
Odekanyin Oludele Olayemi
Department of Biochemistry, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Kuku Adenike
Department of Biochemistry, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Akinpelu David Ayinde
Department of Microbiology, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Research Journal of Microbiology
Year: 2015 | Volume: 10 | Issue: 4 | Page No.: 132-144







ABSTRACT
A galactose-specific lectin purified from the skin mucus of African catfish (Clarias gariepinus) was tested for antimicrobial activities against microbial pathogens isolated from the catfish pond water with a view to examining the involvement of the protein in the host defense mechanism. Standard methods of microbial isolation and identification were employed and disc-diffusion method was used for the anti-microbial assay. Anti-microbial evaluation of standard antibiotics ampicillin and nystatin were used as comparative study. The microbial load of the fish pond water was high, with highest total bacterial count of 7.1×108 CFU mL-1 and fungal count of 6.7×103 CFU mL-1. Twelve bacterial species and 14 fungi species were isolated and identified from the pond water. The lectin specifically agglutinated different Gram-negative bacteria, such as Aeromonas hydrophila, Alcaligenes faecalis, Klebsiella edwardsii and Vibrio metschnikovii but had no effect on Gram-positive bacteria. It inhibited strongly the growth of A. hydrophila, A. faecalis, Bacillus cereus, B. polymyxa, K. pneumoniae, K. edwardsii, Pseudomonas aeruginosa and V. metschnikovii. In the same manner, the lectin inhibited the mycelia growth of yeast, Kluyveromyces marxianus and a phytopathogenic fungus, Fusarium oxysporum. The ability of skin mucus lectin to agglutinate and inhibit the growth of bacteria and fungi will make available effective defense mechanism for the African catfish against invading microbes.
PDF Abstract XML References Citation
Received: April 19, 2015;
Accepted: June 01, 2015;
Published: June 30, 2015
How to cite this article
Odekanyin Oludele Olayemi, Kuku Adenike and Akinpelu David Ayinde, 2015. Evaluation of Antimicrobial Potential of a Galactose-Specific Lectin in the Skin Mucus of African Catfish (Clarias gariepinus, Burchell, 1822) against some Aquatic Microorganisms. Research Journal of Microbiology, 10: 132-144.
URL: https://scialert.net/abstract/?doi=jm.2015.132.144
URL: https://scialert.net/abstract/?doi=jm.2015.132.144
INTRODUCTION
Lectins are sugar-binding proteins, with multiple binding sites. They agglutinate cells and precipitate glycoconjugates. They are widely distributed in various organisms from microorganism to higher vertebrates. The unique properties of animal lectins allow them to play important role in biological processes like detection of cell-surface carbohydrates, signal-transduction, metastasis and apoptosis, opsonization, agglutination and proliferation (Tasumi et al., 2002; Argayosa et al., 2011). Lectins have a significant role in the immune responses of the host (Dutta et al., 2005). They bind specifically to the carbohydrate molecules expresses on the pathogens and help in their rapid clearance by enhancing opsonization and phagocytosis and also by increasing the oxidative burst activities (Tateno et al., 2002).
Good number of lectins has been reported from fish and has been characterized in terms of agglutination activities, carbohydrate specificities and involvement in the immune response of the fishes (Dutta et al., 2005; Tsutsui et al., 2007; Argayosa et al., 2011; Odekanyin and Kuku, 2014).
The aquatic environment, where fish live provides suitable conditions for high numbers of both pathogenic and non-pathogenic microorganism. The first defense against pathogens in aquatic organism is the skin mucus ,which is equipped with mechanisms protecting against pathogen entry. Certainly, it is well established that the fish skin secretions contain a variety of biologically active substances, which include immunoglobulins, complements, C-reactive protein, hemolysin, lysozyme, antimicrobial peptides and lectins that are constitutively expressed to provide immediate protection to fish from potential pathogen (Suzuki et al., 2003; Tsutsui et al., 2007). The composition and rate of mucus secretion has been observed to change in response to microbial exposure or to environment perturbations such as hyperosmolarity and acidity (Subramanian et al., 2008). Apart from disease-resistance ability, mucus layer on the fish surface also carried out a number of biological roles including respiration, ionic and osmotic regulation, locomotion, reproduction, communication feeding and nest building (Ingram, 1980; Shephard, 1994).
Some infectious and non-infectious microorganisms have been shown to majorly inhabit the aquatic environment of fish. Reports have shown that there is correlation between the type of microorganism isolated from the aquatic environment of fishes and the one isolated from various organs or parts of fishes (Apun et al., 1999; Nahiduzzaman et al., 2000; Ali and Osman, 2010; Torimiro et al., 2014). The presence of the following bacterial genera Aeromonas, Klebsiella, Micrococcus, Alcaligenes, Vibrio, Flavobacterium, Bacillus, Pseudomonas and Coryneforms have been found to be most common in fish pond water and various organ of fish. Fungi species of Mucor, Aspergillus, Microsporum, Trichophyton and Chrysosporum were more prevalent in pond water (Okaeme and Olufemi, 1997). Some of these microbes have been implicated has the major causative organism of known diseases of fish (Nahiduzzaman et al., 2000; Khatun et al., 2011; Sarkar and Rashid, 2012).
From the literature, there is growing evidence that lectins from the skin mucus of fish have the ability to agglutinate, opsonize and/or suppresses microbial growth (Suzuki et al., 2003; Tasumi et al., 2004; Dutta et al., 2005; Tsutsui et al., 2006b, 2007; Argayosa et al., 2011). Antimicrobial activity of epidermal mucus extracts against a broad range of microbial pathogens was observed by Hellio et al. (2002). This study supported the hypothesis that the epidermal mucus plays a protective function against microbial infection in fish.
We have previously purified and characterized a galactose-specific lectin from the skin mucus of African catfish (Odekanyin and Kuku, 2014). The present study isolated and identified various microorganisms from pond water of catfish farm and checked the functional properties of the African catfish skin mucus galactose-specific lectin against pathogenic and non-pathogenic microbes identified from the pond water. This is with a view to determine the involvement of the protein in the host defense mechanism.
MATERIALS AND METHODS
Collection of water samples: Water samples were collected aseptically at ten different points from three different fish ponds in Osin Farm Ltd, Yakoyo, via Ile-Ife into ten sterile brown bottles. The samples were immediately moved to the laboratory in an ice bag for microbial analysis.
Estimation of total heterophilic bacteria and fungal: Aliquots of 1.0 mL from appropriately diluted water samples were inoculated in duplicates into nutrient agar and Sabouraud dextrose agar using pour plate technique for bacterial and fungal count, respectively. The nutrient agar plates were incubated aerobically at 37°C for 48 h, while the Sabouraud dextrose agar plates were incubated at 30°C for 5-7 days. At the end of the incubation periods, only plates showing between 30 and 300 colonies were enumerated. The average count on duplicate plates was multiplied by dilution factor at that dilution and expressed as colony forming unit per milliliter (CFU mL-1) of the original sample.
Isolation and identification of pure cultures: Each of the distinct colonies from the master plates was carefully subcultured on to fresh plates using a flamed wire-loop for the bacterial colonies and a flamed inoculating needle for the fungi. Nutrient agar plates were used for subculturing bacterial isolates, while Sabourand dextrose agar was used for fungal isolates. The bacterial isolates were incubated at 37°C in a Gallenkamp incubator for 24 h, while fungi plates were incubated at room temperature for four days. Colonies representative of each type of bacterium were stained by Gram’s Method and with Malachite green to detect spore formation and later examined microscopically. As for fungi, the appearance of only one strain of the fungus on the plates indicated a pure culture. All bacterial isolates were identified based on their cultural, morphological and biochemical characteristics tests (Cheesbrough, 2006), which include catalase, Triple Sugar Iron (TSI) medium, Sulphur Indole Motility (SIM), nitrate reduction, fermentation of sugars, citrate utilization, methyl Red/Voges-Proskauer (MR-VP), Oxidation-Fermentation (OF) and Gelatin Liquefaction (GL). The identity of the bacterial isolates were determined with reference to Bergey’s Manual of Determinative Bacteriology (Buchanan and Gibbons, 1974). Fungi isolates were identified according to Barnett and Hunter (1972) and Harrigan and McCance (1976). Actively growing portions of pure fungus colony was aseptically picked and placed on a microscope slide containing a drop of water and later stained with lactophenol solution and examined under the microscope. Identification was based on characteristics, such as colour of hyphae, septation, nature of conidia, etc.
Assay of lectin for antimicrobial activity
Bacterial agglutination test: All the isolated bacteria were tested for agglutination with previously purified lectin (Odekanyin and Kuku, 2014). Both Gram negative and Gram positive bacteria were grown in nutrient broth for about 24 h. The cells were harvested by centrifugation at 3000×g for 2 min and washed with Phosphate Buffered Saline (PBS) three times. The packed cells were suspended in 0.5% formalin solution and shaken at 25°C for 24 h. Formalin-killed cells were collected by centrifugation, washed with PBS and resuspended in PBS to 1.5×108 colony forming unit milliliter (Mcfarland 0.5 standard). Agglutination assays with the formalin-killed cells was performed in the microtitre plates. An equal volume of each bacterial isolate suspension was mixed with a two fold serial dilution of the lectin in a microtitre plate and incubated at room temperature for 1 h. The bacterial agglutination titer was expressed as the reciprocal of the highest dilution giving a visible agglutination upon illumination of the microtitre plates (Dutta et al., 2005; Tasumi et al., 2004).
Antibacterial sensitivity test: The in vitro sensitivity tests of the bacteria to the purified African catfish skin mucus lectin were done by disc diffusion method with little modification (Akinpelu et al., 2009; Adegoke et al., 2010). About 0.2 mL of the standardized 24 h old culture of the test organisms in nutrients broth was inoculated into pre-sterilized molten Mueller-Hinton agar medium at 45°C in MacCartney bottle. The medium was poured into a sterile petri dish and allowed to set. With the aid of a sterile cork borer, three wells of about 6 mm in diameter were bored on the plates equidistant from the centre of the plates. About 0.5 mL of the purified lectin was dispensed into one of the wells and the same volumes of antimicrobial standard drugs-ampicillin and streptomycin at a concentration of 10 and 1 mg mL-1, respectively were dispensed into the remaining wells. The plate was then allowed to stand for about 15 min for prediffusion of the protein to occur. These were then incubated at 37°C for 24 h. At the end of the period, zones of inhibition formed on the agar plates indicates susceptibility of the test isolates to the suspension and were evaluated in mm (Junaid et al., 2006). The mean diameter of the triplicates readings were determined and recorded in mm.
Assay of lectin for antifungal activity: Antifungal activity of the lectin was tested using agar well-diffusion method described by Igbinosa et al. (2009). An antifungal drug (Nystatin) was used as standard. The fungal isolates were allowed to grow on a Sabouraud Dextrose Agar (SDA) (Biotec, UK) at 25°C until they sporulated. The fungal spores were harvested after sporulation by pouring a mixture of sterile glycerol and distilled water to the surface of the plate and spores were later scraped with a sterile glass rod. The harvested fungal spores were standardized to absorbance of 0.1 at 600 nm before use. One hundred microliter of the standardized fungal spore suspension was evenly spread on the SDA (Biotec, UK) using a glass spreader. Wells were then bored into the agar media using a sterile 6 mm cork borer and the wells filled with the solution of the extract taking care not to allow spillage of the solution to the surface of the agar medium. The plates were allowed to stand on the laboratory bench for 1 h to allow for proper diffusion of the extract into the media. Plates were incubated at 25°C for four days and later observed for zones of inhibition. The effect of the lectin on fungal isolates was compared with Nystatin at a 1 mg mL-1.
RESULTS
Microbial load of water samples: The water samples of all the ponds showed various degrees of bacterial and fungal populations (Table 1). The highest total bacterial count of 7.1×108 CFU mL-1 was recorded in pond B, which is used for breeding purposes. The highest fungal count of 6.7×103 CFU mL-1 was recorded in pond A.
Identification of bacteria and fungi in the water samples: The morphological and biochemical characteristics of bacteria isolated from the water samples and their probable identity are shown in Table 2. The total number of bacterial species isolated and identified was 12. Both Gram positive and Gram negative bacteria were isolated.
| Table 1: | Total heterophilic bacterial and fungal counts of C. gariepinus water sample |
 | |
| THB: Total heterophilic bacterial, THF: Total heterophilic fungal | |
| Table 2: | Biochemical characteristics of some bacteria isolated from water samples of C. gariepinus environment |
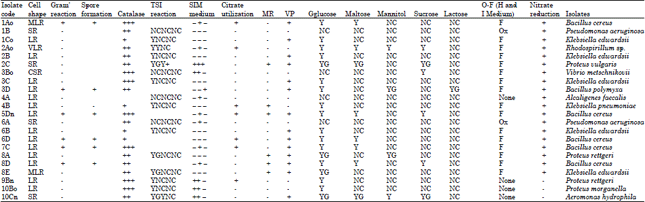 | |
| LR: Long rod, SR: Short rod, MLR: Medium long rod, CSR: Curved short rod, VLR: Very long rod, Y: Acid production only, YG: Acid and gas production, NC: No change, F: Fermentative, Ox: Oxidative, VP: Voges-Proskaurer test, MR: Methyl red test | |
The bacteria isolated were Aeromonas hydrophila, Alcaligenes faecalis, Bacillus cereus, B. polymyxa, Klebsiella pneumoniae, K. edwardsii, Proteus morganella, P. rettgeri, P. vulgaris, Pseudomonas aeruginosa, Rhodospirillum sp. and Vibrio metschnikovii.
The total number of fungi species isolated was 14 and these are Aspergillus niger, A. repens, Botryodiplodia acerina, Fusarium oxysporum, Heliscus aquaticus, Kluyveromyces marxianus, Mucor mucedo, Neosartorya fischeri, Pullularia pullulans, Rhizopus japonicas, Scopulariopsis brevicaulis, Tricelophorus monosporus, Trichoderma sp. and Zygodesmus corda.
Bacterial agglutination study: The fish lectin was able to agglutinate several Gram negative bacteria at varying degree (Table 3). At a concentration equivalent to the titre of 25 that was fully effective in inducing rabbit erythrocyte hemagglutination, fish lectin showed agglutinating activity against four bacteria species namely Aeromonas hydrophila, A. faecalis, Klebsiella edwardsii and Vibrio metschnikovii. The highest titre of 24 was obtained for V. metschnikovii followed by A. hydrophila with 23 and A. faecalis and K. edwardsii have 22.
Antibacterial sensitivity test: Eight of the test bacteria were susceptible to the fish lectin with zones of inhibition ranging between 10.07 and 33.00 mm. The highest activity of the fish lectin was 32.74±0.71 mm diameter of zone of inhibition found against B. cereus followed by 31.34±0.87 and 28.37±0.96 mm against K. edwardsii and V. metschnikovii, respectively. The lowest activity of the fish lectin was against A. faecalis with 10.07±1.14 mm diameter of zone of inhibition (Fig. 1a-g). The fish lectin was inactive against all the species of Proteus and Rhodospirillium. The comparative antibacterial effects of the fish lectin with the standard drugs (Ampicillin and Streptomycin) are shown in Table 4.
| Table 3: | Agglutination of different bacteria on incubation with the African catfish skin mucus lectin |
 | |
| Key: *Denotes degree of agglutination, - denotes no agglutination | |
| Table 4: | Zone of inhibition exhibited by African catfish skin mucus Lectin against the isolated bacteria |
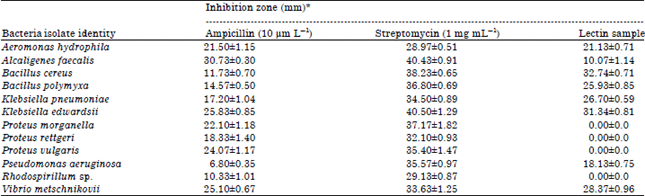 | |
| Values are Mean±SEM of three replicates | |
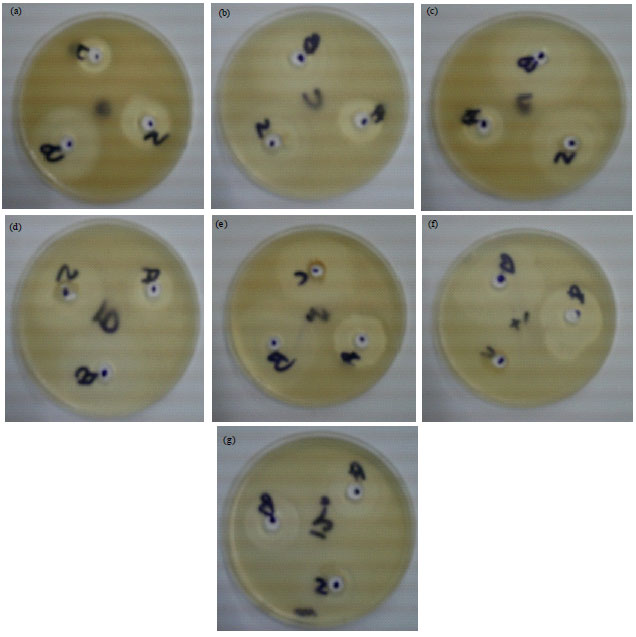 | |
| Fig. 1(a-g): | Inhibition zone around the standard drug (A and B) and sample (lectin), (a) Pseudomonas aeruginosa, (b) Aeromonas hydrophila, (c) Klebsiella pneumonia, (d) Bacillus polymyxa, (e) Vibrio metschnikovii, (f) Klebsiella edwardsii and (g) Bacillus cereus |
Antifungal activities assay: Table 5 showed the antifungal activity of the fish lectin by agar well-diffusion assay. The fish lectin was effective against only two of the fungi tested. The maximum antifungal activity was observed against Kluyveromyces marxianus and minimum was against Fusarium oxysporum. The inhibition zone is comparatively lower than that of standard drug used.
DISCUSSION
In the present study, the bacterial load recorded at various points of the three ponds studied ranges between 1.3×104 and 7.1×108 CFU mL-1. A higher bacterial load was recorded at about the same point in pond A and pond B. This is because the points are very close to the inlet point of the ponds.
| Table 5: | Zone of inhibition exhibited by African catfish skin mucus Lectin against the isolated fungi |
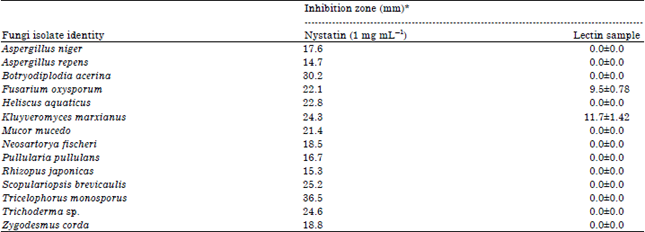 | |
| Values are mean of two replicates | |
The inlet dam possibly have higher bacterial load. It has been noted that water from inlet dam has a slightly higher microbial load than the conventional pond (Apun et al., 1999). The results obtained are in agreement with the reports of Trakroo and Agarwal (2011) and Torimiro et al. (2014). The finding is also in line with the report of Oni et al. (2013) in a study on associated microbial load of artificially cultured C. gariepinus fingerlings, in which high microbial load with low mortality of the fingerlings was recorded. It can therefore be suggested that C. gariepinus is adequately suited to withstand high microbial load.
The present study detected ten different species of Gram-negative and two species of Gram-positive bacteria in the pond water. All belongs to eight genera: Aeromonas, Alcaligenes, Bacillus, Klebsiella, Proteus, Pseudomonas, Rhodospirillum and Vibrio. This is relatively lower than the number of genera obtained by Al-Harbi and Uddin (2003) and Newaj-Fyzul et al. (2008). However, the type and prevalence of organisms differ. Newaj-Fyzul et al. (2008) reported that sixteen genera of bacteria were isolated from pond water with Bacillus species, Staphylococcus sp., Alcaligene sp. and Aeromonas sp. being the most prevalent. No Vibrio sp. was found in the pond water. Trakroo and Agarwal (2011) isolated Proteus sp., Aeromonas hydrophila, Pseudomonas aeruginosa, Flavobacter sp., Vibrio sp. and Corynebacterium sp. from pond water of farm raised Rohu, Labeo rohita. It is evident from this study that bacterial types isolated from catfish environment were all aerobic heterotrophic. It has been reported that potential disease causing bacteria, such as Aeromoonas hydrophila, Pseudomonas aeruginosa and Vibrio sp. (Austin and Austin, 1999) were found in the African catfish pond water, an indication that the fish skin is continually exposed to pathogen attack. Studies conducted by Ogbondeminu (1993) and Ikpi and Offem (2008), (2011) revealed, prevalence of Flexibacter columnaris, Pseudomonas sp., Aeromonas sp., Vibrio sp., Enterobacteriaceae and Gram positive bacteria as common fish pathogens responsible for different bacterial diseases in fish farms in Nigeria.
In the same manner, fungal count revealed high load of fungi in the three ponds studied, with the highest count of 6.7×103 CFU mL-1 found in pond A. The species of fungi isolated were opportunistic and non-pathogenic fungi. Adesulu (2001) suggested that the fungi may have entered the pond water through a number of sources, which include airborne spores, water borne and hyphae, fish feed and fish handling. Singh and Wadhwani (1989) reported that fungi were abundant in stagnant water when compared to flowing water. The high amount of fungal flora can be attributed to the stagnancy of water which provided better chances for germination, growth and survival of fungi on dead and decaying organic matter lying at the pond bottom (Singh and Wadhwani, 1989). Some of the fungi isolated, such as Botryodiploda acerina, Fusarium oxysporum are phytopathogenic fungi. A number of fungi are used in bioremediation in which the fungi are mixed with polluted water or soil, where they decompose the organic material in pollutants and in the detoxify them in the process. Fungi employed in this way include many that are commonly found in soils, such as Aspergillus, Fusarium, Rhizopus, Mucor, Penicillium and Trichoderma (Ammirati and Seidl, 2008).
Most animal lectins function internally, that is in the tissues, blood plasma or serum but the skin mucus lectin function externally (Muramoto et al., 1999; Tasumi et al., 2002, 2004; Tsutsui et al., 2006a, b). The biological roles of internal lectins include activation of the classical complement pathway by mannose-binding proteins. So, they play important roles in conferring innate immunity. It has also been reported that lectins functioned in pathogen recognition and clearance as part of the innate immune response because of their carbohydrate-binding specificity. Lectins with affinity for galactose have been shown to have vital role in modulating immune response in fish (Yousif et al., 1994; Mistry et al., 2001).
In the current study, African catfish skin mucus (a galactose-specific lectin) lectin demonstrated agglutination activity against two pathogenic and two non-pathogenic but opportunistic species of bacteria. Interestingly, when different species of Gram-positive and Gram-negative bacteria were incubated with African catfish skin mucus lectin, only Gram-negative bacteria were agglutinated but growth of Gram-positive bacteria such as Bacillus cereus and B. polymxa as well as Gram-negative bacteria like Aeromonas hydrophila, Vibrio metschnikovii and Pseudomonas aeruginosa were significantly suppressed (Fig. 1a-g). Also, the growth of Klebsiella sp. and Alcaligene faecalis were inhibited. On the other hand, the lectin was not effective against Proteus sp. and Rhodospirillum sp. tested. Proteus sp. and Rhodospirillum sp. have not been reported to be a threat to survival of fishes in their environment, though they have been shown to be prominent among bacterial flora of aquatic environment (Al-Harbi and Uddin, 2003; Newaj-Fyzul et al., 2008). It is clearly shown here that African catfish skin mucus lectin possess agglutination activity against some environmental bacteria, suggesting that the lectin may provide a self-defense mechanism on the skin surface of the fish. This might be responsible for the catfish ability to tolerate extreme environmental conditions and also for the increase in mucus secretion noticed, when the fish was moved into new water environment.
To date, a number of fish skin mucus lectins have been reported to possess agglutination activity against bacteria but the activity was specific for certain bacterial species. Congerin in Conger eel was shown to agglutinate but not inhibit the growth of Vibrio anguillarum (Kamiya et al., 1988). In Japanese eel, AJL-2 agglutinated Escherichia coli and suppressed its growth (Tasumi et al., 2002) and AJL-1 was the first lectin demonstrated to agglutinate a Gram-positive bacterium, Streptococcus difficile (Tasumi et al., 2004). Pufflectin-s in torafugu (Takifugu rubripes) showed agglutination activity towards some bacterial isolates, mainly of the genus Vibrio but was unable to agglutinate five pathogenic bacteria tested (Tsutsui et al., 2006a). The intelectin catfish, Silurus asotus showed agglutination activity against the pathogenic bacterium Aeromonas salmonicida (Tsutsui et al., 2011).
Dutta et al. (2005) reported agglutination of Gram-negative bacteria, when Gram-positive and Gram-negative were incubated with catfish serum lectin. It was noted that incubation of the lectin at nanogram level brought about significant growth inhibition of various Aeromonas strains. Fucose-binding lectin in bighead carp gill specifically agglutinated Vibrio harveyi, a fish pathogen and also inhibited its growth. Conversely, the lectin showed no agglutination activity towards the fungi and Gram-positive bacteria tested (Pan et al., 2010). Argayosa et al. (2011) observed positive microbial cell agglutination activities of the African catfish serum lectin against Candida albican, Saccharomyces cerevisiae, Aeromonas hydrophila, Bacillus subtilis, Enterococcus faecalis, E. coli and Pseudomonas aeruginosa. However, it is notable that skin mucus lectins in fishes cannot agglutinate all bacteria but exhibit activities specific for certain bacteria species, which means most bacteria might have mechanisms for evading the lectin associated self-defense system of fish. It has been hypothesized that the agglutination of pathogenic organisms may inhibit their penetration into the body or allows them to be trapped in the skin mucus to be washed away with further secretion of mucus. In addition, lectin may inhibit the growth of such organisms in the mucus, some of which may serve as bacterial nutrients. It is also possible for skin mucus lectin to act together with other humoral factors that confer innate immunity such as immunoglobulin and complement.
Antifungal activity of African catfish skin mucus lectin was determined against all the isolated fungi but mycelia growth of Kluyveromyces marxianus and Fusarium oxysporum were only inhibited by the lectins. Until now, the antifungal activity of fish skin mucus lectin has been determined in only few species of fish. Kamiya and Shimizu (1980) reported the first skin mucus lectin that agglutinates fungi. Windowpanr flounder (Lophopsetta maculata) skin mucus lectin specifically agglutinated Saccharomyces cerevisiae. Recently, a mannose-specific C-type lectin with agglutination against yeast, Saccharomyces cerevisiae was purified from Conger eel skin mucus by Tsutsui et al. (2007). Apart from skin mucus, serum lectin isolated from African catfish has also been found to agglutinate two species of fungi: Candida albicans and Saccharomyces cerevisiae (Argayosa et al., 2011).
CONCLUSION
In summary, the African catfish skin mucus galactose-specific lectin demonstrates its assumed biological role in protection against pathogens by its ability to identify, agglutinate and inhibit the growth of pathogenic microbe with galactosides-containing glycans, isolated from catfish pond water. The unique ability prevents the pathogens from attaching themselves to the skin of fish or allowing such pathogens to be washed away together with old mucus into surrounding water.
REFERENCES
- Adegoke, A.A., P.A. Iberi, D.A. Akinpelu, O.A. Aiyegoro and C.I. Mboto, 2010. Studies on phytochemical screening and antimicrobial potentials of Phyllanthus amarus against multiple antibiotic resistant bacteria. Int. J. Applied Res. in Nat. Prod., 3: 6-12.
Direct Link - Akinpelu, D.A., O.A. Aiyegoro and A.I. Okoh, 2009. Studies on the biocidal and cell membrane disruption potentials of stem bark extracts of Afzelia Africana (Smith). Biol. Res., 42: 339-349.
CrossRefDirect Link - Al-Harbi, A.H. and N. Uddin, 2003. Quantitative and qualitative studies on bacterial flora of hybrid tilapia (Oreochromis niloticus x O. aureus) cultured in earthen ponds in Saudi Arabia. J. Aquacult. Res., 33: 43-48.
CrossRefDirect Link - Ali, M.S. and G.A. Osman, 2010. Microbial load as pollution indicator in water of El-Khadra lake at Wadi El-Natroun, Egypt. J. Am. Sci., 6: 489-496.
Direct Link - Apun, K., A.M. Yusof and K. Jugang, 1999. Distribution of bacteria in tropical freshwater fish and ponds. Int. J. Environ. Health Res., 9: 285-292.
CrossRefDirect Link - Argayosa, A.M., R.A.D. Bernal, A.U. luczon and J.S. Arboleda, 2011. Characterization of mannose-binding protein isolated from the African catfish (Clarias gariepinus) serum. Aquaculture, 310: 274-280.
CrossRefDirect Link - Dutta, S., B. Sinha, B. Bhattacharya, B. Chatterjee and S. Mazumder, 2005. Characterization of a galactose binding serum lectin from the Indian catfish, Clarias batrachus: possible involvement of fish lectins in differential recognition of pathogens. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 141: 76-84.
CrossRefDirect Link - Hellio, C., A.M. Pons, C. Beaupoil, N. Bourgougnon and Y. Le Gal, 2002. Antibacterial, antifungal and cytotoxic activities of extracts from fish epidermis and epidermal mucus. Int. J. Antimicrob. Agents, 20: 214-219.
CrossRefDirect Link - Igbinosa, O.O., E.O. Igbinosa and O.A. Aiyegoro, 2009. Antimicrobial activity and phytochemical screening of stem bark extracts from Jatropha curcas (Linn). Afr. J. Pharm. Pharmacol., 3: 58-62.
Direct Link - Ikpi, G. and B. Offem, 2011. Bacterial infection of mudfish Clarias gariepinus (Siluriformes: Clariidae) fingerlings in tropical nursery ponds. Revised Biol. Tropics, 59: 751-759.
Direct Link - Ingram, G.A., 1980. Substances involved in the natural resistance of fish to infection: A review. J. Fish Biol., 16: 23-60.
CrossRefDirect Link - Junaid, S.A., A.O. Olabode, F.C. Onwuliri, A.E.J. Okwori and S.E. Agina, 2006. The antimicrobial properties of Ocimum gratissimum extracts on some selected bacterial gastrointestinal isolates. Afr. J. Biotechnol., 5: 2315-2321.
Direct Link - Kamiya, H., K. Muramoto and R. Goto, 1988. Purification and properties of agglutinins from conger eel Conger myriaster. Dev. Comp. Immunol., 12: 309-318.
CrossRefDirect Link - Kamiya, H. and Y. Shimizu, 1980. Marine biopolymers with cell specificity: II. Purification and characterization of agglutinins from mucus of windowpane flounder Lophopsetta maculata. Biochimica et Biophysica Acta (BBA)-Protein Structure, 622: 171-178.
CrossRefDirect Link - Khatun H., M.D. Hossain. S.N. Jahan and D.A. Khanom, 2011. Bacterial infestation in different fish at Rajshahi. J. Sci. Foundation, 9: 77-84.
CrossRefDirect Link - Mistry, A.C., S. Honda and S. Hirose, 2001. Structure, properties and enhanced expression of galactose-binding C-type lectins in mucous cells of gills from freshwater Japanese eels (Anguilla japonica). Biochem. J., 360: 107-115.
Direct Link - Muramoto, K., D. Kagawa, T. Sato, T. Ogawa, Y. Nishida and H. Kamiya, 1999. Functional and structural characterization of multiple galectins from the skin mucus of conger eel, Conger myriaster. Comp. Biochem. Physiol., 123B: 33-45.
CrossRefDirect Link - Nahiduzzaman, M., M.A. Ehshan, B.R. Chowdhury and M.A.R. Mridha, 2000. Studies on bacterial flora in a farmed catfish, Clarias hybrid. Pak. J. Biol. Sci., 3: 429-432.
CrossRefDirect Link - Newaj-Fyzul, A., A. Mutani, A. Ramsubhag and A. Adesiyun, 2008. Prevalence of bacterial pathogens and their anti-microbial resistance in tilapia and their pond water in trinidad. Zoonoses Public Health, 55: 206-213.
CrossRefDirect Link - Odekanyin, O.O. and A. Kuku, 2014. Charaterization of galactose-specific lectin from the skin mucus of African catfish Clarias gariepinus (Burchell, 1822). Sci. Res. Essay, 9: 869-879.
CrossRefDirect Link - Pan, S., J. Tang and X. Gu, 2010. Isolation and characterization of a novel fucose-binding lectin from the gill of bighead Carp (Aristrichthys nobilis). Veter. Immunol. Immunopathol., 133: 154-164.
CrossRefDirect Link - Sarkar, M.J.A. and M.M. Rashid, 2012. Pathogenicity of the bacterial isolate Aeromonas hydrophila to catfishes, carps and perch. J. Bangladesh Agric. Univ., 10: 157-161.
CrossRefDirect Link - Shephard, K.L., 1994. Functions for fish mucus. Rev. Fish Biol. Fish., 4: 401-429.
CrossRefDirect Link - Subramanian, S., N.W. Ross and S.L. MacKinnon, 2008. Comparison of the biochemical composition of normal epidermal mucus and extruded slime of hagfish (Myxine glutinosa L.). Fish Shellfish Immunol., 25: 625-632.
CrossRefDirect Link - Suzuki, Y., S. Tasumi, S. Tsutsui, M. Okamoto and H. Suetake, 2003. Molecular diversity of skin mucus lectins in fish. Biochem. Mol. Biol., 136: 136-730.
CrossRefDirect Link - Tasumi, S., T. Ohira, I. Kawazoe, H. Suetake, Y. Suzuki and K. Aida, 2002. Primary structure and characteristics of a lectin from skin of mucus of Japanese eel Anguilla japonica. J. Biol. Chem., 277: 27305-27311.
CrossRefDirect Link - Tasumi, S., W.J. Yang, T. Usami, S. Tsutsui, T. Ohira, I. Kawazoe, M.N. Wilder, K. Aida and Y. Suzuki, 2004. Characteristics and primary structure of galectin in the skin mucus of Japanese eel, Anguilla japonica. Comp. Biochem. Physiol., 28C: 325-335.
CrossRefDirect Link - Tateno, H., T. Ogawa, K. Muuramoto, S. Kamiya and M. Saneyoshi, 2002. Rhamnose- binding lectins from steelhead trout (Oncorhynchus mykiss) eggs recognize bacterial lipopolysaccharides and lipoteichoic acid. Biosci. Biotechnol. Biochem., 66: 604-612.
CrossRefDirect Link - Tsutsui, S., K. Iwamoto, O. Nakamura and T. Watanabe, 2007. Yeast-binding C-type lectin with opsonic activity from conger eel (Conger myriaster) skin mucus. Molec. Immunol., 44: 691-702.
CrossRefDirect Link - Tsutsui, S., H. Nishikawa, N. Mano, H. Hirose, S. Tasumi, H. Suetake and Y. Suzuki, 2006. Possible role of a skin mucus lectin from fugu Takifugu rubripes in excluding marine bacteria from the body surface. Fish. Sci., 72: 455-457.
CrossRefDirect Link - Tsutsui, S., M. Okamoto, S. Tasumi, H. Suetake, K. Kikuchi and Y. Suzuki, 2006. Novel mannose-specific lectins found in torafugu Takifugu rubripes: A review. Comp. Biochem. Physiol., 1D: 122-127.
CrossRefDirect Link