
Rebeca Inhoque Pereira
Department of Microbiology, Immunology and Parasitology, Federal University of Rio Grande do Sul (UFRGS), Sarmento Leite, 500, Room 158, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
Janira Prichula
Department of Microbiology, Immunology and Parasitology, Federal University of Rio Grande do Sul (UFRGS), Sarmento Leite, 500, Room 158, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
Naiara. Aguiar Santestevan
Department of Microbiology, Immunology and Parasitology, Federal University of Rio Grande do Sul (UFRGS), Sarmento Leite, 500, Room 158, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
Pedro Alves d`Azevedo
Gram-positive Coccus Laboratory, Federal University of Health Sciences of Porto Alegre (UFCSPA), Sarmento Leite, 245, Room 204, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
Amanda de Souza Motta
Department of Microbiology, Immunology and Parasitology, Federal University of Rio Grande do Sul (UFRGS), Sarmento Leite, 500, Room 158, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
Ana Paula Guedes Frazzon
Department of Microbiology, Immunology and Parasitology, Federal University of Rio Grande do Sul (UFRGS), Sarmento Leite, 500, Room 158, ZIP Code 90050-170, Porto Alegre, Rio Grande do Sul, Brazil
LiveDNA: 55.16346
Research Journal of Microbiology
Year: 2017 | Volume: 12 | Issue: 4 | Page No.: 248-254







ABSTRACT
Background and Objective: The buffalo milk consumption and derivatives have increased significantly in the last year due to the healthy food demand. Enterococci play a beneficial role during the maturation of some cheese and sausages; they have been used as probiotics in humans and animals. On the other hand, they are indicators of fecal contamination and are frequently associated with foodborne illnesses by biogenic amines. The aim of this study was to evaluate the presence of virulence profiles in enterococci strains isolated from raw buffalo’s milk samples. Materials and Methods: Seventy-nine enterococci species were selected which previously identified by conventional biochemical methods. The strains were submitted to genotypic identification using genus-specific and species-specific primers. Strains were tested for the presence of virulence genes (agg, ace, gelE) by PCR, their ability to form biofilms and to produce the enzyme gelatinase by phenotypic methods. The optical density (OD) of bacterial biofilms was quantified in a spectrophotometer. Results: The phenotypic and genotypic identification were similar in more than 96% of the strains. The frequency of ace (96 vs. 10.34%) and gelE (96 vs. 17.24%) genes were higher in E. faecalis than in E. faecium, while the agg gene was detected only in E. faecalis strains (26%). The in vitro biofilm ability was observed in both strains; however, it was superior among E. faecalis (90%) than in E. faecium (24.1%). The presence of gelE and the activity of gelatinase were not fully concordant. Conclusion: It was concluded that the presences of enterococci harboring virulent factors in raw buffalo’s milk suggest a situation of risk for the community, since enterococci are opportunist pathogens. The ability to form biofilm is important for food safety and the protection of public health. In this sense, the present study sought to collaborate with the status quo of scientific knowledge to improve safety and quality of the food for human consumption.
PDF Abstract XML References Citation
Received: April 27, 2017;
Accepted: August 23, 2017;
Published: September 15, 2017
How to cite this article
Rebeca Inhoque Pereira, Janira Prichula, Naiara. Aguiar Santestevan, Pedro Alves d`Azevedo, Amanda de Souza Motta and Ana Paula Guedes Frazzon, 2017. Virulence Profiles in Enterococcus spp. Isolated from Raw Buffalo’s Milk in South Brazil. Research Journal of Microbiology, 12: 248-254.
URL: https://scialert.net/abstract/?doi=jm.2017.248.254
URL: https://scialert.net/abstract/?doi=jm.2017.248.254
INTRODUCTION
The buffalo milk has a high concentration of proteins and fats, low cholesterol content, than other animals1. Buffalo milk is the raw material used to prepare buffalo mozzarella cheese and other dairy products. The mozzarella cheese is the main product, being the largest part of the production of milk intended for their manufacture1,2. In Brazil, the mozzarella cheese is produced from the pasteurized buffalo’s milk, however some production system follows Italian process and work with raw milk3,4. This process has the purpose to ensure the particular organoleptic characteristics and inherent to the product, despite the importance of pasteurization3.
Enterococcus spp. are lactic acid bacteria, producing bacteriocins and are widely distributed in nature, present in soils, waters, foods, plants and vegetables5-7. Moreover, this genera is commonly found in the gastrointestinal tract of humans and animals8-11. In addition, they are tolerant to high concentrations of salts, pH variations and a wide temperature range. This rusticity has been demonstrated that some enterococcal strains can survive at temperatures of milk pasteurization7.
The genus Enterococcus contains over 50 recognized species and Enterococcus faecalis (E. faecalis) and Enterococcus faecium (E. faecium) are the most frequent species isolated from clinical and food7. Among the species, E. faecalis is recognized to have a well elucidated arsenal of virulence factors, when compared to the others, possessing multiple virulence determinants, which reinforces the evidence of pathogenicity12,13. The presence of enterococci in foods can be an indicator of fecal contamination in the production and/or processing of food and also serve as reservoirs of genes for resistance, which can spread through the food chain, thus contributing to the spread of antibiotic resistance in the human population. Epidemiological studies show that by their nature opportunist, enterococci have emerged as pathogens associated with nosocomial infections, which may be related, in part, to antibiotic resistance and the presence of virulence determinants7,14,15.
Virulence determinants are factors which enable the bacteria to colonize, invade, prevent the defense system and cause tissue damage to the host16. Among the virulence factors widely found in this genus, highlights the gelatinase enzyme (encoded by the gelE); aggregation substance of Enterococcus (encoded by agg gene), adhesion of collagen from E. faecalis (encoded by ace) and the ability to form biofilm17.
Studies that assess the microbiological quality of buffalo’s milk are still scarce6, due to the difficulty in obtaining samples. In this scenario, the knowledge of strains circulating in food, contributes to a better understanding of the opportunistic behavior of these microorganisms in the food chain. Given this, the aim of this study was to evaluate the presence of virulence genes, the ability to form in vitro biofilm and to produce the gelatinase in enterococci isolates from raw buffalo’s milk samples in South Brazil.
MATERIALS AND METHODS
Enterococcus strains: The study occurred in March, 2014 with 79 enterococci (51 E. faecalis; 23 E. faecium; two E. duran and three Enterococcus spp.), isolated from raw buffalo’s milk samples in South Brazil6, during June-August, 2012, were selected (Table 1). All the strains have already been subjected to conventional biochemical tests by Prichula et al.6. The strains were maintained in a solution of 10% (w/v) of skim milk (Difco) and 10% (v/v) glycerol (Neon Comercial Ltda.), frozen at -20°C, in the Coccus Gram-positive Laboratory and Molecular Microbiology at Federal University of Health Sciences of Porto Alegre (UFCSPA).
It is important to note that the 79 bacteria used in this study, 13.9% (11/79) showed resistance to nitrofurantoin, 12.7% (10/79) to tetracycline, 1.3% (1/79) to chloramphenicol, 1.3% (1/79) to streptomycin, 1.3% (1/79) to norfloxacin and 1.3% (1/79) to erythromycin. Forty-seven isolates (59.5%) showed intermediate resistance to norfloxacin, 58.2% (46/79) to ciprofloxacin, 51.9% (41/79) to erythromycin and 6.3% (5/79) to the nitrofurantoin6.
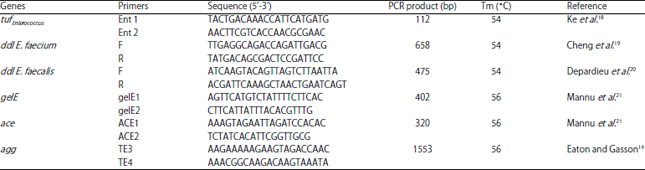
Molecular identification of genus by PCR: All strains were confirmed as genus by polymerase chain reaction (PCR) using a genus-specific primer of tufEnterococcus gene, according to the protocols described by Ke et al.18 (Table 2).
| Table 1: | Genotypic and phenotypic profile of enterococci isolated from buffalo’s milk evaluated in this study |
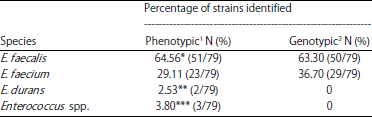 | |
1Phenotypic identification by Prichula et al.4. 2Identification based on species-specify PCR. *One strain was reclassified as E. faecium after PCR. **Species reclassified as E. faecium after PCR. ***Species identified as Enterococcus spp. and identified an E. faecium after PCR | |
| Table 2: | Primers and annealing temperatures used in the PCRs carried out in this study |
 | |
Genomic DNA was extracted by chemical analysis method following the method described by Donato22. The PCR was carried18 out in a reaction volume of 25 μL, containing 100 ng of genomic DNA, 1.5 mM MgCl2, 10 μM of each primer tuf gene, 200 μM of dNTPs, 1U Taq DNA polymerase and reaction buffer 1X. Amplification was carried out in an Eppendorf Mastercycler Personal 5332 Thermocycler (Eppendorf®) according to the following program: 95°C for 3 min followed by 35 cycles of 95°C for 30 sec, 54°C for 30 sec, 72°C for 1 min, followed by final extension at 72°C for 7 min. The PCR products were analyzed by electrophoresis gel with 1.5% agarose, stained with ethidium bromide solution.
Species confirmation by polymerase chain reaction: A specie-specific PCR assay followed was used to confirm the phenotypic identification. Listed in Table 2 are the primers. The conditions of the PCR were previously described18. The DNA fragment amplified by PCR was analyzed in 1.5 % (w/v) agarose gels.
Virulence profile
Detection of virulence genes by PCR: The presence of the agg (aggregation substance of Enterococcus), ace (adhesion of collagen from E. faecalis) and gelE (expression of gelatinase enzyme) genes followed as described by Medeiros et al.23. The PCR for ace, agg and gelE genes was performed in 25 μL total volume and conditions have been described previously14,18,21. The DNA fragments amplified by PCR were analyzed in 1.5% (w/v) agarose gels.
Gelatinase activity: The gelatinase activity was performed in tubes, following the protocol described by Marra et al.24.
In vitro biofilm formation on polystyrene microplate: All strains were tested for the ability to form biofilm in vitro by using the crystal violet (CV) protocol 25. Briefly, the experiment was divided into three stages: First, the bacteria were previously inoculated on agar plates containing Brain Heart Infusion (BHI-Himedia) and incubated to 35-37°C for 18 h. After the growth, the colonies were resuspended in saline and adjusted scale of 0.5 McFarland standards (approximately 108 CFU mL–1). The wells of sterile 96-well flat-bottomed polystyrene microplates were filled with 180 μL of Tryptic Soy Broth (TSB-Himedia) supplemented with 1% glucose and 20 μL of bacterial suspension (containing approximately 108 CFU mL–1). Afterwards, the plates were incubated for 18 h at 36°C. The optical density (OD) of bacterial biofilms was quantified in a using 492 nm wavelength in a spectrophotometer (Anthos 2010 Microplate Reader). The OD of each strain was determined by the arithmetic mean of the absorbance of the wells and this value was compared with the mean absorbance of negative controls (ODnc). The strains were separated into categories using the O.D values, as: non-biofilm (ODs<ODnc), weak biofilm (ODnc<ODs<2.ODnc), moderate biofilm (2.ODnc<ODs<4.ODnc) and strong biofilm producers (4.ODnc<ODs). All biofilms assays were run in 8 replicates and repeated three times. Staphylococcus epidermidis American type culture collection 35984 classified as a strong adherent was used as the positive control and TSB plus 1% glucose as a negative control25.
RESULTS
Genotypic identification of Enterococcus spp. isolates from raw buffalo’s milk: The results between phenotypic and genotypic identification were similar in more than 96% of the strains. Only three strains (two E. durans and one E. faecalis) had discordant between phenotypic and genotypic identification (Table 1). However, for the genotypic identification those strains were reclassified as atypical E. faecium. These isolates had phenotypic traits similar to E. durans and E. faecalis, indicating the need to further evaluate their taxonomic position. The three strains are phenotypic characterized as Enterococcus spp. were genotypically identified as E. faecium.
| Table 3: | Distribution of virulence genes among E. faecalis and E. faecium isolated from buffalo’s milk in South Brazil |
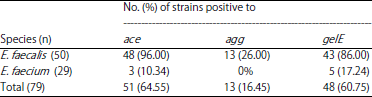 | |
Analysis of virulence profiles
Frequency of the virulence genes by PCR and detection of gelatinase activity: All isolates were evaluated for the presence of virulence genes by PCR and the results are presented in Table 3. The virulence genes were more frequent in E. faecalis when compared to E. faecium. In the present study, the ace gene was detected in 96% (48/50) of E. faecalis and 10.34% (3/29) of E. faecium. The agg gene was detected only in E. faecalis strains (26%).
The gelE gene was detected in 86% (43/50) and 17.24% (5/29) and E. faecium, respectively. The gene gelE encodes a gelatinase and of the 79 strains evaluated, 28 showed gelatinase activity, in addition this phenotype was observed only in E. faecalis. For some strains, the gelatinase activity in vitro was not correlate with the presence of gelE genes. Analysis of congruence between gelatinase and gelE pointed out that 30% (15/50) E. faecalis and 17.24% (5/29) E. faecium positive to gelE gene were unable to degraded gelatin as a substrate.
In vitro biofilm formation ability of enterococci isolates from raw buffalo’s milk: Biofilm formation of E. faecalis and E. faecium isolates from raw buffalo’s milk samples were assayed and categorized based on the OD492. The biofilm ability was observed in high frequency among E. faecalis (90%) than in E. faecium (24.1%). Of the 50 E. faecalis strains, 10 (20%), 32 (64%) 3 (6%) and 5 (10%) were classified as strong, moderate, weak and non-biofilm formers, respectively. Among the 29 E. faecium, only 4 (13.8%) were strong biofilm former and two (10.3%) moderates and 22 (75.9%) non-biofilm former.
Since the expression of gelatinase may be influencing the formation of biofilm, it was observed that more than half of E. faecalis strains gelatinase positive (56%) were the strains classified as moderate (64%) and strong (20%) biofilm former.
DISCUSSION
Identification of Enterococcus spp. by PCR: Of the 79 enterococci tested, three strains had discordant between phenotypic and genotypic identification and were reclassified as atypical E. faecium. Atypical enterococci have already been identified in strains isolated from food origin5. In addition, Jahan et al.26 reported that negative results of biochemical tests can be explained the metabolic diversity of organisms in this genus. Nevertheless, genotypic assays are quicker, practical and more sensitive when compared with the biochemical tests. Knudtson and Hartman27 and Moore et al.28, showed that identification based on molecular tools are more accurate detection, avoid long periods of incubation of culture and eliminate phenotypic results ambiguous.
Analysis of virulence factors
Virulence genes and detection of gelatinase activity: The acquisition of virulence factors enables the bacteria to survive and reproduce in competitive environments. Differences in the frequency of virulence genes were observed among E. faecalis and E. faecium isolates from raw buffalo’s milk samples. Enterococcus faecalis are known to harbored more virulence genes than E. faecium. Cariolato et al.13, Gomes et al.29 and Barbosa et al.30, have demonstrated a high frequency of multiple virulence factors in E. faecalis strains, however the same pattern was not observed in E. faecium strains. Mannu et al.21 pointed out that E. faecium is a species generally free of known virulence factors, although there are some references related to occasional virulence determinants in this isolated species14,31,32.
Virulence genes detected in this study were ace, agg and gel. A high frequency of ace gene in E. faecalis (96%) was reported in this study. Gomes et al.29, in agreement with this study, detected this gene in food and clinical isolates, belonging to species other than E. faecalis. However, Cariolato et al.13 described the gene ace only in E. faecalis strains isolated dairy products. The ace gene encodes a collagen adhesion protein involved in mediating adherence to epithelial. The ace gene has been associated with infections caused by E. faecalis, antibodies against ace have been almost always found in serum from patients with endocarditis15.
The agg gene which encodes an aggregation substance that facilitates cell adhesion was detected only in E. faecalis isolates. This result is in agreement with several researchers that described agg gene only in E. faecalis strains14,28. Eaton and Gasson14 when analyzing Enterococcus foodborne isolates, isolated clinical and used as starter cultures, established the agg gene only in strains of E. faecalis.
In the present study, gelE gene was detected in E. faecalis and E. faecium. Moraes et al.33 reported a high frequency of gelE in Enterococcus strains isolated from raw milk and cheese in Minas Gerais state, Brazil. The gelatinase is an enzyme capable of hydrolyzing substrates such as gelatin, collagen, casein, hemoglobin and other small biologically active peptides14,34. The expression of gelatinase in Enterococcus isolates from raw buffalo’s milk samples can be explained by the fact that these samples are rich in casein and the production of gelatinase may be a selection mechanism for growth allowing the microorganisms using the casein as a source of amino acids. The presence of gelE genes did not always correlate with gelatinase. According to Marra et al.24, the presence of the gene gelE is not necessarily correlated with the phenotype of gelatinase. This result may be justified by four factors: (a) The loss of gelatinase activity during laboratory manipulation11, (b) Presence of silent genes, (c) Low levels of down regulation of gene expression or (d) Due to the influence of environmental factors in the expression of the enzyme14.
Biofilm formation in enterococci isolates from raw buffalo’s milk: In the food industries, biofilm has become a problem, mainly related to the fact that microorganisms in food processing equipment are resistant to sanitizers and sources of contamination. Ability to form biofilm was observed in E. faecalis and E. faecium isolated from buffalo’s milk samples; however, this ability was more frequently in E. faecalis strains. The results report in the literature about the formation of biofilm by enterococci in foods, corroborate with the results demonstrate here23,29. Microorganisms growing on food surfaces and in processing environments can cross-contaminate and cause post-processing contamination. The milk industries pay special attention to the cross-contamination during food process. Many outbreaks have been associated with biofilm. The clinical relevance of biofilms formed by enterococci is, mainly, their significantly higher antibiotic resistance relative to their free-living17.
CONCLUSION
In conclusion, the data indicate that enterococci isolated from buffalo’s milk samples may act as reservoirs of virulence genes, which can spread these genes to human’s microbiota through the food chain. The ability to form biofilm by the strains is important for food safety and the protection of public health, since enterococci within the biofilms are protected from sanitizer increasing the likelihood of survival and subsequent contamination of food. Additional investigation of these microorganisms in food, as well as, the characterization and identification of virulence profile is necessary to ensure the quality of the food. The increasing importance of Enterococcus as a nosocomial opportunistic and at the same time their utilization in the food industry as starter cultures and flavor aggregators, researches are necessary to ensure safety and quality of the food for human consumption.
SIGNIFICANCE STATEMENTS
This study evaluated virulence profiles and enterococci strains isolated from raw buffalo's milk samples. Since the buffalo milk consumption and derivatives have increased significantly in the last year, this study suggests an alert situation for the community, as enterococci are opportunist pathogens. The ability to form biofilm is important for food safety and the protection of public health. Additional investigation of these microorganisms in food, as well as, the characterization and identification of virulence profile is necessary to ensure the food security and the quality of the food.
ACKNOWLEDGMENTS
Authors thank the government agencies, National Council for Scientific and Technological Development (CNPq- #444335/2014-5, #300912/2012-9, #302421/2012-2 and #303251/2014-0) and the Coordination for the Improvement of Higher Education Personnel (CAPES) of the Brazilian government.
REFERENCES
- Abd El-Salam, M.H. and S. El-Shibiny, 2011. A comprehensive review on the composition and properties of buffalo milk. Dairy Sci. Technol., 91: 663-699.
CrossRefDirect Link - Buzi, K.A., J.P.A.N. Pinto, P.R.R. Ramos and G.F. Biondi, 2009. Microbiological analysis and electrophoretic characterization of mozzarella cheese made from buffalo milk. Food Sci. Technol., 29: 7-11.
CrossRefDirect Link - Hotta, R.M., C.A.F. Oliveira, O.C. Cunha Neto, A.M. Fernandes, P.J.A. Sobral and R.F. Neto, 2005. Effect of direct acidification and pasteurization of buffalo milk on the quality of marajoara cheese. Ital. J. Food Sci., 17: 295-303.
Direct Link - Pelicioli Riboldi, G., E.P. de Mattos, A.P. Guedes Frazzon, P.A. d'Azevedo and J. Frazzon, 2008. Phenotypic and genotypic heterogeneity of Enterococcus species isolated from food in Southern Brazil. J. Basic Microbiol., 48: 31-37.
Direct Link - Prichula, J., D.D.A. Zvoboda, R.I. Pereira, A.W. Medeiros and A.S. Motta et al., 2013. Antimicrobial susceptibility profile and diversity of enterococci species isolated from raw milk of buffalo in South Brazil. Rev. Bras. Cien. Vet., 20: 104-109.
CrossRefDirect Link - Lebreton, F., R.J.L. Willems and M.S. Gilmore, 2014. Enterococcus Diversity, Origins in Nature and Gut Colonization. In: Enterococci: From Commensals to Leading Causes of Drug Resistant Infection, Gilmore, M.S., D.B. Clewell, Y. Ike and N. Shankar (Eds.). Massachusetts Eye and Ear Infirmary, Boston, MA., USA.
- D'Azevedo, P.A., C.A.G. Dias and L.M. Teixeira, 2006. Genetic diversity and antimicrobial resistance of enterococcal isolates from Southern region of Brazil. Rev. Inst. Med. Trop. Sao Paulo, 48: 11-16.
CrossRefDirect Link - Cassenego, A.P.V., P.A. d'Azevedo, A.M.L. Ribeiro, J. Frazzon, S.T. Van Der Sand and A.P.G. Frazzon, 2011. Species distribution and antimicrobial susceptibility of enterococci isolated from broilers infected experimentally with Eimeria spp and fed with diets containing different supplements. Brazil. J. Microbiol., 42: 480-488.
Direct Link - Prichula, J., R.I. Pereira, G.R. Wachholz, L.A. Cardoso and N.C.C. Tolfo et al, 2016. Resistance to antimicrobial agents among enterococci isolated from fecal samples of wild marine species in the Southern coast of Brazil. Mar. Pollut. Bull., 105: 51-57.
CrossRefDirect Link - Medeiros, A.W., D.B. Amorim, M. Tavares, T.M. de Moura and A.C. Franco et al., 2017. Enterococcus species diversity in fecal samples of wild marine species as determined by real-time PCR. Can. J. Microbiol., 63: 129-136.
Direct Link - Abriouel, H., N.B. Omar, A.C. Molinos, R.L. Lopez and M.J. Grande et al., 2008. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol., 123: 38-49.
CrossRefPubMedDirect Link - Cariolato, D., C. Andrighetto and A. Lombardi, 2008. Occurrence of virulence factors and antibiotic resistances in Enterococcus faecalis and Enterococcus faecium collected from dairy and human samples in North Italy. Food Control, 19: 886-892.
CrossRefDirect Link - Eaton, T.J. and M.J. Gasson, 2001. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Applied Environ. Microbiol., 67: 1628-1635.
CrossRefDirect Link - Lin, Y.T., K.S. Hsieh, Y.S. Chen, I.F. Huang and M.F. Cheng, 2013. Infective endocarditis in children without underlying heart disease. J. Microbiol. Immunol. Infect., 46: 121-128.
CrossRefDirect Link - Moreno, M.R.F., P. Sarantinopoulos, E. Tsakalidou and L. de Vuyst, 2006. The role and application of enterococci in food and health. Int. J. Food Microbiol., 106: 1-24.
CrossRefDirect Link - Mohamed, J.A. and D.B. Huang, 2007. Biofilm formation by enterococci. J. Med. Microbiol., 56: 1581-1588.
CrossRefDirect Link - Ke, D., F.J. Picard, F. Martineau, C. Menard, P.H. Roy, M. Ouellette and M.G. Bergeron, 1999. Development of a PCR assay for rapid detection of enterococci. J. Clin. Microbiol., 37: 3497-3503.
Direct Link - Cheng, S., F.K. McCleskey, M.J. Gress, J.M. Petroziello and R. Liu et al., 1997. A PCR assay for identification of Enterococcus faecium. J. Clin. Microbiol., 35: 1248-1250.
PubMedDirect Link - Depardieu, F., B. Perichon and P. Courvalin, 2004. Detection of the van alphabet and identification of enterococci and staphylococci at the species level by multiplex PCR. J. Clin. Microbiol., 42: 5857-5860.
CrossRefDirect Link - Mannu, L., A. Paba, E. Daga, R. Comunian, S. Zanetti, I. Dupre and L.A. Sechi, 2003. Comparison of the incidence of virulence determinants and antibiotic resistance between Enterococcus faecium strains of dairy, animal and clinical origin. Int. J. Food Microbiol., 88: 291-304.
CrossRefDirect Link - Medeiros, A.W., R.I. Pereira, D.V. Oliveira, P.D. Martins and P.A. d'Azevedo et al., 2014. Molecular detection of virulence factors among food and clinical Enterococcus faecalis strains in South Brazil. Braz. J. Microbiol., 45: 327-332.
CrossRefDirect Link - Marra, A., F. Dib-Hajj, L. Lamb, F. Kaczmarek and W. Shang et al., 2007. Enterococcal virulence determinants may be involved in resistance to clinical therapy. Diagn. Microbiol. Infect. Dis., 58: 59-65.
CrossRefDirect Link - Stepanovic, S., D. Vukovic, I. Dakic, B. Savic and M. Svabic-Vlahovic, 2000. A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. J. Microbiol. Methods, 40: 175-179.
CrossRefDirect Link - Jahan, M., D.O. Krause and R.A. Holley, 2013. Antimicrobial resistance of Enterococcus species from meat and fermented meat products isolated by a PCR-based rapid screening method. Int. J. Food Microbiol., 163: 89-95.
CrossRefDirect Link - Knudtson, L.M. and P.A. Hartman, 1992. Routine procedures for isolation and identification of enterococci and fecal streptococci. Applied Environ. Microbiol., 58: 3027-3031.
Direct Link - Moore, D.F., M.H. Zhowandai, D.M. Ferguson, C. McGee, J.B. Mott and J.C. Stewart, 2006. Comparison of 16S rRNA sequencing with conventional and commercial phenotypic techniques for identification of enterococci from the marine environment. J. Applied Microbiol., 100: 1272-1281.
CrossRefDirect Link - Gomes, B.C., C.T. Esteves, I.C.V. Palazzo, A.L.C. Darini and G.E. Felis et al., 2008. Prevalence and characterization of Enterococcus spp. isolated from Brazilian foods. Food Microbiol., 25: 668-675.
CrossRefPubMedDirect Link - Barbosa, J., P.A. Gibbs and P. Teixeira, 2010. Virulence factors among enterococci isolated from traditional fermented meat products produced in the North of Portugal. Food Control, 21: 651-656.
CrossRefDirect Link - Soares-Santos, V., A.S. Barreto and T. Semedo-Lemsaddek, 2015. Characterization of enterococci from food and food-related settings. J. Food Prot., 78: 1320-1326.
CrossRefDirect Link - Abouelnaga, M., A. Lamas, M. Quintela-Baluja, M. Osman, J.M. Miranda, A. Cepeda and C.M. Franco, 2016. Evaluation of the extent of spreading of virulence factors and antibiotic resistance in enterococci isolated from fermented and unfermented foods. Ann. Microbiol., 66: 577-585.
CrossRefDirect Link - Moraes, P.M., L.M. Perin, S.D. Todorov, A. Silva Jr., B.D.G.M. Franco and L.A. Nero, 2012. Bacteriocinogenic and virulence potential of Enterococcus isolates obtained from raw milk and cheese. J. Applied Microbiol., 113: 318-328.
CrossRefDirect Link - Jett, B.D., M.M. Huycke and M.S. Gilmore, 1994. Virulence of enterococci. Clin. Microbiol. Rev., 7: 462-478.
Direct Link