
Widanarni
Department of Aquaculture, Faculty of Fisheries and Marine Science, Bogor Agricultural University, Jalan Agatis, Dramaga Campus, Bogor, West Java, 16680, Indonesia
Tita Nopitawati
Study Program of Aquaculture, Graduate School, Bogor Agricultural University, Indonesia
Dedi Jusadi
Department of Aquaculture, Faculty of Fisheries and Marine Science, Bogor Agricultural University, Jalan Agatis, Dramaga Campus, Bogor, West Java, 16680, Indonesia
Research Journal of Microbiology
Year: 2015 | Volume: 10 | Issue: 4 | Page No.: 145-157







ABSTRACT
This study aimed to get the probiotics bacteria from the gastrointestinal tract which had protease, amylase and lipase activities to improve growth performances of Pacific white shrimp (Litopenaeus vannamei). The probiotics were isolated from the gastrointestinal tract of Pacific white shrimp. The in vitro assays were carried out to isolate probiotics bacteria and selected the candidates based on digestive enzymes production (protease, amylase and lipase), resistance to gastric acid and bile salts, rapid bacterial growth phase, antagonistic activity against pathogenic bacteria, the attachment ability and pathogenicity to Pacific white shrimp. The selected isolates were then tested in vivo to observe their ability to improve growth performances of white shrimp. Five treatments with three replications consisted of control (feed without addition of any probiotic) and addition of four selected probiotics. Probiotics in a dose of 1% (v/w) feed were added every day for 60 days. Based on the results of in vitro selection, there were four selected isolates then applied to white shrimp including K9, M2, Z3 and S3. These four isolates have the highest ability in protein hydrolysis (K9), starch hydrolysis (M2), lipid hydrolysis (Z3) and protein, starch and lipid hydrolysis (S3). In addition, these isolates were also able to survive in acidic and alkaline condition, had antagonistic activity against pathogenic bacteria (V. harveyi), had the ability to adhere and not pathogenic for Pacific white shrimp. The feeding trial showed that isolate K9 showed the best results in improving protein digestibility (79.93±1.37%), protein retention (46.97±3.27%) and growth of Pacific white shrimp (1.09±0.01%).
PDF Abstract XML References Citation
Received: April 22, 2015;
Accepted: June 03, 2015;
Published: June 30, 2015
How to cite this article
Widanarni, Tita Nopitawati and Dedi Jusadi, 2015. Screening of Probiotic Bacteria Candidates from Gastrointestinal Tract of Pacific White Shrimp Litopenaeus vannamei and their Effects on the Growth Performances. Research Journal of Microbiology, 10: 145-157.
URL: https://scialert.net/abstract/?doi=jm.2015.145.157
URL: https://scialert.net/abstract/?doi=jm.2015.145.157
INTRODUCTION
The Pacific white shrimp (Litopenaeus vannamei) is one of the important economic value of fisheries commodities. Indonesia is one of the largest Pacific white shrimp exporting country in the world beside Ecuador, Thailand, Vietnam, China, India and Malaysia (FAO., 2013). Therefore, intensive farming system is conducted by using a high quantity of feed in order to increase the production of Pacific white shrimp. Feed is the energy source that holds 40-89% in the cost of aquaculture production (Suprayudi, 2010). On the other hand, the feed is not fully digested but it is released as waste in the form of feces. One strategy that can be developed is by focusing on the use of probiotic bacteria that give some benefits to the host by improving the digestibility, immune response and inhibit the growth of pathogenic microorganisms (Verschuere et al., 2000; Vine, 2004; Yanbo and Zirong, 2006; Gatesoupe, 2007; Salem et al., 2010; Elghandour et al., 2015; Puniya et al., 2015).
The presence of probiotic bacteria in the digestive tract can produce exogenous enzymes such as amylase, protease, lipase and cellulase which can increase the activity of the digestive tract enzymes and feed digestibility in shrimp (Sahu et al., 2008; Wang et al., 2008). The existence of these exogenous enzymes will help the endogenous enzymes in hydrolyzing feed nutrients to break down long chains of macromolecules such as carbohydrates, proteins and fats into simpler molecules, so they will be easier to be digested and absorbed by the intestine.
The use of commercial probiotics in aquatic organisms are often less effective because it is isolated from non-fish sources, which can not be able to survive in a high density in the intestines during the growing phase of fish (Moriaryty, 1996). Bacteria isolated from the host is expected to give a better effect because it can adhere to the host’s gut and adapt well to compete with the pathogens in getting nutrients (O'Sullivan, 2001). Therefore, probiotics isolation from the gastrointestinal tract of the host is the best approach to obtain the probiotic candidates. This study aimed to get the probiotic bacteria from the gastrointestinal tract which had protease, amylase and lipase activities to improve feed digestibility and growth performances of Pacific white shrimp.
MATERIALS AND METHODS
Isolation of probiotic bacteria candidates: The probiotic bacteria candidates were isolated from the gastrointestinal tract of Pacific white shrimp. Pacific white shrimps used in this study were obtained from Center of Aquaculture Production Business Service (CAPBS) Karawang, West Java, Indonesia. The gastrointestinal tract was removed from the shrimp with body weight of 10-15 g through aseptically disection. The gastrointestinal tract and its material inside will be the source for the next assays of probiotic bacteria candidates selection. The shrimp gastrointestinal tract samples were weighed as much as 1 g, then ground aseptically and diluted into 9 mL sterile physiological solution (NaCl 0.85%). The dilution then were spread onto Sea Water Complete (SWC) agar (1.25 g bacto peptone, 0.25 g yeast extract, 3 mL glycerol, 750 mL seawater, 250 mL distilled water and 30 g bactoagar), then incubated at 29°C for 24 h. Colonies with different morphology (colour and shape) were separated and re-cultured onto SWC agar. The colonies were purified several times until singular and uniform bacterial colonies were obtained.
Selection of probiotic bacteria candidates
Proteolytic, lipolytic and amylolytic properties test: Each probiotic bacteria isolates were tested for the proteolytic, lipolytic and amylolytic properties through the hydrolysis test of casein, fat and starch. The samples were grown on SWC agar +2% skim milk for proteolytic property, SWC agar +2% olive oil for lipolytic property and SWC agar +2% starch for amylolytic property. The samples were incubated at 29°C for 24-48 h. Proteolytic property was observed by measuring the clear zone around the colony after incubation. The amylolytic property was observed by measuring the clear zone around the colony after incubation by adding 1% Potassium Iodide (KI) while, the lipolytic property was shown by the bright green color around the colony after incubation and by saturated CuSO4. All isolates which have at least one digestive enzymatic property, will be subjected for the subsequent test (gastric acid and bile salts resistance, bacterial growth test, antagonistic test to the pathogen, adhesion test, pathogenicity test and also growth and digestibility test through feeding trial in Pacific white shrimp).
Resistance to gastric acid and bile salts test: The resistance test to gastric acid and bile salts was performed according to the method by Ngatirah et al. (2000). The probiotic bacteria candidates in amount of 1 mL was inoculated into tubes containing 9 mL SWC with pH 2.5 (the pH was obtained by the addition of HCl) and pH 7.5 (the pH was obtained by the addition of NaOH) and then incubated at 29°C. Observations were conducted at 2, 4, 6 and 8 h after inoculation and the number of bacteria counted by total plate count method. Resistance to gastric acid and bile salts is determined by the difference of bacterial number between the controls (the media with a pH of 7.0) and treatments. The smaller the difference, the more resistant to gastric acid and bile salts.
Bacterial growth test: This test was done to determine the probiotic bacteria candidates growth phase, especially the time to reach the exponential phase of each candidates. Fresh culture of each candidate was taken as much as 10 mL and inoculated into 90 mL SWC broth then incubated in water-bath shaker at 29°C with a speed of 140 rpm. Bacterial growth was monitored every 2 h by measuring Optical Density (OD) using a spectrophotometer with a wavelength of 620 nm (Hadioetomo, 1990).
Test of antagonistic activity to the pathogen: The pathogenic bacteria used were Vibrio harveyi that are pathogenic to the shrimp. The fresh culture of probiotic bacteria candidates in amount of 0.1 mL diluted in 0.9 mL physiological solution to obtain a suspension of probiotic candidates with a density of 106 CFU mL-1. Therefore, 0.1 mL of the pathogen solution with the same density was spread over SWC agar. Sterile paper disc was dipped into the probiotic candidates suspension and placed on the media that has been spread with the pathogen. Samples were incubated at 29°C for 24 h. Antagonistic activity was observed by measuring the inhibition zone diameter around the paper disc.
Bacterial adhesion test: Adhesion or attachment test was performed according to the method by Dewanti and Wong (1995). Sterile steel plate was placed in a standing position in the Erlenmeyer flask containing 250 mL SWC broth, which had been inoculated with 1 mL fresh culture of probiotic candidates. Samples were incubated in a water bath shaker for 24 h at a temperature of 29°C. After that, the surface of the steel plate was swab thoroughly. The swab was then inserted into the tubes containing 10 mL phosphate buffer and vortexed for 1 min. Furthermore, the bacterial population was calculated by total plate count method and stated in CFU mm-2.
Pathogenicity test of probiotic bacteria candidates: This test was performed by injecting a candidate probiotic bacterial suspension via intramuscular route to the shrimp with a concentration of 106 CFU mL-1 in amount of 0.1 mL per individual. The control was used as the comparison by injecting physiological solution to the shrimp. The shrimp were reared for 7 days and the observed parameter was the survival rate.
In vivo assays of probiotic bacteria candidates through feeding trial: This study used a Completely Randomized Design (CRD) which consisted of five treatments with three replications, namely K (feed without addition of probiotic candidate), M2 (feed plus the best amylolytic probiotic candidate), Z3 (feed plus the best lipolytic probiotic candidate), K9 (feed plus the best proteolytic probiotic candidate) and S3 (feed plus the best amylolytic, proteolytic and lipolytic probiotic candidate). The probiotic candidates used in the feeding trial selected based on in vitro assays results especially their higher digestive enzymes properties than the other candidates and also their capability to survive in acidic and alkaline condition. The selected probiotic candidates were inoculated into SWC broth and incubated in a water-bath shaker at a temperature of 29°C until reaching exponential phase of each probiotic candidate. The fresh culture were centrifuged at 5000 rpm for 15 min and rinsed twice with sterile PBS (Phosphate Buffer Saline). The probiotic bacteria candidates solution were then mixed into a commercial diet with 32% protein content. The dose probiotic candidates used was 1% (Wang, 2007) then added to the feed by spraying thoroughly using a syringe and added 2% egg yolk as a binder.
The shrimp were reared in a plastic tank with a volume of 90 L (30 individuals per tank), with initial weight of 8.35±0.16 g and length of 10.14±0.21 cm. During treatment, the shrimp were given feed containing probiotic candidates and without probiotic by ad satiation at 06.00, 10:00, 14:00, 18:00 and 22:00 Western Indonesia Time. The amount of feed given during feeding trial were noted and the total feed calculated at the end of the feeding trial. The feeding trial was conducted for 60 days. Water quality was maintained by siphoning water in the rearing tanks every morning and water replacement was done every three days.
Measurement of growth performances parameters and enzymes activity: The growth performances parameters measured were Survival Rate (SR), Specific Growth Rate (SGR), feed conversion according to Huisman (1987), protein and fat retention were calculated according to the method by Takeuchi (1988). Enzyme activity assays were performed according to the method developed by Bergmeyer and Grassi (1983) using 6 shrimp samples in each rearing tank.
Digestibility test: The test feed before being mixed with the probiotics were mashed into powder and then added 0.6% Cr2O3 as a marker and 20 g kg-1 feed of CMC as a binder. The feed powder was repelleted and dried using an oven. The feed were given to the shrimp for a week and the feces collection was conducted on the 7th day. Furthermore, the analysis of Cr2O3 content in the feces was conducted (Takeuchi, 1988).
Statistical analysis: The data of isolation and selection of probiotic bacteria candidates within in vitro assays and enzymes activity of the shrimp digestive tract within feeding trial were analyzed descriptively, while the growth performances and digestibility data were analyzed using analysis of variance with a confidence interval of 95%, followed by the Duncan test.
RESULTS
Isolation of probiotic bacteria candidates: The result of isolation of probiotic bacteria candidates from the digestive tract of Pacific white shrimp were obtained 30 isolates to be selected their enzymatic properties. The result of proteolytic, amylolytic and lipolytic properties of probiotic bacteria candidates were obtained 12 isolates that showed hydrolysis of protein, starch and fat (Fig. 1). Those isolates will be subjected to the subsequent in vitro assays. The clear zone diameters of protein hydrolysis ranged from 1-20 mm. The best isolates in producing protease were K9, K19 and Z1 with the same hydrolysis zone diameters (20 mm). Isolates with the best lipolytic property were Z3 and Z5 (9; 12 mm). Isolates which had the best amylolytic property were M1 and M2 (15; 15 mm).
Resistance to gastric acid and bile salts: The result of resistance to gastric acid and bile salts test is presented in Fig. 2a-b. The initial number of probiotic candidates in normal, acidic and alkaline pH (2 h) was in range of 1.1×107-2.9×108 CFU mL-1. Isolates S3 and K9 had the smallest difference of the log bacterial number to the control that meant that these isolates had the ability to live in acidic and alkaline pH than other isolates.
Bacterial growth phase: The observation of Optical Density (OD) value resulted each isolate bacterial growth curves that had a varying growth (Fig. 3). Exponential phase of all isolates occurred at the 16th and 18th.
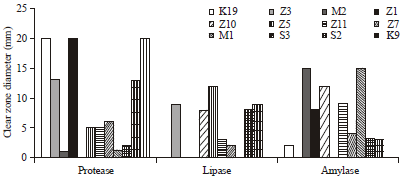 | |
| Fig. 1: | Clear zone diameter of proteolytic, lipolytic and amylolytic bacteria isolates |
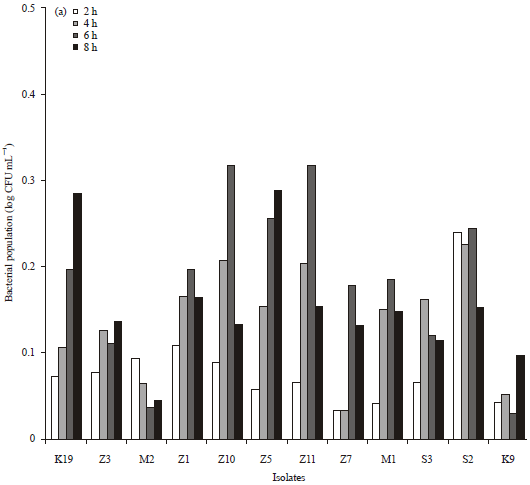 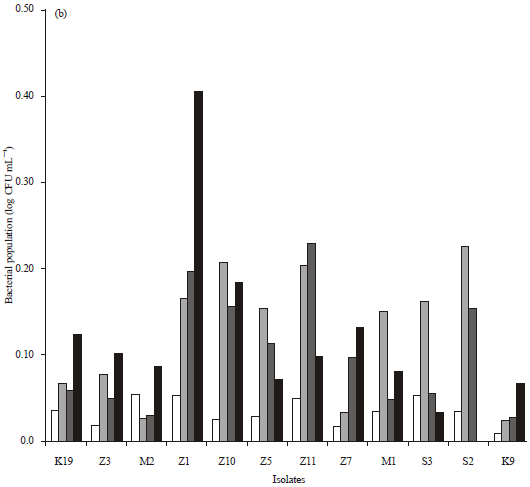 | |
| Fig. 2(a-b): | Differences of bacterial number in (a) pH 2.5 with normal pH and (b) pH 7.5 with normal pH |
Antagonistic activity to pathogen bacteria: The result of inhibition zone of probiotic bacteria candidates against pathogenic V. harveyi are presented in Fig. 4. All probiotic candidates except Z10 isolate were able to suppress the growth of V. harveyi with inhibition zones ranged from 2-13 mm. Best inhibition zone was produced by S2 isolate with a diameter of 13 mm.
Adhesion of probiotic bacteria candidates: The result of probiotic bacteria candidates adhesion is shown in Fig. 5. The K9 isolate had the highest bacterial number (14.222×103 CFU mm-2) compared to other isolates which meant that this isolate was able to attach well to the substrate.
Pathogenicity test of probiotic bacteria candidates: The result of pathogenicity test showed that all tested probiotic bacteria candidates were not pathogenic. During the pathogenicity test, the survival rate of the shrimp on the probiotic treatments and control were 100%.
Growth performances, digestibility and digestive tract enzymes activity of the shrimp: The result showed that the best growth of shrimp was shown in treatment K9 (1.09±0.01%) that was significantly different (p<0.05) with all treatments. The better feed conversion was obtained at treatment S3 (1.93±0.036%) that was not significantly different (p>0.05) with K9 (1.83±0.09%) but it was significantly different (p<0.05) with treatment M2, Z3 and control. The best total digestibility was obtained at treatment S3 (78.55±0.76%) that was significantly different (p<0.05) with control, M2 and K9.
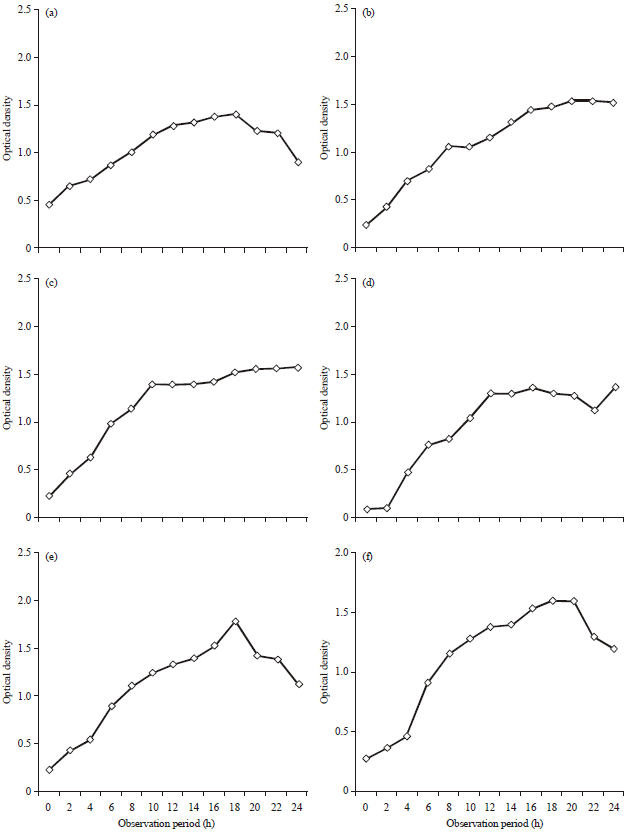 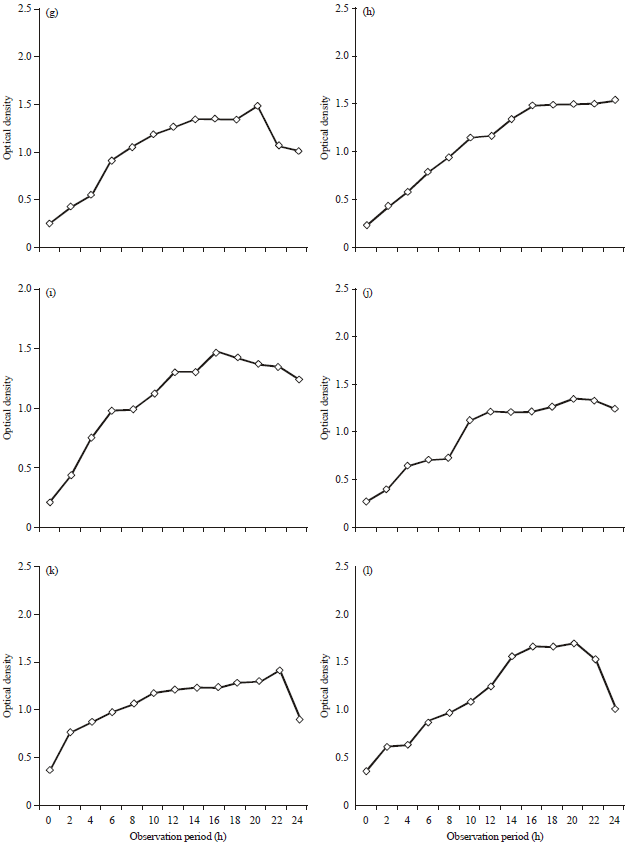 | |
| Fig. 3(a-l): | Optical density curves of probiotic bacteria candidates, (a) M1, (b) K9, (c) S2, (d) K19 (e) Z10, (f) Z11, (g) Z5, (h) S3, (i) M2, (j) Z3, (k) Z7 and (l) Z1 |
The higher protein digestibilities were obtained on treatment K9 and S3 (79.93±1.37; 79.27±1.03%) that were significantly different (p<0.05) with control. The survival rate in probiotic treatments ranged from 95.56-100% that were significantly different (p<0.05) with the control (81.11%) (Table 1).
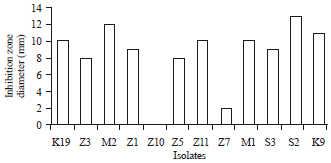 | |
| Fig. 4: | Inhibition zone diameter of probiotic bacteria candidates to Vibrio harveyi |
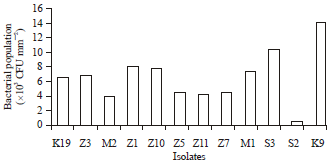 | |
| Fig. 5: | Result of adhesion test of bacteria isolates on stainless steel plates |
| Table 1: | Specific growth rate, survival rate, feed conversion, protein retention, total digestibility and protein digestibility of Pacific white shrimp |
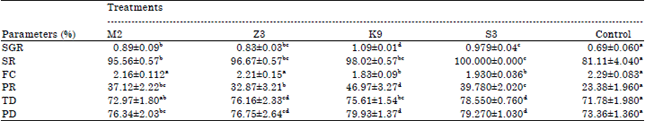 | |
| Different letters in the same row indicates significantly different results (p<0.05), SGR: Specific growth rate, SR: Survival rate, FC: Feed conversion, PR: Protein retention, TD: Total digestibility and PD: Protein digestibility | |
| Table 2: | Protease, lipase and amylase enzyme activity in the digestive tract of Pacific white shrimp |
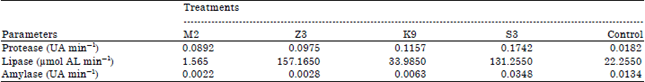 | |
The enzymes activity in the digestive tract of shrimp in probiotic treatment showed higher values than control. The higher protease activity in the digestive tract of shrimp were obtained on treatment K9 and S3 (0.1157; 0.1742 UA min-1). Treatment S3 also resulted in the highest amylase activity (0.0348 UA min-1) while, the highest lipase activity was obtained at treatment Z3 (157.165 μmol AL min-1) (Table 2).
DISCUSSION
The in vitro assays were carried out to isolate probiotic bacteria and selected the candidates based on several characteristics such as digestive enzymes production (protease, amylase and lipase), resistance to gastric acid and bile salts, rapid bacterial growth phase, antagonistic activity against pathogenic bacteria, the attachment ability and pathogenicity. Based on the results of in vitro selection, there was obtained four isolates applied for feeding trial to Pacific white shrimp including K9, M2, Z3 and S3. These four isolates have the highest ability in protein hydrolysis (K9), starch hydrolysis (M2), lipid hydrolysis (Z3) and protein, starch and lipid hydrolysis (S3). In addition, these isolates were also able to survive in acidic and alkaline condition, had antagonistic activity against pathogenic bacteria (V. harveyi), had the ability to adhere and not pathogenic for Pacific white shrimp. The presence of a clear zone produced by the probiotic bacteria candidate in the test of protein, starch and fat hydrolysis showed that the bacteria are able to hydrolyze macromolecules as a source of energy for their life. The hydrolysis ability was obtained from the presence of extracellular enzymes secreted by the bacteria. The ability of bacterial isolates to produce extracellular enzymes such as protease, lipase and amylase become a factor in determining a probiotic candidate. Some study showed that the ability of bacteria to produce extracellular enzymes such as protease, lipase and amylase can increase the digestibility of the host (Balcazar et al., 2006). Observation of the growth phase of bacteria is required to obtain growth curve of an isolate that is useful to determine the best time to harvest the cells. Furthermore, tolerance to gastric acid and bile salts is an essential condition for the probiotic candidate (Kesarcodi-Watson et al., 2008), because when probiotics enter the aquatic organism body, they will through the stomach which has acidic condition and meet bile salts, which has alkaline condition when through the intestine. The other important requirement in probiotics selection is having antagonistic activity that is able to suppress the growth of pathogenic bacteria in the digestive tract of shrimp. This can occur through local and systemic immune stimulation, nutrient and attachment location competition (Verschuere et al., 2000; Irianto, 2003). In addition, the candidate must be safe to its host, so when it is applied, it will not cause disease symptoms or death in the host. The characterization of probiotic candidates is necessary for quality control and patent but it is not a test that must be done in probiotic selection (Gomez-Gil et al., 2000), in which the identification of probiotic candidates in this study have not been done yet.
The in vivo trials are required to validate the effectiveness of probiotic (Vine et al., 2004) after getting in vitro trials results. The feeding trial in this study aimed to know the ability of all probiotic candidates to improve growth performances in Pacific white shrimp by influencing its metabolic activities. The highest growth rate of Pacific white shrimp was obtained on treatment K9 with the better feed conversion were on treatment K9 and S3. This was presumably due to the addition of probiotics through feed could improve the digestibility of shrimp. The Pacific white shrimp have a higher preference for utilizing protein as a source of energy than carbohydrates and fats. The Pacific white shrimp is also known requiring higher protein requirement than fresh water fish for growth (Watanabe, 1988). It was thought to be one of the factors that led treatment K9 (addition of best proteolytic probiotic candidate) and S3 (addition of best proteolytic, amylolytic and lipolytic probiotic candidate) had better growth and feed conversion than other treatments. The more protein that is able to be digested, the more feed is converted into body protein that is reflected in protein retention, growth and feed conversion in Pacific white shrimp. This is due to probiotics can enhance the activity of exogenous enzymes that can increase the digestibility of the feed, so that digestion of feed nutrients will increase and growth of shrimp will also increase (Chandran et al., 2014). This was in line with the value of digestive tract enzymes activity in Pacific white shrimp treated probiotics that produced a higher value than the control. Previous study also showed that administration of L. plantarum and B. subtilis could improve the growth of L. vannamei (Kongnum and Hongpattarakere, 2012; Li et al., 2009; Wang, 2007). Zokaeifar et al. (2012) stated that the addition of B. subtilis to the feed increased the activity of protease, amylase, total protein and white shrimp growth compared to control.
The result showed that administration of probiotics also produced high survival on Pacific white shrimp. The higher survival rates of probiotics treated shrimp than control were suspected due to probiotics could improve the immune system of shrimp. The increasing survival in probiotic treatments could also occur due to antimicrobial compounds produced by probiotic bacteria can inhibit pathogenic bacteria that allegedly contained in the shrimp or their living medium. This was in line with the in vitro assays results, which showed that these four probiotics isolates were able to inhibit the growth of pathogenic V. harveyi. Some of the previous studies also showed that the administration of probiotics could improve the survival of shrimp (Widanarni et al., 2003; Liu et al., 2010; Nurhayati et al., 2015).
CONCLUSION
The screening of probiotic bacteria candidates from the gastrointestinal tract of Pacific white shrimp resulted four isolates, which were potential as probiotic candidates namely K9 as the best proteolytic isolate, Z3 as the best lipolytic isolate, M2 as the best amylolytic isolate and S3 as the best isolate of those three enzymatic activity. Isolate K9 showed the best results in improving protein digestibility of the feed, protein retention and growth of Pacific white shrimp.
ACKNOWLEDGMENTS
The authors would like to acknowledge to Diah Ayu Satyari Utami, SPi, MSi for help and support in preparing this manuscript.
REFERENCES
- Balcazar, J.L., I. de Blas, I. Ruiz-Zarzuela, D. Cunningham, D. Vendrell and J.L. Muzquiz, 2006. The role of probiotics in aquaculture. Vet. Microbiol., 114: 173-186.
CrossRefPubMedDirect Link - Dewanti, R. and A.C.L. Wong, 1995. Influence of culture conditions on biofilm formation by Escherichia coli O157:H7. Int. J. Food Microbiol., 26: 147-164.
CrossRefPubMedDirect Link - Elghandour, M.M.Y., A.Z.M. Salem, J.S.M. Castaneda, L.M. Camacho, A.E. Kholif and J.C.V. Chagoya, 2015. Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. J. Integr. Agric., 14: 526-533.
CrossRefDirect Link - Gomez-Gil, B., A. Roque and J.F. Turnbull, 2000. The use and selection of probiotic bacteria for use in the culture of larval aquatic organisms. Aquaculture, 191: 259-270.
CrossRefDirect Link - Gatesoupe, F.J., 2007. Live yeasts in the gut: Natural occurrence, dietary introduction and their effects on fish health and development. Aquaculture, 267: 20-30.
CrossRefDirect Link - Kesarcodi-Watson, A., H. Kaspar, M.J. Lategan and L. Gibson, 2008. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture, 274: 1-14.
CrossRefDirect Link - Kongnum, K. and T. Hongpattarakere, 2012. Effect of Lactobacillus plantarum isolated from digestive tract of wild shrimp on growth and survival of white shrimp (Litopenaeus vannamei) challenged with Vibrio harveyi. Fish Shellfish Immunol., 32: 170-177.
CrossRefDirect Link - Li, J., B. Tan and K. Mai, 2009. Dietary probiotic Bacillus OJ and isomaltooligosaccharides influence the intestine microbial populations, immune responses and resistance to white spot syndrome virus in shrimp (Litopenaeus vannamei). Aquaculture, 291: 35-40.
CrossRefDirect Link - Liu, K.F., C.H. Chiu, Y.L. Shiu, W. Cheng and C.H. Liu, 2010. Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol., 28: 837-844.
CrossRefDirect Link - Moriaryty, D.J.W., 1996. Microbial biotechnology: A key ingredient for sustainable aquaculture. Infofish Int., 4: 29-33.
Direct Link - Chandran, M.N., P. Iyapparaj, S. Moveendhan, R. Ramasubburayan, S. Prakash, G. Immanuel and A. Palavesam, 2014. Influence of probiotic bacterium Bacillus cereus isolated from the gut of wild shrimp Penaeus monodon in turn as a potent growth promoter and immune enhancer in P. monodon. Fish Shellfish Immunol., 36: 38-45.
CrossRefDirect Link - Nurhayati, D., Widanarni and M. Yuhana, 2015. Dietary synbiotic influence on the growth performances and immune responses to co-infection with infectious myonecrosis virus and Vibrio harveyi in Litopenaeus vannamei. J. Fish. Aquatic Sci., 10: 13-23.
CrossRefDirect Link - O'Sullivan, D.J., 2001. Screening of intestinal microflora for effective probiotic bacteria. Agric. Food Chem., 49: 1751-1760.
CrossRefPubMedDirect Link - Puniya, A.K., A.Z.M. Salem, S. Kumar, S.S. Dagar and G.W. Griffith et al., 2015. Role of live microbial feed supplements with reference to anaerobic fungi in ruminant productivity: A review. J. Integr. Agric., 14: 550-560.
CrossRefDirect Link - Sahu, M.K., N.S. Swarnakumar, K. Sivakumar, T. Thangaradjou and L. Kannan, 2008. Probiotics in aquaculture: Importance and future perspectives. Indian J. Microbiol., 48: 299-308.
CrossRefDirect Link - Salem, A.Z.M., P.H. Robinson, S. Lopez, Y.M. Gohar, R. Rojo and J.L. Tinoco, 2010. Sensitivity of sheep intestinal lactic acid bacteria to secondary compounds extracted from Acacia saligna leaves. Anim. Feed Sci. Technol., 161: 85-93.
CrossRefDirect Link - Verschuere, L., G. Rombaut, P. Sorgeloos and W. Verstraete, 2000. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev., 64: 655-671.
CrossRefPubMedDirect Link - Vine, N.G., W.D. Leukes, H. Kaiser, S. Daya, J. Baxter and T. Hecht, 2004. Competition for attachment of aquaculture candidate probiotic and pathogenic bacteria on fish intestinal mucus. J. Fish Dis., 27: 319-326.
CrossRefDirect Link - Yanbo, W. and X. Zirong, 2006. Effect of probiotics for common carp (Cyprinus carpio) based on growth performance and digestive enzyme activities. Anim. Feed Sci. Technol., 127: 283-292.
CrossRefDirect Link - Wang, Y.B., 2007. Effect of probiotics on growth performance and digestive enzyme activity of the shrimp Penaeus vannamei. Aquaculture, 269: 259-264.
CrossRefDirect Link - Wang, Y.B., J.R. Li and J. Lin, 2008. Probiotics in aquaculture: Challenges and outlook. Aquaculture, 281: 1-4.
CrossRefDirect Link - Widanarni, W., A. Suwanto, S. Sukenda and B.W. Lay, 2003. Potency of vibrio isolates for biocontrol of vibriosis in tiger shrimp (Penaeus monodon) larvae. J. Biotropia, 20: 11-23.
Direct Link - Zokaeifar, H., J.L. Balcazar, C.R. Saad, M.S. Kamarudin, K. Sijam, A. Arshad and N. Nejat, 2012. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol., 33: 683-689.
CrossRefPubMedDirect Link