
O.S. Folorunso
Department of Biochemistry,
K.O. Amisu
Department of Microbiology, Faculty of Science, Lagos State University, Ojo Lagos State, Nigeria
B.F. Ogungbe
Department of Biochemistry,
Research Journal of Microbiology
Year: 2015 | Volume: 10 | Issue: 4 | Page No.: 158-169







ABSTRACT
The ability of microbial cell membrane to bind organic dye is an age long protocol that preludes the identification and characterization of bacteria cells and its components. We employed formation of keratoconjunctivitis, pigmentations and Congo red binding assay to assess the virulence of eight enteric pathogenic bacteria. Staphylococcus aureus and Salmonella typhimurium, at 3.0×108 CFU mL-1, was sufficient to cause keratoconjunctivitis in the eyes of Albino rat (Sereny test), while observing traces in Streptococcus faecalis, Klebsiella pneumoniae, Pseudomonas aeruginosa and Bacillus subtilis. Pigmentation (PCr+) occurred in Staphylococcus aureus, Streptococcus faecalis and Klebsiella pneumoniae. Highest protein secretion was found in Escherichia coli. Hydrophobicity of the microbial cell membrane increased in the order of chemical pre-treatments (0.9% w/v physiological saline, 50 mM Tris HCl, pH 7.8, 50 mM Tris HCl, pH 7.8+50 mM EDTA). Thus, Tris-EDTA adjunct increased hydrophobicity of the membrane to bind more Congo red than other treatments. Virulence strains of Gram-positive bacteria may likely cause a more severe keratoconjunctivitis. Pigmentations favoured Gram-positive than negative probably because of the membrane differentiation. The effect of multiple subculturing on the Congo red binding ability of these bacteria remained inconclusive as less than half of the tested bacteria complied with our hypothesis though, all were expectedly affected, when pre-treated. The virulence of bacteria is pre-determined by the inherent plasmid factors but their expression is a function of the bacteria cells to bind organic dye. Therefore, physico-chemical treatments that promote hydrophobicity of the bacteria cell membrane are likely to reduce their virulence.
PDF Abstract XML References Citation
Received: April 06, 2015;
Accepted: May 23, 2015;
Published: June 30, 2015
How to cite this article
O.S. Folorunso, K.O. Amisu and B.F. Ogungbe, 2015. EDTA-Treated Cell Membrane and Multiple Subculturing Affect the Virulence of Enteropathogenic Bacteria. Research Journal of Microbiology, 10: 158-169.
URL: https://scialert.net/abstract/?doi=jm.2015.158.169
URL: https://scialert.net/abstract/?doi=jm.2015.158.169
INTRODUCTION
Enteropathogenic bacteria continually threaten human health due to the secretion of toxigenic proteins, part of which make them resistant to common antibiotics. The infections caused by Grams positive and negative bacteria are characterized with different levels of pathological symptoms (Chopra et al., 2008; Kinney, 2010; Proft and Baker, 2009; Tamma et al., 2012). A selection of these bacteria, like Enterobacter and Pseudomonas species, are opportunistic and only infect those with already suppressed immune defences (Berg et al., 2013; Healy et al., 2010; Ong et al., 2011). Contrarily, few of the Bacillaceae are non-pathogenic (Everest, 2007). The infections caused by this pathogen are expedited by a vast array of different virulence factors which not only enable evasion of host immune responses (Hong et al., 2009) but also facilitate adhesion, colonization, dissemination to other tissues and scavenging nutrients and minerals from the host ambience (Foster, 2005; Dinges et al., 2000). Reports suggested that hydrophobicity of the cells membrane and extracellularly expressed proteins and lipopolysaccharides (LPS) pre-set the pathogenicity and virulence of these organisms (Thomas and Elkinton, 2004; Molnar et al., 1994; Podschun and Ullmann, 1998; Magnusson, 1981) but there was no clear relationship between the integrity of the membrane with bacteria virulence.
Practically, multiple subculturing of pathogens may relegate virulence strain to avirulence (Haider et al., 1993). Similarly, chemical treatment of bacteria cell surface independently altered the hydrophobicity of the cellular membrane (Hale et al., 1985; Janda and Abbott, 1993) but reports are unable to reveal the interplay of these two factors with virulence ability of pathogenic bacteria. However, Sereny test has been used to assess the virulence of pathogens (Murayama et al., 1986; Sereny, 1957) and established that invasion and proliferation in epithelial cells were key features of pathogenic bacteria but this test may not corroborate the virulence and hydrophobicity of bacterial cell membrane.
Still, the pathogenic mechanisms of these bacteria appeared complex and unresolvable. Apart from the chromosomal and plasmid-based factors (Hale et al., 1985; Maurelli et al., 1984), many undetermined factors may play critical roles in the virulence of these pathogens. We posited that the virulence of a pathogen is a function of its ability to bind Congo red dye and to cause keratoconjunctivitis in the eyes of mouse (Sereny test); in addition, we aimed at relating the preponderant membrane protein secretion of pathogenic bacteria to their virulence. Nevertheless, not all secreted proteins are virulence factors.
The need for preliminary investigation of pathogenic virulence of bacteria is paramount. Therefore, using Congo red binding assay, we harnessed the interplay of multiple subculturing and EDTA-treated cell membrane alteration of bacteria on the virulence of eight different pathogenic bacteria.
MATERIALS AND METHODS
Experimental animals: Nine Albino rats of 6 weeks old were obtained from Animal Breeding Unit of the College of Medicine of the University of Lagos, Akoka Lagos State Nigeria. The animals, weighed 100-120 g, were maintained in 12 h light: 12 h dark at room temperature and humidity and kept in the animal house of the Department of Biochemistry, Lagos State University, Ojo, Lagos State Nigeria. The animals were allowed to acclimatize for one week in a plastic cage (2×2 ft.) at two per cage (male and female each). Certified commercial feed and water were given ad libitum. All animals were treated in accordance with the recommendations of the Ethical Committee on the Use and Management of Laboratory Animals, College of Medicine, University of Lagos, Nigeria as adopted from the 1996, Guide for the Care and Use of Laboratory Animals (Clark et al., 1997).
Microorganisms: All pure bacteria isolates used in this work were obtained from the Microbiology Laboratory of May Hospital Ilasamaja Lagos State and the Department of Microbiology, Lagos State University, Lagos State Nigeria. These microorganisms were made up of three Gram-positive bacteria: Staphylococcus aureus (isolated from septic wound and High Vaginal Swab, HVS), Streptococcus faecalis (isolated from patient suffering from Urinary Tract Infection, UTI) and Bacillus subtilis. Others were Gram-negative bacteria: Klebsiella pneumoniae (isolated from sputum sample), Enterobacter aerogenes, Pseudomonas aeruginosa, Salmonella typhimurium and Escherichia coli.
Bacteria were maintained on either MacConkey Agar (MCA) or Tryptic Soy Agar (TSA) (Oxoid) containing 0.6% w/v yeast extract.
Cell concentration using McFarland turbidity standards (Smibert and Krieg, 1994): We used spectrophotometric method to estimate the concentration of each of the bacteria used for Congo red binding assay. This involved adding 3.3 mL of 1% BaCl2 solution to 6.7 mL of 1% strong H2SO4 and the absorbance of this 10.0 mL solution at 600 nm was juxtaposed with suspension of bacteria cells in deionized water. The concentration (1×1010 CFU mL-1) of the bacteria was extracted according to the McFarland’s standard concentration.
Formation of pigmented bacteria colonies (Maurelli et al., 1984): We used Tryptic Soy Broth (TSB), which contained 0.6% w/v yeast extract, 1.5% w/v pure agar and 0.003% w/v Congo red to inoculate each bacterium at 37°C for 18 h. Then, the media were carefully examined for the formation of pigmented (PCr+) and non-pigmented (PCr‾) bacteria colonies.
Sereny test for microbial virulence: The virulence of bacteria was assessed in the eyes of nine Albino rats as described by Murayama et al. (1986) with little modifications. A colony from each of PCr+ and PCr‾ bacteria obtained after 18 h of growth was subcultured by layering on the TSA containing 0.6% yeast extract. Cells were collected with a sterile swab stick (Zhejiang Gongdong Medical Tech) and diluted with sterile normal saline (0.90% w/v) vials to a concentration of 3.0×108 CFU mL-1 and 20 μL of this suspension was carefully introduced just under the iris of the Albino rat right eye while the left eye was inoculated with physiological saline and served as control. Strains which produced keratoconjunctivitis within 72 h, were considered virulent. Symptoms and signs of keratoconjunctivitis in Albino rats infected with bacteria were observed within 72 h post-inoculation. The invasiveness of these bacteria was scored according to the following system (Cai et al., 2011):
| • | ‘-’ indicates lack of inflammation but an infection level 0 |
| • | ‘±’ indicates low levels of keratoconjunctivitis but an infection level 0.5 |
| • | ‘+’ indicates formation of slight conjunctival inflammation with eyelid edema and an infection level 1 |
| • | ‘2+’ indicates formation of infection, mild keratoconjunctivitis with eyelid edema, increased tear film evaporation and periocular hair loss and an infection level 2 |
| • | ‘3+’ indicates formation of infection with fully developed keratoconjunctivitis with eyelid swelling, periocular hair loss, conjunctival follicles and purulent discharge |
Congo red binding assay (Qadri et al., 1988; Ishiguro et al., 1985): With little modifications, this involved the use of sterilized TSB containing 0.6% w/v yeast extract. A 10 mL TSB was incubated with a colony of each bacterium at 37°C for 18 h. The cells were centrifuged at 6000 g for 10 min. The cell pellet was collected and washed with 0.05 M Phosphate Buffer Saline (PBS) (pH 7.4) and then re-centrifuged at 6000 g for another 10 min.
These cells at 1.0×1010 CFU mL-1 were then incubated for 10 min at 25°C in 3 mL 50 mM PBS containing 50 μg Congo red. The cells were removed by centrifugation at 16000 g for 50 min at 20°C. The filtrate (Congo red supernatant) was measured at 480 nm against cell-free Congo red solution. The percentage Congo red binding activity of each of the bacteria was calculated using the formula below:
Ao represents a cell-free Congo red assay, while A1 represents cell inoculated Congo red binding assay. Inferentially, the higher the hydrophobicity of the cell membrane, the higher the Congo red bound by the bacteria cells. Consequently, this will reduce the concentration of Congo red in the medium. Therefore, chemicals that do favour the removal of lipopolysaccharides (LPS) from the bacterial cell wall will favour the binding of Congo red. This method indirectly showed the amount of Congo red bound to the bacteria cells.
Pre-treated cells (Qadri et al., 1988): Each of the bacteria was subjected to three different chemical treatments and then examined for Congo red binding ability. Our conjecture was that multiple subculturing and/or chemical treatments could reduce the virulence/pathogenicity of enteric bacteria. Hence, each bacterium in TSA was repeatedly cultured eight times. The CRBA was carried out on the eighth inoculum and this was concurrently run with non-subcultured inoculum, as positive control so as to monitor the loss of pigmentation as formed earlier by the cells. The procedure involved the use of 10 mL TSB to culture a colony of each of the bacteria in shaker incubator at 37°C for 18 h. After 18 h, the cells were centrifuged at 6000 g for 10 min at room temperature. The cell pellet was divided into four equal parts after reconstitution with deionized water. Each part was washed three times with Solution A (deionized water as control), Solution B (0.90% w/v physiological saline), Solution C (50 mM Tris HCl buffer, pH 7.8) and Solution D (50 mM Tris HCl buffer, pH 7.8, which contained 50 mM EDTA) using rotary shaker (Labor Technik) at 60 rpm for 1 h at 20°C. Each cell was then centrifuged at 6000 g for 10 min and Congo red binding assay was carried out as earlier described. The bacteria were treated separately under non-subcultured and multiple subcultured following chemical treatments for each of the groups.
Protein secretion (Adamus and Romanowska, 1979): A colony of the microorganisms was inoculated into 5.0 mL of TSB in McCartney bottle and incubated between 18 and 24 h at 37°C with constant shaking at 250 rpm. The TSB was centrifuged at 12000 g and 10°C for 15 min. The supernatant was pipetted and filtered using 0.45 μm millipore (MCE HAWP 0.45 μm 047 mm 100/PK) and further confirmed as cell-free by re-inoculating it on Nutrient agar and incubating for 18 h.
Quantification of secreted protein (Bradford, 1976): The total amount of protein secreted by each of the bacteria was spectrophotometrically determined using Bradford method. Briefly, 5.0 mL of Bradford dye reagent was added to 1.0 mL of supernatant from each bacterium culture. The solution was mixed thoroughly and incubated at 37°C for 5 min. The absorbance was read at 595 nm against the reagent blank. Bovine Serum Albumin (BSA) at 0.1 mg mL-1 was used as standard protein.
Statistical analysis: All data used were presented as Mean±SEM. All experimental readings were read in duplicate. Bonferroni’s Multiple Comparison Test (BMCT) using one way analysis of variance (ANOVA) was adopted and the comparing means were considered significant at p<0.05.
RESULTS
Pigmentation of bacteria colonies: After 18 h, each plate of the bacteria was carefully examined for the formation of colonial pigment. The PCr+ was used to denote the formation of dense colonies with dark red centres as found in Staphylococcus aureus, Streptococcus faecalis and Klebsiella pneumoniae. This appeared as dense reddish mucous with varying colour intensities. The formation of whitish colonies characterized with extricated red colour as found in Bacillus subtilis, Enterobacter aerogenes, Pseudomonas aeruginosa, Salmonella typhimurium and Escherichia coli was represented as PCr‾. However, it was observed that prolonged incubation period of more than 30 h made the two ambiguous. Table 1 shows the bacteria that formed red pigmentation (PCr+) with Congo red assay and those that were negative to pigmentation (PCr‾).
Sereny test for microbial virulence: Bacteria strains that were positive to Congo red assay as a marker for virulence may likely be positive to Sereny test. The 20 μL of 3.0×108 CFU mL-1 was sufficient to cause symptoms of keratoconjunctivitis within 72 h of post-infection at the right eyes of each of the rats (Fig. 1). Only Staphylococcus aureus and Salmonella typhimurium were able to cause mild keratoconjunctivitis at the right while, we observed traces of symptoms in Streptococcus faecalis, Klebsiella pneumoniae, Pseudomonas aeruginosa and Bacillus subtilis.
Congo red binding activity: Figure 2a-i show the percentage Congo red binding affinity (pCRBA) of different bacteria under different chemical treatments in non-subculturing (NSC) and multiple subculturing (MSC) forms. It was envisaged that multiple subculturing and different chemical treatments of bacteria cell wall may independently reduce the pCRBA of each of the bacteria, a prognostic index of bacteria virulence.
Careful comparisons of these conditions generally confirmed that MSC had significant effects on the virulence of Staphylococcus aureus, Bacillus subtilis and Enterobacter aerogenes (p<0.0001, p<0.0001 and p<0.01, respectively, Fig. 2a, b, g and h) while other bacteria showed no significant difference at p<0.05.
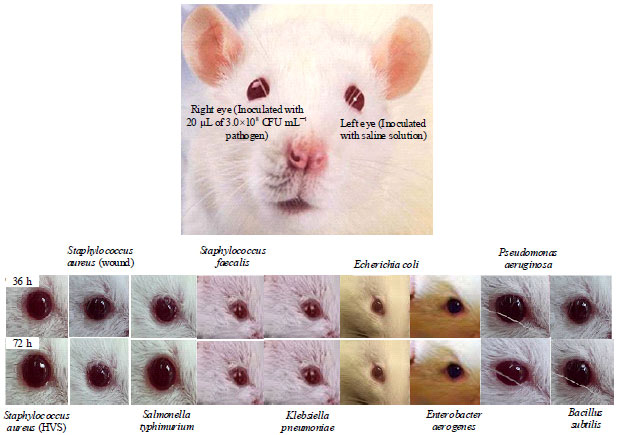 | |
| Fig. 1: | Formation of keratoconjunctivitis in the eyes of albino rats after 72 h of post-infection (hpi) |
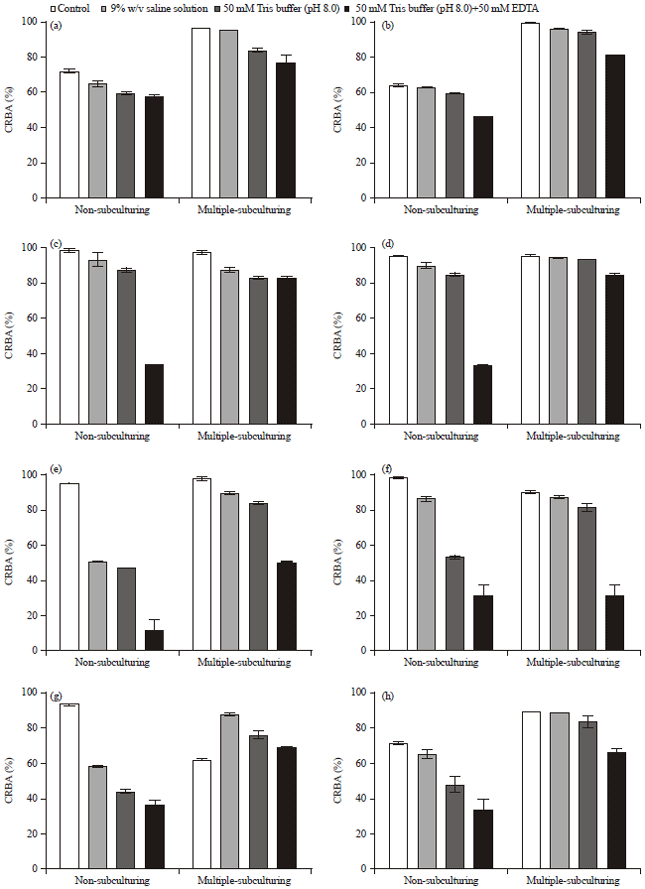 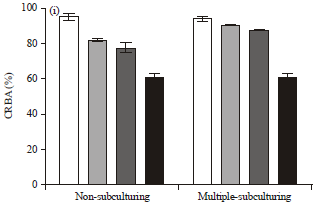 | |
| Fig. 2(a-i): | CRBA of (a) Staphylococcus aureus (HVS), (b) Staphylococcus aureus (Wound), (c) Streptococcus faecalis, (d) Klebsiella pneumoniae, (e) Salmonella typhimurium, (f) Escherichia coli, (g) Bacillus subtilis, (h) Enterobacter aerogenes and (i) Pseudomonas aeruginosa |
| Table 1: | Pigmentation and virulence assessment of the enteric bacteria |
 | |
| PCr+: Dense colonies with dark red centres (pigmented), PCr‾: Formation of whitish colonies devoid of affinity for red dye (nonpigmented), -: No inflammation but an infection level 0, ±: Low levels of keratoconjunctivitis but an infection level 0.5, +: Slight conjunctival inflammation with eyelid edema and an infection level 1 | |
Propinquity in the average pCRBA of Streptococcus faecalis, Klebsiella pneumoniae, Salmonella typhimurium, Escherichia coli and Pseudomonas aeruginosa in their NSC and MSC assays indicated that MSC adopted in this study was not enough to affect the virulence of these bacteria or another mechanisms undetermined this function. Since the binding of bacteria cells to Congo red is indirectly proportional to the intensity of the red dye in the medium, then the higher the binding the lower the intensity of the dye. A more virulent bacterium would bind more dye than the less virulent bacterium.
Effects of chemical treatments in NSC and MSC: We observed the effects of chemical treatments (0.90% w/v physiological saline, 50 mM Tris HCl buffer, pH 7.8 and 50 mM Tris HCl buffer, pH 7.8+50 mM EDTA) on the hydrophobicity of all the bacteria cell membrane. There were random effects of the chemical treatments on the hydrophobicity of the cell membrane. It was generally observed that treatment with 50 mM Tris HCl buffer pH 7.8+50 mM EDTA significantly affected the cell membrane as compared to other chemical treatments. The EDTA-treated cells mostly bound higher proportion of the Congo red than any other treatment and in this case recorded lower pCRBA.
| Table 2: | Significant differences in the pCRBA between NSC and MSC when subjected to different chemical treatments |
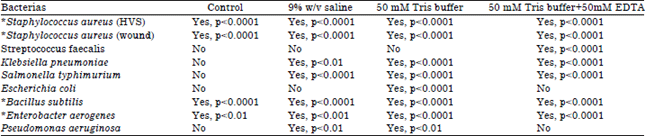 | |
| Mean differences are significant at p<0.05, *: Highly significant | |
When corresponding treated groups were compared (Table 2),*Staphylococcus aureus, *Bacillus subtilis and *Enterobacter aerogenes showed high significant impact of the chemical treatment on their ability to bind the dye; there were higher bindings in the NSC bacterial cells than the MSC cells. When saline, Tris and Tris-EDTA treatments were used on NSC and MSC, there was no difference in the CRBA of Streptococcus faecalis (saline and Tris) and Escherichia coli (saline and Tris-EDTA) and their control (Fig. 2c and f and Table 2).
In NSC, there was no significant difference (p>0.05) in the pCRBA of all the saline treated bacteria cells and the control cells (untreated) except in Salmonella typhimurium (50.68±0.37 vs. 94.80±0.30, p<0.001, Fig. 2e), Bacillus subtilis (57.96±0.30 vs. 92.84±0.43, p<0.0001, Fig. 2g) and Pseudomonas aeruginosa (81.82±0.92 vs. 95.35±1.96, p>0.01, Fig. 2i). Similarly, there was no significant difference (p<0.05) in the pCRBA of saline and Tris buffer treated groups except in Bacillus subtilis (57.96±0.30 vs. 43.77±0.92, p<0.01, Fig. 2g) and Escherichia coli (86.29±1.22 vs. 52.94±1.29, p<0.001, Fig. 2f). Tris buffer treated cells particularly showed lower pCRBA as compared to the control except in Streptococcus faecalis and Enterobacter aerogenes (87.21±1.29 vs. 98.16±1.22 and 48.13±4.59 vs. 71.88±1.16, respectively, p>0.05). This was an indication that, Tris buffer treated cells bound more dyes than the untreated control bacteria cells. There were significant differences (0.01<p<0.0001) between the Tris buffer+EDTA solution treated cells in all the isolates, when compared to the control; saline (except in Staphylococcus aureus (HVS), 58.30±0.54 vs. 65.23±1.71) and Tris buffer (except in Staphylococcus aureus (HVS), 58.30±0.54 vs. 59.90±0.80; Bacillus subtilis, 31.40±5.82 vs. 52.94±1.29 and Enterobacter aerogenes, 33.69±6.67 vs. 48.13±4.59).
In MSC, we observed significant differences similar to NSC in the effects of Tris buffer+EDTA against the control, saline and Tris buffer treated cells. The pCRBA of Tris+EDTA treated groups were generally lower than the control (except in Bacillus subtilis), saline (except in Streptococcus faecalis) and Tris buffer (except in Staphylococcus aureus (HVS), Streptococcus faecalis and Bacillus subtilis) (p>0.05). However, there were no significant differences in the Tris buffer treated cells when compared to the saline (except in Bacillus subtilis, 75.58±2.27 vs. 87.27±0.86, p<0.01) and the control except in Staphylococcus aureus (wound), p<0.01; Streptococcus faecalis, Salmonella typhimurium and Bacillus subtilis (p<0.001). Lastly, there was no significant difference in the control and saline treated cells except in Streptococcus faecalis, 97.0±1.41 vs. 87.15±1.59, p<0.01; Salmonella typhimurium, 97.61±1.16 vs. 89.66±0.80, p<0.01 and Bacillus subtilis, 61.82±0.61 vs. 87.27±0.86, p<0.0001.
DISCUSSION
We carried out Sereny test at the eyes of Albino rats using eight different enteric pathogenic bacteria. There was a formation of keratoconjunctivitis at the right eyes of the rat. We monitored this effect with physiological saline at the left eyes and there was no any trace of conjunctivitis.
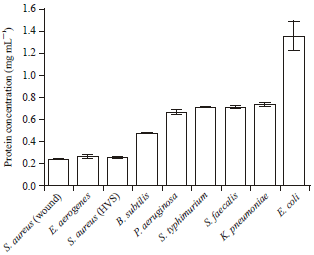 | |
| Fig. 3: | Total protein secreted by the pathogen when inoculated in TSB |
There was no transfer of infection from the right eye to the left showing that it was non-contagious. Though bacteria like Streptococcus faecalis, Klebsiella pneumoniae, Enterobacter aerogenes, Pseudomonas aeruginosa, Bacillus subtilis and Escherichia coli were negative to Sereny test, yet this may not be substituted for their virulence traits or perhaps 3.0×108 CFU mL-1 used in this work may not be sufficient to cause noticeable pathological change in the eyes of the rats. However, Staphylococcus aureus and Salmonella typhimurium showed noticeable conjunctival symptoms in the eyes of the tested rats (Table 1). Meanwhile, we may carefully suggest that Gram-positive may be more severe to cause keratoconjunctivitis in the eyes than the Gram-negative. We support this hypothesis on the fact that Staphylococcus species and Bacillus species are more of peripheral pathogens. Information from MedicineNet (Health and Medical Information) claimed that apart from eye disease causing viruses, Staphylococci and Streptococci were known to cause bacterial conjunctivitis. Again, Gram-positive bacteria do release exotoxins more regularly than endotoxin/lipopolysaccharide released by Gram-negative and this in turn may favour the formation of keratoconjunctivitis in the eyes of the rats more easily.
Gram-positive bacteria possess a thick cell wall containing many layers of peptidoglycan and teichoic acids. In contrast, Gram-negative bacteria have a relatively thin cell wall consisting of a few layers of peptidoglycan surrounded by a second lipid membrane containing lipopolysaccharides and lipoproteins. These factors predict the extent to which bacteria bind Congo red. Pigmentation favoured Gram-positive more than Gram-negative. We made this deduction because Gram-positive bacteria do engage in constitutive secretion of toxins than Gram-negative (Ray, 2010). Richard Friedrich Johannes Pfeiffer (1858-1945) had once said that exotoxins were usually released by bacteria into the surrounding, while endotoxins were toxins kept within the bacterial cell and released only after lysis (Parija, 2009). Other works have shown that LPS/endotoxins from Gram-negative are secreted as part of routine mechanisms in physiological membrane vesicle trafficking and this may also contain other virulence factors (Kulp and Kuehn, 2010). The association of the dye with the microbial cell membrane resided in its ability to interact with the membrane components. It binds in the increasing order from lipopolysaccharides, phospholipids and proteins. The hydrophobic portion of membrane protein has been found with greater affinity for organic dyes (Qadri et al., 1988). However, it is pertinent to know that not all secreted proteins are necessarily virulence factors, though they may bind dyes. We found that Escherichia coli secreted highest protein in our choice of medium but showed no comparable virulence (Fig. 1 and 3).
Disrupting membrane components could open way for increasing association of Congo red dye with the microbial membrane. Hydrophobicity resulted when the cellular membrane components are disengaged. This invariably increases the binding of dyes. We assumed that chemical treatment of the cell membrane of pathogenic bacteria vis-a-vis multiple subculturing could reduce the virulence of these bacteria. A more virulence bacteria would always bind more dye, when its membrane components are exposed. Meanwhile, certain bacterial plasmids have been found to be responsible for the expression of virulence factors and thus, Congo red binding ability (Daskaleros and Payne, 1986). Because these components are secreted as either endotoxins (mostly for the Gram-negative) or exotoxins (mostly for the Gram-positive) at the cell membrane, treating the peptidoglycan of the bacterial cell wall will allow them to bind the dye. We carried out this investigation and discovered that Tris+EDTA treated cells generally increased the hydrophobicity of the microbial cell membrane; hence, increasing affinity for Congo red. The higher the Congo red bound the lower the pCRBA. The Tris+EDTA interacts with the LPS and disengaged them from the cell membrane (Leive et al., 1968) thereby reducing membrane hydrophilicity. Report from Qadri et al. (1988) showed that isolated LPS did not bind Congo red but isolated outer membrane protein as well as Bovine Serum Albumin (BSA) bound more Congo red. When hydrophilicity of the cell membrane reduces, its hydrophobicity components will be exposed to bind dyes. Among the chemical adopted in our study, Tris+EDTA had the highest significant impact on the hydrophobicity exposure of the membrane, followed by Tris buffer and saline solutions. The reason been that EDTA is a chelating agent, causes increased cell surface permeability and membrane bulges (Alakomi et al., 2006).
We further speculated that multiple subculturing of pathogenic bacteria might reduce their virulence. The passaging of the cells reduced the efficiency of the virulence plasmids in them and this could cause poor expression of cellular components due to the rejuvenation period. In the work of Haider et al. (1993), multiple subculturing could result into the formation of avirulence strains from virulence. The level of subculturing may not be affirmative. We employed eight times subculturing passages in this work but multiple subculturing could only significantly reduce the binding of Congo red by Staphylococcus aureus, Bacillus subtilis and Enterobacter aerogenes, while, others showed no difference, when compared to non-subcultured forms. We may predict, not with certainty, that multiple subculturing may have more reducing effect on gram-positive than gram-negative to bind Congo red. The reason for this may not be clear but we attributed it to a more complex nature of the cell membrane of gram-positive bacteria.
Generally, there were close proximity in the pCRBA of the enteric bacteria probably, because they share similar pathway of toxin secretion. We also noted that not all pathogenic bacteria that were positive to pigmentation would be positive to Sereny test and that being negative to Sereny test was not alone to rationalize the non-virulence of bacteria. Hydrophobicity of the outer cell membrane of bacteria increases with the treatment of EDTA containing reagents and this increases the Congo red bound by such virulence cells. However, the virulence ability of bacteria resided in its intrinsic plasmid but its expression is directly associated with binding Congo red. Multiple passaging of these cells could be tantamount to avirulence but this may not rub off pathogenicity of the organism.
ACKNOWLEDGMENT
The authors wish to acknowledge Mr Ogunfiditimi Kehinde at the Microbiology Laboratory, May Hospital, Ilasamaja, Lagos State Nigeria for his effort to provide us clinical pathogenic samples of bacteria.
REFERENCES
- Adamus, G. and E. Romanowska, 1979. Outer membrane proteins of Shigella sonnei. II. Comparative studies on virulent and avirulent strains of phase I. Archivum Immunologiae at Therapiae Experimentalis, 28: 553-558.
Direct Link - Alakomi, H.L., A. Paananen, M.L. Suihko, I.M. Helander and M. Saarela, 2006. Weakening effect of cell permeabilizers on gram-negative bacteria causing biodeterioration. Applied Environ. Microbiol., 72: 4695-4703.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cai, X., J. Zhang, M. Chen, Y. Wu and X. Wang et al., 2011. The effect of the potential PhoQ histidine kinase inhibitors on Shigella flexneri virulence. PloS One, Vol. 6.
CrossRef - Clark, J.D., G.F. Gebhart, J.C. Gonder, M.E. Keeling and D.F. Kohn, 1997. The 1996 guide for the care and use of laboratory animals. ILAR J., 38: 41-48.
CrossRefDirect Link - Daskaleros, P.A. and S.M. Payne, 1986. Characterization of Shigella flexneri sequences encoding congo red binding (crb): Conservation of multiple crb sequences and role of IS1 in loss of the Crb+ phenotype. Infect. Immun., 54: 435-443.
Direct Link - Dinges, M.M., P.M. Orwin and P.M. Schlievert, 2000. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev., 13: 16-34.
CrossRefPubMedDirect Link - Haider, K., A. Hossain, C. Wanke, F. Qadri, S. Ali and S. Nahar, 1993. Production of mucinase and neuraminidase and binding of Shigella to intestinal mucin. J. Diarrhoeal Dis. Res., 11: 88-92.
PubMedDirect Link - Hale, T.L., E.V. Oaks and S.B. Formal, 1985. Identification and antigenic characterization of virulence-associated, plasmid encoded proteins of Shigella spp. and enteroinvasive Escherichia coli. Infect. Immun., 50: 620-629.
PubMed - Healy, B., S. Cooney, S. O'Brien, C. Iversen and P. Whyte et al., 2010. Cronobacter (Enterobacter sakazakii): An opportunistic foodborne pathogen. Foodborne Pathogens Dis., 7: 339-350.
CrossRefDirect Link - Hong, H.A., R. Khaneja, N.M.K. Tam, A. Cazzato and S. Tan et al., 2009. Bacillus subtilis isolated from the human gastrointestinal tract. Res. Microbiol., 160: 134-143.
CrossRefPubMedDirect Link - Ishiguro, E.E., T. Ainsworth and W.W. Kay, 1985. Congo red agar, a differential medium for Aeromonas salmonicida, detects the presence of the cell surface protein array involved in virulence. J. Bacteriol., 164: 1233-1237.
Direct Link - Janda, J.M. and S.L. Abbott, 1993. Microbial hydrophobicity-the sticky end of things. Clin. Microbiol. Newslett., 15: 9-12.
CrossRef - Kinney, K.K., 2010. Treatment of infections caused by antimicrobial-resistant gram-positive bacteria. Am. J. Med. Sci., 340: 209-217.
CrossRefDirect Link - Kulp, A. and M.J. Kuehn, 2010. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol., 64: 163-184.
CrossRefDirect Link - Leive, L., V.K. Shovlin and S.E. Mergenhagen, 1968. Physical, chemical and immunological properties of lipopolysaccharide released from Escherichia coli by ethylenediaminetetraacetate. J. Biol. Chem., 243: 6384-6391.
Direct Link - Magnusson, K.E., 1981. Hydrophobic interaction-a mechanism of bacterial binding. Scand. J. Infect. Dis. Suppl., 33: 32-36.
Direct Link - Maurelli, A.T., B. Blackmon and R. Curtiss, 1984. Loss of pigmentation in Shigella flexneri 2a is correlated with loss of virulence and virulence-associated plasmid. Infect. Immun., 43: 397-401.
Direct Link - Molnar, C., Z. Hevessy, F. Rozgonyi and C.G. Gemmell, 1994. Pathogenicity and virulence of coagulase negative Staphylococci in relation to adherence, hydrophobicity and toxin production in vitro. J. Clin. Pathol., 47: 743-748.
CrossRefDirect Link - Murayama, S.Y., T. Sakai, S. Makino, T. Kurata, C. Sasakawa and M. Yoshikawa, 1986. The use of mice in the Sereny test as a virulence assay of Shigellae and enteroinvasive Escherichia coli. Infect. Immun., 51: 696-698.
Direct Link - Ong, D.S., I.P. Jongerden, A.G. Buiting, M.A. Leverstein-van Hall, B. Speelberg, J. Kesecioglu and M.J. Bonten, 2011. Antibiotic exposure and resistance development in Pseudomonas aeruginosa and Enterobacter species in intensive care units. Crit. Care Med., 39: 2458-2463.
CrossRefDirect Link - Podschun, R. and U. Ullmann, 1998. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods and pathogenicity factors. Clin. Microbiol. Rev., 11: 589-603.
CrossRefPubMedDirect Link - Proft, T. and E.N. Baker, 2009. Pili in Gram-negative and Gram-positive bacteria-structure, assembly and their role in disease. Cell. Mol. Life Sci., 66: 613-635.
CrossRefDirect Link - Qadri, F., S.A. Hossain, I. Ciznar, K. Haider, A. Ljungh, T. Wadstrom and D.A. Sack, 1988. Congo red binding and salt aggregation as indicators of virulence in Shigella species. J. Clin. Microbiol., 26: 1343-1348.
Direct Link - Tamma, P.D., S.E. Cosgrove and L.L. Maragakis, 2012. Combination therapy for treatment of infections with gram-negative bacteria. Clin. Microbiol. Rev., 25: 450-470.
CrossRefPubMedDirect Link - Thomas, S.R. and J.S. Elkinton, 2004. Pathogenicity and virulence. J. Invertebrate Pathol., 85: 146-151.
CrossRefDirect Link