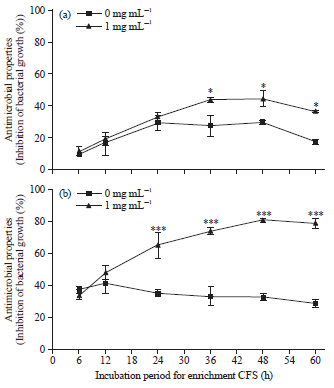 | |
| Fig. 4 (a-b): | Growth inhibition (%, Mean±SD, n = 3) of V. coralliilitycus SWA07 grown by CFSs harvested at various incubation periods (6, 12, 24, 36, 48 and 60 h) of (a) MH66 and (b) MB3 |
| (*p<0.01, **p<0.001 and ***p<0.0001) | |
Results presented in this study revealed that glycerol concentrations from 0.5 to 10 mg mL–1 significantly enhanced the antimicrobial properties of MH66 and MB3 in comparison to without glycerol (0 mg mL–1). Glycerol concentration of 1 mg mL–1 showed to be one of the most significant enhancing effects on antimicrobial properties of MH66 and MB3.
Effect of glycerol on the antimicrobial properties in different growth phases: The percentage inhibition against the growth of V. coralliilyticus SWA07 varied significantly with respect to antimicrobial properties of CFSs from MH66 and MB3 at 1 mg mL–1 glycerol concentration and incubation period for enrichment. As shown in Fig. 4a, the growth inhibition (%) of MH66 CFS enriched with 1 mg mL–1 was significantly higher than without glycerol at 36, 48 and 60 h incubation periods. For the case of MB3, CFS enriched with 1 mg mL–1 was significantly greater than without glycerol at 24, 36, 48 and 60 h incubation periods (Fig. 4b). From the above results, the fourth hypothesis was supported indicating that glycerol enhanced the antimicrobial properties of MH66 and MB3.
DISCUSSION
This study findings added more knowledge to the roles of glycerol played in the coral holobiont by increasing the concentration of glycerol when the corals were exposed to light and temperature stresses and it could also help in structuring certain Vibrio bacteria by enhancing their growth, antibacterial and antagonistic effects.An increase in glycerol concentration when healthy coral M. digitata was exposed to extended lighting period (18 h) or combined with thermal stress (32°C) was noted. This finding were related to the results of Suescun-Bolivar et al.11,13, who proposed an increase in the synthesis of glycerol as redox balance for zooxanthellae and to prevent feedback inhibition of photosynthesis13. Results in this study showed that thermal stress alone did not increase glycerol concentration in the tissue of M. digitata. Similar results were obtained with the coral Porites cylindrica40. This could be so because high temperature alone did not enhance the glycerol synthesis. However, the effect of high temperature stress might enhance the release of glycerol from the zooxanthellae to the coral tissue (translocation). A similar observation was noted in two species of algae, Dunaliella and Asteromonas, which increased the release of glycerol when exposed to high temperature41. These patterns could explain the effect of combined light-thermal stresses, in which highest concentration of glycerol was noticed.
In the present study, the antimicrobial properties of culturable bacteria isolated from coral M. digitata showed to be complex. Six (8.5%) of the screened coral bacterial isolates showed antimicrobial properties against V. coralliilyticus SWA07 which is known as a coral pathogen. Similarly, Shiroyama et al.25 reported that 13% of screened isolates from M. digitata displayed growth inhibition against V. coralliilyticus. These findings agreed with previous reports stated that bacteria isolated from corals inhibited putative coral pathogens in vitro and various coral-associated bacteria14,18,25,42. Out of the six coral bacterial isolates, MH66 and MB3 displayed antimicrobial and/or antagonistic activities against most of the target bacteria. MH66 was closely related to V. alginolyticus, which displayed antimicrobial properties by producing the bioactive compound Tetrodotoxin43. MB3 was closely related to V. coralliilyticus, which is also known for its antimicrobial properties through the production of the antibiotic compound Andrimid44. Therefore, these Vibrio-associated isolates MH66 and MB3 in this study might protect the coral by producing antimicrobial agents from secondary metabolism that could compete with other coral bacteria to keep the coral healthy. However, previous studies had also found that Vibrio-associated isolates, for example V. alginolyticus 22 and V. coralliilyticus 21, could be potential coral pathogens, although they displayed antimicrobial/antagonistic activity16,17,25. Since both Vibrio isolates, MH66 and MB3, possessed antagonistic and antimicrobial properties, they could play important roles in influencing and shaping the bacterial community, abundance and diversity in the coral holobiont15,17. Glycerol significantly enhanced the antimicrobial activity of Vibrio MH66 and MB3 against the growth of target bacteria but was dependent on the glycerol concentrations used for enrichment of CFSs, especially at the stationary phase of MH66 and MB3. This finding agreed with previous studies which showed that glycerol enhanced antibiotic synthesis of bacteria through secondary metabolism45,46. For instance, glycerol enhanced the antibiotic compound, Andrimid, by Serratia plymuthica 47. Another study reported that V. coralliilyticus S2052 produced antibiotic Andrimid in the late exponential phase44,48. Several mechanisms proposed by Gorke and Stulke49 and Sanchez et al.30tried to explain for glycerol-regulated antibiotic synthesis from bacterial secondary metabolism.
Results from this study indicated that the presence of glycerol in bacterial culture enhanced the growth of Vibrio MH66 and MB3, which were in line with that of Ramesh et al.50 who showed that glycerol assisted in the optimum growth of the marine bacteria V. campbelli. Likewise, some previous studies have demonstrated that glycerol promoted the growth of Gluconobacter oxydans and Clostridium butyricum at higher concentrations (>10 mg mL–1of glycerol)37,51. Previous related studies demonstrated that glycerol was a second carbon source which was consumed for the growth of bacteria through different metabolic pathways26,28,29,52. These might explain why in this study, the presence of glycerol in the culture medium enhanced the growth of the isolates MH66 and MB3.
CONCLUSION
An extended light exposure and combined with high temperature would increase the glycerol concentration in healthy coral M. digitata. Two Vibrio-associated bacteria isolated from M. digitata (MH66 and MB3) displayed antimicrobial activity against coral potential pathogens and antagonistic activity against other coral bacterial isolates. Glycerol enhanced the growth and antimicrobial properties of MH66 and MB3.
SIGNIFICANCE STATEMENT
Findings of this study will help to better understand the functions and roles of glycerol in the coral holobiont. Moreover, glycerol could influence the population of certain Vibrio associated-coral bacteria. Besides, this study will help further research on antibiotic compounds from bacteria, especially Vibrio spp., isolated from coral/s. Finally, this study highlighted the importance of glycerol and its indirect benefits between zooxanthellae and coral-associated bacteria.
ACKNOWLEDGMENT
S
The authors are grateful to the staff at Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus, Japan, for providing the necessary facilities during this study. We would like to especially thank Mr. Uehara S. for his help with the fieldwork. The authors are also thankful to all the members of the Marine Ecosystem and Biogeochemistry Laboratory, Shizuoka University, Japan.
REFERENCES
- Rosenberg, E., O. Koren, L. Reshef, R. Efrony and I. Zilber-Rosenberg, 2007. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Microbiol., 5: 355-362.
Direct Link - Thompson, J.R., H.E. Rivera, C.J. Closek and M. Medina, 2015. Microbes in the coral holobiont: Partners through evolution, development and ecological interactions. Front. Cell. Infect. Microbiol., Vol. 4.
CrossRefDirect Link - Sangsawang, L., B.E. Casareto, H. Ohba, H.M. Vu and A. Meekaew et al., 2017. 13C and 15N assimilation and organic matter translocation by the endolithic community in the massive coral Porites lutea. R. Soc. Open Sci., Vol. 4.
Direct Link - Davy, S.K., D. Allemand and V.M. Weis, 2012. Cell biology of cnidarian-dinoflagellate symbiosis. Microbiol. Mol. Biol. Rev., 76: 229-261.
CrossRefDirect Link - Gaither, M.R. and R. Rowan, 2010. Zooxanthellar symbiosis in planula larvae of the coral Pocillopora damicornis. J. Exp. Mar. Biol. Ecol., 386: 45-53.
CrossRefDirect Link - Tremblay, P., R. Grover, J.F. Maguer, L. Legendre and C. Ferrier-Pages, 2012. Autotrophic carbon budget in coral tissue: A new 13C-based model of photosynthate translocation. J. Exp. Biol., 215: 1384-1393.
CrossRefDirect Link - Muscatine, L., 1967. Glycerol excretion by symbiotic algae from corals and Tridacna and its control by the host. Science, 156: 516-519.
CrossRefDirect Link - Trench, R.K., 1971. The physiology and biochemistry of zooxanthellae symbiotic with marine coelenterates I. The assimilation of photosynthetic products of zooxanthellae by two marine coelenterates. Proc. R. Soc. Lond. B, 177: 225-235.
CrossRefDirect Link - Burriesci, M.S., T.K. Raab and J.R. Pringle, 2012. Evidence that glucose is the major transferred metabolite in dinoflagellate-cnidarian symbiosis. J. Exp. Biol., 215: 3467-3477.
CrossRefDirect Link - Molina, V.H., R.E. Castillo-Medina and P.E. Thome, 2017. Experimentally induced bleaching in the sea anemone exaiptasia supports glucose as a main metabolite associated with its symbiosis. J. Mar. Biol., Vol. 2017.
CrossRefDirect Link - Suescun-Bolivar, L.P., R. Iglesias-Prieto and P.E. Thome, 2012. Induction of glycerol synthesis and release in cultured symbiodinium. PLoS One, Vol. 7.
CrossRefDirect Link - Mayfield, A.B. and R.D. Gates, 2007. Osmoregulation in anthozoan-dinoflagellate symbiosis. Comp. Biochem. Physiol.-A Mol. Integr. Physiol., 147: 1-10.
CrossRefDirect Link - Suescun-Bolivar, L.P., G.M.I. Traverse and P.E. Thome, 2016. Glycerol outflow in Symbiodinium under osmotic and nitrogen stress. Mar. Biol., Vol. 163.
CrossRefDirect Link - Peixoto, R.S., P.M. Rosado, D.C. de Assis Leite, A.S. Rosado and D.G. Bourne, 2017. Beneficial microorganisms for corals (BMC): Proposed mechanisms for coral health and resilience. Front. Microbiol., Vol. 8.
CrossRefDirect Link - Kvennefors, E.C.E., E. Sampayo, C. Kerr, G. Vieira, G. Roff and A.C. Barnes, 2012. Regulation of bacterial communities through antimicrobial activity by the coral holobiont. Microb. Ecol., 63: 605-618.
CrossRefDirect Link - Ritchie, K.B., 2006. Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar. Ecol. Prog. Ser., 322: 1-14.
CrossRefDirect Link - Rypien, K.L., J.R. Ward and F. Azam, 2010. Antagonistic interactions among coral-associated bacteria. Environ. Microbiol., 12: 28-39.
CrossRefDirect Link - Nissimov, J., E. Rosenberg and C.B. Munn, 2009. Antimicrobial properties of resident coral mucus bacteria of Oculina patagonica. FEMS Microbiol. Lett., 292: 210-215.
CrossRefDirect Link - Gajigan, A.P., L.A. Diaz and C. Conaco, 2017. Resilience of the prokaryotic microbial community of Acropora digitifera to elevated temperature. MicrobiologyOpen, Vol. 2017.
CrossRefDirect Link - Tout, J., N. Siboni, L.F. Messer, M. Garren and R. Stocker et al., 2015. Increased seawater temperature increases the abundance and alters the structure of natural Vibrio populations associated with the coral Pocillopora damicornis. Front. Microbiol., Vol. 6.
CrossRefDirect Link - Ben-Haim, Y., M. Zicherman-Keren and E. Rosenberg, 2003. Temperature-regulated bleaching and lysis of the coral Pocillopora damicornis by the novel pathogen Vibrio coralliilyticus. Applied Environ. Microbiol., 69: 4236-4242.
CrossRefDirect Link - Zhenyu, X., K. Shaowen, H. Chaoqun, Z. Zhixiong, W. Shifeng and Z. Yongcan, 2013. First characterization of bacterial pathogen, Vibrio alginolyticus, for Porites andrewsi white syndrome in the South China sea. Plos One, Vol. 8.
CrossRefDirect Link - Munn, C.B., 2015. The role of vibrios in diseases of corals. Microbiol. Spectrum, Vol. 3, No. 4.
CrossRefDirect Link - Holmstrom, C. and S. Kjelleberg, 1999. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS. Microbiol. Ecol., 30: 285-293.
CrossRefPubMedDirect Link - Murarka, A., Y. Dharmadi, S.S. Yazdani and R. Gonzalez, 2008. Fermentative utilization of glycerol by Escherichia coli and its implications for the production of fuels and chemicals. Applied Environ. Microbiol., 74: 1124-1135.
CrossRefDirect Link - Bizzini, A., C. Zhao, A. Budin-Verneuil, N. Sauvageot, J.C. Giard, Y. Auffray and A. Hartke, 2010. Glycerol is metabolized in a complex and strain-dependent manner in Enterococcus faecalis. J. Bacteriol., 192: 779-785.
CrossRefDirect Link - Kopp, J., C. Slouka, S. Ulonska, J. Kager, J. Fricke, O. Spadiut and C. Herwig, 2017. Impact of glycerol as carbon source onto specific sugar and inducer uptake rates and inclusion body productivity in E. coli BL21 (DE3). Bioengineering, Vol. 5, No. 1.
CrossRefDirect Link - Claret, C., J. Salmon, C. Romieu and A. Bories, 1994. Physiology of Gluconobacter oxydans during dihydroxyacetone production from glycerol. Applied Microbiol. Biotechnol., 41: 359-365.
CrossRefDirect Link - Sanchez, S., A. Chavez, A. Forero, Y. Garcia-Huante and A. Romero et al., 2010. Carbon source regulation of antibiotic production. J. Antibiot., 63: 442-459.
CrossRefPubMedDirect Link - Martin, J.F. and A.L. Demain, 1980. Control of antibiotic biosynthesis. Microbiol. Rev., 44: 230-251.
PubMedDirect Link - Suzuki, T., B.E. Casareto, Y. Shioi, Y. Ishikawa and Y. Suzuki, 2015. Finding of 132, 173-cyclopheophorbide a enol as a degradation product of chlorophyll in shrunk zooxanthellae of the coral Montipora digitata. J. Phycol., 51: 37-45.
CrossRefDirect Link - Ramphul, C., B.E. Casareto, H. Dohra, T. Suzuki, K. Yoshimatsu, K. Yoshinaga and Y. Suzuki, 2017. Genome analysis of three novel lytic Vibrio coralliilyticus phages isolated from seawater, Okinawa, Japan. Mar. Genomics, 35: 69-75.
CrossRefDirect Link - Sousa, A.M., I. Machado, A. Nicolau and M.O. Pereira, 2013. Improvements on colony morphology identification towards bacterial profiling. J. Microbiol. Methods, 95: 327-335.
CrossRefDirect Link - Dilhari, A., A. Sampath, C. Gunasekara, N. Fernando and D. Weerasekara et al., 2017. Evaluation of the impact of six different DNA extraction methods for the representation of the microbial community associated with human chronic wound infections using a gel-based DNA profiling method. AMB Express, Vol. 7, No. 1.
CrossRefDirect Link - Weisburg, W.G., S.M. Barns, D.A. Pelletier and D.J. Lane, 1991. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol., 173: 697-703.
CrossRefPubMedDirect Link - Stasiak-Rozanska, L., S. Blazejak and I. Gientka, 2014. Effect of glycerol and dihydroxyacetone concentrations in the culture medium on the growth of acetic acid bacteria Gluconobacter oxydans ATCC 621. Eur. Food Res. Technol., 239: 453-461.
CrossRefDirect Link - Shnit-Orland, M., A. Sivan and A. Kushmaro, 2012. Antibacterial activity of Pseudoalteromonas in the coral holobiont. Microb. Ecol., 64: 851-859.
CrossRefDirect Link - Couch, C.S., L.D. Mydlarz, C.D. Harvell and N.L. Douglas, 2008. Variation in measures of immunocompetence of sea fan coral, Gorgonia ventalina, in the Florida keys. Mar. Biol., 155: 281-292.
CrossRefDirect Link - Fitt, W.K., R.D. Gates, O. Hoegh-Guldberg, J.C. Bythell and A. Jatkar et al., 2009. Response of two species of Indo-Pacific corals, Porites cylindrical and Stylophora pistillata, to short-term thermal stress: The host does matter in determining the tolerance of corals to bleaching. J. Exp. Mar. Biol. Ecol., 373: 102-110.
CrossRefDirect Link - Wegmann, K., A. Ben-Amotz and M. Avron, 1980. Effect of temperature on glycerol retention in the halotolerant algae Dunaliella and Asteromonas. Plant Physiol., 66: 1196-1197.
CrossRefDirect Link - Krediet, C.J., K.B. Ritchie, V.J. Paul and M. Teplitski, 2013. Coral-associated micro-organisms and their roles in promoting coral health and thwarting diseases. Proc. R. Soc. Lond. B: Biol. Sci., Vol. 280.
CrossRefDirect Link - Noguchi, T., D.F. Hwang, O. Arakawa, H. Sugita, Y. Deguchi, Y. Shida and K. Hashimoto, 1987. Vibrio alginolyticus, a tetrodotoxin-producing bacterium, in the intestines of the fish Fugu vermicularis vermicularis. Mar. Biol., 94: 625-630.
CrossRefDirect Link - Wietz, M., M. Mansson and L. Gram, 2011. Chitin stimulates production of the antibiotic andrimid in a Vibrio coralliilyticus strain. Environ. Microbiol. Rep., 3: 559-564.
CrossRefDirect Link - Bhattacharyya, B.K., S.C. Pal and S.K. Sen, 1998. Antibiotic production by Streptomyces hygroscopicus D1. 5: Cultural effect. Rev. Microbiol., 29: 167-169.
CrossRefDirect Link - El-Banna, N.M., 2006. Effect of carbon source on the antimicrobial activity of Corynebacterium kutscheri and Corynebacterium xerosis. Afr. J. Biotechnol., 5: 833-835.
Direct Link - Matilla, M.A., V. Nogellova, B. Morel, T. Krell and G.P. Salmond, 2016. Biosynthesis of the acetyl-CoA carboxylase-inhibiting antibiotic, andrimid in Serratia is regulated by Hfq and the LysR-type transcriptional regulator, AdmX. Environ. Microbiol., 18: 3635-3650.
CrossRefDirect Link - Giubergia, S., C. Phippen, K.F. Nielsen and L. Gram, 2017. Growth on chitin impacts the transcriptome and metabolite profiles of antibiotic-producing Vibrio coralliilyticus S2052 and Photobacterium galatheae S2753. Msystems, Vol. 2, No. 1.
CrossRefDirect Link - Gorke, B. and J. Stulke, 2008. Carbon catabolite repression in bacteria: Many ways to make the most out of nutrients. Nat. Rev. Microbiol., 6: 613-624.
Direct Link - Ramesh, C., R. Mohanraju, K.N. Murthy, P. Karthick and S. Narayana, 2014. Impact of light, temperature, salinity and glycerol on the intensity of luminescence and growth of marine bioluminescent bacteria Vibrio campbelli (strain STF1). Curr. Sci., 106: 511-513.
Direct Link - Szymanowska-Powalowska, D., 2015. The effect of high concentrations of glycerol on the growth, metabolism and adaptation capacity of Clostridium butyricum DSP1. Electron. J. Biotechnol., 18: 128-133.
CrossRefDirect Link - Ameyama, M., E. Shinagawa, K. Matsushita and O. Adachi, 1985. Solubilization, purification and properties of membrane-bound glycerol dehydrogenase from Gluconobacter industrius. Agric. Biol. Chem., 49: 1001-1010.
CrossRefDirect Link