
Magdy M. Afifi
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Assuit, 71524, Egypt
Houssam M. Atta
Department of Botany and Microbiology, Faculty of Science (Boys), Al-Azhar University, Cairo, Egypt
Reda A. Bayoumi
Department of Biotechnology, Faculty of Science and Education, Al-Khurmah Branch, Taif University, KSA
Research Journal of Microbiology
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 1-15







ABSTRACT
This research aims to isolate and identify some actinomycetes, that having antimicrobial substances against some pathogenic bacteria and fungi, from soil samples in Al-Khurmah governorate. Only one actinomycete culture KSA-20 exhibited to produce wide spectrum of antibacterial activities. The nucleotide sequence of the 16S rRNA gene of this strain evidenced an 95% similarity with Streptomyces griseoviridis. From the taxonomic features, the actinomycete isolate KSA-20 matches with Streptomyces griseoviridis in the morphological, physiological and biochemical characters. Thus, it given the suggested name Streptomyces griseoviridis KSA-20. The parameters controlling the biosynthetic process of antibacterial agent were fully investigated. The separation of the active ingredient and its purification were performed using both Thin Layer Chromatography (TLC) and Column Chromatography (CC) techniques. The minimum inhibition concentrations “MICs” of the purified antibiotic were also determined. The purified antibiotic suggestive of being belonging to griseoviridin antibiotic produced by Streptomyces griseoviridis KSA-20.
PDF Abstract XML References Citation
Received: December 12, 2013;
Accepted: January 04, 2014;
Published: March 01, 2014
How to cite this article
Magdy M. Afifi, Houssam M. Atta and Reda A. Bayoumi, 2014. Microbiological Studies on the Production of Antimicrobial Agent by Actinomycete Isolated from Saudi Arabia. Research Journal of Microbiology, 9: 1-15.
URL: https://scialert.net/abstract/?doi=jm.2014.1.15
URL: https://scialert.net/abstract/?doi=jm.2014.1.15
INTRODUCTION
Actinomycetes play an important ecological role in recycling substances in the nature and able to inhibit both Gram-positive and Gram-negative pathogen (Khucharoenphaisan et al., 2012). In addition, Streptomyces griseoviridis is known to produce antimicrobial metabolites and be value in a biological control program (Cuppels et al., 2013). Griseoviridin/viridogrisein from Streptomyces griseoviridis follow streptogramin family (Ahmed and Donaldson, 2007; Barriere et al., 1998). Several pharmaceutical companies described various similar compounds in streptogramin antibiotic but did not develop them further (Greenwood, 2008). Their regulation is highly complex and influenced by general physiological conditions (Zhang et al., 2012). Many investigations have neen carried out for synthesizes of antibiotics which possess biological activities from Streptomyces from soil samples (Afifi et al., 2012a, b; Atta et al., 2011).
In the present study, the production of the bioactive substances that exhibited inhibitory effects on some pathogenic bacteria and fungi from Streptomyces griseoviridis were reported, along with some physico-chemical properties of secondary metabolites with high biological effects.
MATERIALS AND METHODS
Actinomycete strain: Strain KSA-20 was isolated from a suspension of a soil sample (Williams and Davies, 1965) collected from Al-Khurmah governorate, Saudi Arabia and inoculated onto a Starch-nitrate agar medium (SNA). Plates were incubated at 35°C for five days. The isolates were individually maintained on Starch-nitrate agar at 4°C and stored as a mixture of hyphae and spores in 20% glycerol at 80°C. The selected isolate was allowed to grow on (SNB) in a purpose to get a clear supernatant for antimicrobial activity.
Test organisms
Bacteria
Gram-positive bacteria: Staphylococcus aureus, NCTC 7447; Bacillus subtilis, NCTC 1040; Bacillus pumilus, NCTC 8214 and Micrococcus luteus, ATCC 9341.
Gram-negative bacteria: Escherichia coli, NCTC 10416; Klebsiella pneumonia, NCIMB 9111 and Pseudomonas aeruginosa, ATCC 10145.
Fungi
Unicellular fungi: Candida albicans, IMRU 3669 and Saccharomyces cerevisiae, ATCC 9763.
Filamentous fungi: Aspergillus niger, IMI 31276; A. flavus, IMI 111023, A. fumigatous, ATCC 16424; Fusarium oxysporum and Penicillium chrysogenium.
Screening for antimicrobial activity: The antimicrobial activity was determined by cup method (Kavanagh, 1972).
Taxonomic studies of actinomycete isolate: Morphological characteristics of the most potent produces strain KSA-20 grown on SNA medium at 35°C for 5 days was examined under scanning electron microscopy (JEOL Technics Ltd.) (Fig. 1).
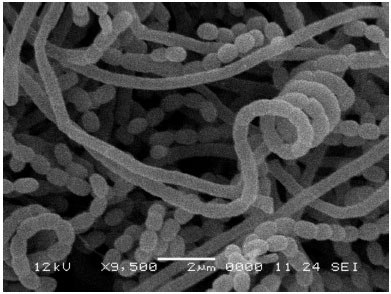 | |
| Fig. 1: | Scanning electron micrograph of the actinomycete isolate KSA 20 growing on SNA showing spiral shape spore chain and smooth spore surface (X 9.500) |
Physiological and biochemical characteristics: The ability of the strain to produce different enzymes was examined by using standard methods. Lecithinase was conducted on egg-yolk medium (Nitsch and Kutzner, 1969), Lipase (Elwan et al., 1977), Protease (Chapman, 1952), Pectinase (Hankin et al., 1971); α-amylase (Cowan, 1974) and Catalase (Jones, 1949). Melanin pigment (Pridham et al., 1957). Degradation of Esculin and Xanthine (Gordon et al., 1974). Nitrate reduction (Gordon, 1966). Hydrogen sulphide production and oxidase test (Cowan, 1974). The utilization of different carbon and nitrogen sources (Pridham and Gottlieb, 1948). Cell wall was performed (Becker et al., 1964; Lechevalier and Lechevalier, 1970). Cultural characteristics such as color of aerial mycelium, color of substrate mycelium and pigmentation of the selected actinomycete were recorded (Shirling and Gottlieb, 1966). Color characteristics were assessed (Kenneth and Deane, 1955).
DNA isolation and manipulation: The locally isolated actinomycete strain was grown for 5 days on a starch agar slant at 35°C. Two milliliter of a spore suspension were inoculated into the starch-nitrate broth and incubated for 3 days on a shaker incubator at 200 rpm and 30°C to form a pellet of vegetative cells (pre-sporulation). The preparation of total genomic DNA was conducted (Sambrook et al., 1989).
Amplification and sequencing of the 16S rRNA gene: PCR amplification of the 16S rRNA gene of the local actinomycete strain was conducted using two primers, StrepF; 5-ACGTGTGCAGCCCAAGACA-3 and StrepR; 5-ACAAGCCCTGGAAACGGGGT-3 (Edwards et al., 1989). The PCR mixture consisted of 30 pmol of each primer, 100 ng of chromosomal DNA, 200 μM dNTPs and 2.5 units of Taq polymerase, in 50 μL of polymerase buffer. Amplification was conducted for 30 cycles of 1 min at 94°C, 1 min of annealing at 53°C and 2 min of extension at 72°C. The PCR reaction mixture was then analyzed via agarose gel electrophoresis and the remaining mixture was purified using QIA quick PCR purification reagents (Qiagen, USA). The 16S rRNA gene was sequenced on both strands via the dideoxy chain termination method (Sanger et al., 1977). The 16S rRNA gene (1.5 kb) sequence of the PCR product was acquired using a Terminator Cycle Sequencing kit (ABI Prism 310 Genetic Analyzer, Applied Biosystems, USA).
Sequence similarities and phylogenetic analysis: The BLAST program (www.ncbi.nlm.nih.gov/blast) was employed in order to assess the degree of DNA similarity. Multiple sequence alignment and molecular phylogeny were evaluating using BioEdit software (Hall, 1999). The phylogenetic tree was displayed using the TREE VIEW program.
Fermentation: The Streptomyces griseoviridis, KSA-20 inoculum was introduced aseptically into each sterile flask containing the following ingredients (g L-1): glucose, 20; KNO3, 2.0; K2HPO4, 0.8; MgSO4.7H2O, 0.7 and KCl, 0.5. The pH was adjusted at 7.0 before sterilization. After 5 days of incubation at 35°C. Filtration was carried out through cotton wool and followed by centrifugation at 5000 rpm for 15 min.
Extraction: The culture filtrates was extracted twice with n-Butanol and the pooled solvent extracts were evaporated to dryness under vacuum to yield a crude residue.
Precipitation: The precipitation process of the crude compound was carried out using petroleum ether (bp 60-80°C) followed by centrifugation at 5000 rpm for 15 min.
Purification by TLC: Separation of the antimicrobial compound into its individual components was conducted by thin layer chromatography using chloroform and methanol (24: 1, v/v) as a solvent system.
Purification by column chromatography: The purification of the antimicrobial compound was carried out using silica gel column (2.5x50) chromatography. Chloroform and methanol 10:2 (v/v), was used as an eluting solvent. The column was left overnight until the silica gel (Prolabo) was completely settled. One milliliter crude extract to be fractionated was added on the silica gel column surface and the extract was adsorbed on top of silica gel. Fifty fractions were collected (each of 5 mL) and tested for their antimicrobial activities.
Physicochemical properties
Elemental analysis: The elemental analysis including C, H, O, N and S was carried out at the microanalytical center, Cairo University, Egypt.
Spectroscopic analysis: The IR, UV and Mass spectrum were determined at the micro analytical center of Cairo University, Egypt.
Biological activity: The Minimum Inhibitory Concentration (MIC) has been determined by the cup method (Kavanagh, 1972).
Characterization of the antimicrobial agent: The antibiotic produced by Streptomyces griseoviridis, KSA-20 was identified according to the recommended international references of Berdy (1974) and Umezawa (1977).
RESULTS
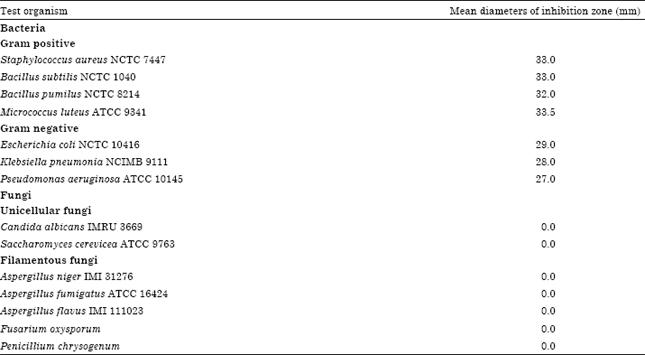
Screening for the antimicrobial activities: The active metabolites produced by Streptomyces griseoviridis, KSA-20 exhibited various degrees of activities against Gram positive and Gram negative bacteria as reported in Table 1.
Identification of the actinomycete isolate
Morphological characteristics: Spore chain is spiral, spore mass is red; spore surface is smooth and the reverse is light grayish yellow; diffusible pigment production is grayish red orange.
Cell wall hydrolysate: The cell wall hydrolysate contains LL-diaminopimelic acid (LL-DAP) while sugar pattern not detected.
Physiological and biochemical characteristics: The actinomycete isolate KSA-20 could hydrolyze protein, starch, cellulose, lipid, lecithin and pectin, while catalase and melanin pigment are negative. In addition, degradation of xanthine, esculine production of H2S, decomposition of urea, nitrate reduction and utilization of citrate are positive but coagulation of milk are negative.
| Table 1: | Mean diameters of inhibition zones (mm) caused by 100 μL of the antimicrobial activities produced by Streptomyces griseoviridis, KSA-20 in the agar plate diffusion assay (The diameter of the used cup assay was 10 mm) |
 | |
The isolate under study utilizes raffinose, mannitol, L-arabinose, meso-inositol, lactose, maltose, trehalose, D-meliztose, D-mannose, D-glucose and rhamnose but do not utilize D- galactose, D- fructose and sucrose, whereas, D-xylose and sodium citrate are doubtful. Good growth on L-cycteine, L-histidine and L-arginine. No growth on L-valine, L-phenyl alanine, L-lysine, L-Hydroxyproline and L-glutamic acid. Moreover, good growth in the presence of 4% NaCl. The growth is not inhibited in the presence of sodium azide and phenol. The actinomycete isolate showed non-resistant to Pencillin G, Amoxicillin, Gentamycin and Cephalosporin “Keflex”. Moreover, it resistant to Augmentin, Carpenicillin, Cefadroxil, Cephardine Ceptrine, Chloramphinicol, Cloxacillin, Doxycycline, Neomycin, Erythromycin, Rifampicin and Noroxin. The isolate KSA-20 is active against Gram positive and Gram negative bacteria (S. aureus, NCTC 7447; B. subtilis, NCTC 1040; M. luteus, NCTC 1089; E. coli, NCTC 10416 and K. pneumonia NCIMB 9111) (Table 2).
Color and culture characteristics: The isolate KSA-20 exhibited good growth on SNA, the aerial mycelium is medium red, substrate mycelium is light yellowish brown and the diffusible pigment grayish red orange. No growth on tryptone-yeast extract broth medium (ISP-1), yeast extract-malt extract agar medium (ISP-2) and glycerol-asparagine agar medium (ISP-4). Moderate growth was detected on Oat-meal agar medium (ISP-3), aerial mycelium is medium is red, substrate mycelium light garish orange and failed to produce diffusible pigment. Moderate growth was detected on inorganic salts-starch agar medium (ISP-4), aerial mycelium reddish gray, substrate mycelium light yellowish brown and diffusible pigment grayish red orange.
| Table 2: | Morphological, physiological and biochemical characteristics of the actinomycete isolate |
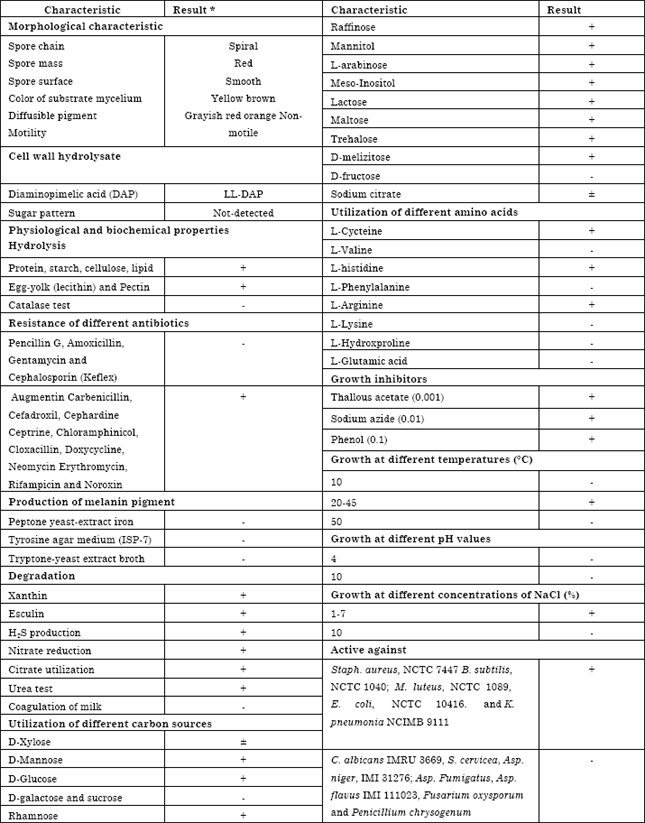 | |
| *+: Positive, -: Negative and ±: Doubtful results | |
| Table 3: | Cultural characteristics of the actinomycete isolate |
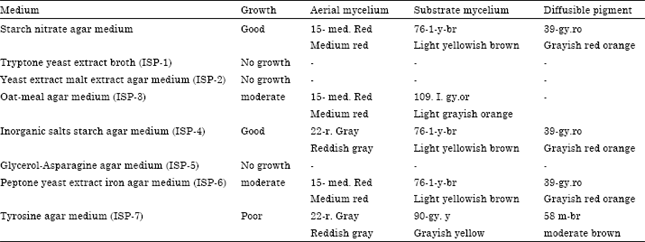 | |
| *The color of the organism under investigation was consulted with the ISCC-NBS color-name charts illustrated with centroid color | |
Moderate growth was detected on peptone yeast extract-iron agar medium (ISP-6), aerial mycelium medium red, substrate mycelium light yellowish brown and diffusible pigment grayish red orange. Poor growth was detected on tyrosine agar medium (ISP-7), aerial mycelium reddish gray, substrate mycelium grayish yellow and diffusible pigment moderate brown (Table 3).
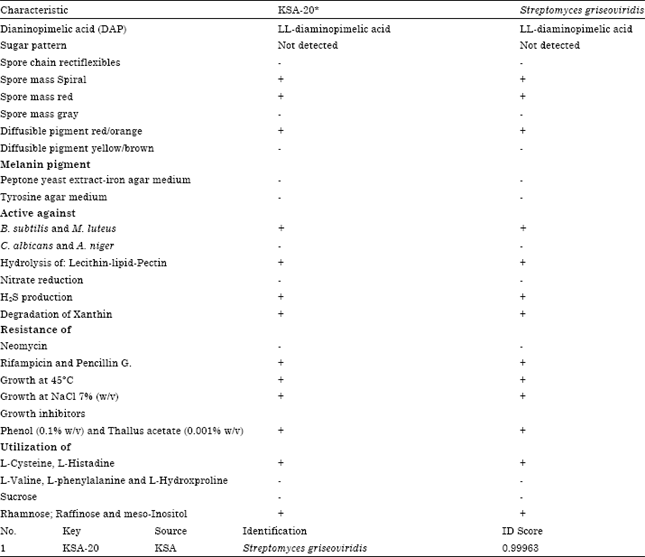
Taxonomy of actinomycete isolate, KSA-20: This was performed basically according to the recommended international Key’s (Buchanan and Gibbons, 1974; Hensyl, 1994; NTP, 1989). On the basis of the previously collected data and in view of the comparative study of the recorded properties of KSA-20 in relation to the most closest reference strain, viz., Streptomyces griseoviridis, it could be stated that actinomycetes isolate, KSA-20 is suggestive of being likely belonging to Streptomyces griseoviridis, KSA-20 (Table 4).
Amplification of the 16S rRNA gene: The 16S rRNA gene was amplified by Polymerase Chain Reaction (PCR) using the universal primers. The primers that was used to 16S rRNA sequencing were of the sequence strepF; 5'-ACGTGTGCAGCCCAAGACA-3' and strepR; 5'-ACAAGCCCTGGAAACGGGGT-3', the product of the PCR was analyzed on 1.5% ethidium bromide gel.
Molecular phylogeny of the selected isolate: The 16S rDNA sequence of the local isolate was compared to the sequences of Streptomyces spp. In order to determine the relatedness of the local isolate to Streptomyces strains, the phylogenetic tree (as displayed by the Tree View program) revealed that the locally isolated strain is closely related to Streptomyces sp., rather related to Streptomyces sp. rather than to Streptomyces griseoviridis (Fig. 2). Multiple sequence alignment was conducted the sequences of the 16S rRNA gene of Streptomyces griseoviridis. Computer assisted DNA searches against bacterial database similarly revealed that the 16S rRNA sequence was 95% identical Streptomyces griseoviridis (Fig. 2).
Fermentation, extraction and purification: The fermentation process was carried out for five days at 35°C. After incubation period, the filtration was conducted followed by centrifugation at 4000 rpm for 15 min. The entire culture broth (20 L) was centrifuged (4000 rpm, 15 min) to separate the mycelium and the supernatant.
| Table 4: | Numerical taxonomy of Streptomyces species program (PIB WIN) |
 | |
| *+: Positive, -: Negative, ±: Doubtful results | |
The supernatant was extracted with n-butanol (1:1, v/v) and the organic layer was evaporated to give an oily material. The oily material was then dissolved in 15% aqueous methanol and defatted by partitioning with petroleum ether (bp 60-80°C) to give a solid extract. Its color is yellowish. Separation of antimicrobial agent into individual components was carried out by TLC using a solvent system composed of chloroform and methanol (24: 1, v/v). Only one band at Rf = 0.7 showed antimicrobial activity. The purification process through column chromatography packed with silica gel, revealed that the most active fractions against the tested organisms ranged between 20 to 30.
Physicochemical characteristics: The purified antibacterial agent produced by Streptomyces griseoviridis, KSA-20 has a characteristic odour and a melting point of 155°C. The compound is freely soluble in chloroform, ethyl acetate, n-butanol, acetone, ethyl alcohol, methanol and 10% isopropyl alcohol but insoluble in petroleum ether, hexan and benzene.
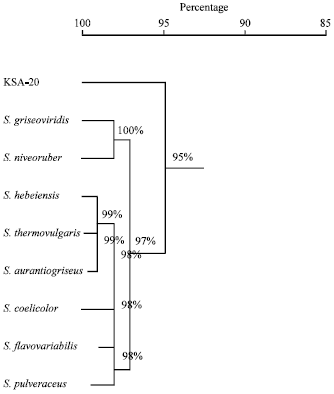 | |
| Fig. 2: | The phylogenetic position of the local Streptomyces sp. strain among neighboring species. The phylogenetic tree was based on the pairwise comparisons of 16S rRNA sequences |
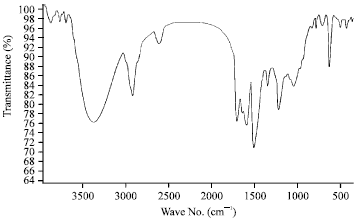 | |
| Fig. 3: | IR spectrum of antibacterial agent produced by Streptomyces griseoviridis, KSA-20 |
Elemental analysis: The elemental analytical data of the antibacterial agent produced by Streptomyces griseoviridis, KSA-20, given the following percent composition: C = 55.33%, H = 5.70%, N = 8.80%, O = 23.45% and S = 6.71%. This analysis indicates a suggested empirical formula of C22H27N3O7S.
Spectroscopic characteristics: The spectroscopic analysis of the purified of antibacterial compound produced by Streptomyces griseoviridis, KSA-20; showed the following data: The infrared (IR) sepectrum showed characteristic band corresponding to 26 peaks (Fig. 3), the ultraviolet (UV) absorption spectrum recorded a maximum absorption peak at 221 nm (Fig. 4) and the mass spectrum showed molecular weight at 477.4 (Fig. 5).
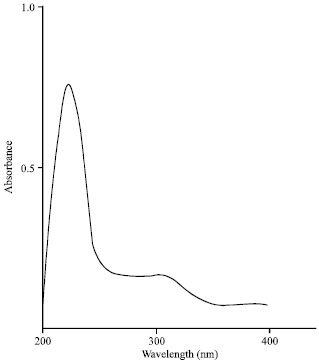 | |
| Fig. 4: | Ultraviolet absorbance of antibacterial agent produced by Streptomyces griseoviridis, KSA-20 |
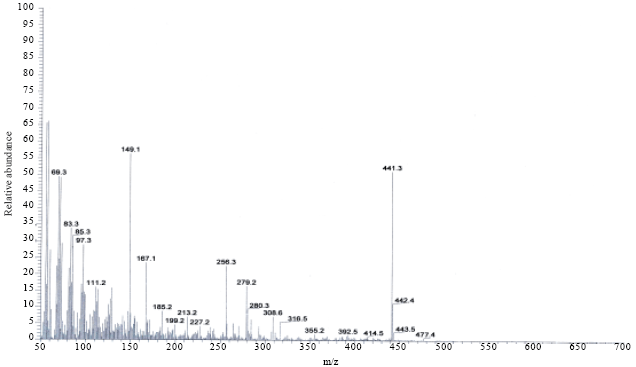 | |
| Fig. 5: | Mass spectrum of antibacterial agent produced by Streptomyces griseoviridis, KSA-20 |
Biological activities of the antimicrobial agent: Data recorded in Table 5 indicated that the antimicrobial agent is fairly active against both Gram positive and Gram negative bacteria.
| Table 5: | Biological activities (MIC) of the antibacterial agent by cup method assay |
 | |
Identification of the antimicrobial agent: On the basis of the recommended keys for the identification of antibiotics and in view of the comparative study of the recorded properties of the antimicrobial agent, it could be stated that the antibacterial compound is suggestive of being belonging to griseoviridin antibiotic (Berdy, 1974; Umezawa, 1977).
DISCUSSION
Actinomycetes had been recognized as the potential producers of metabolites such as antibiotics, growth promoting substances for plants and animals, immunmodifiers, enzyme inhibitors and many other compounds of use to man. The actinomycetes have the ability to produce secondary metabolites with biological activities such as antibiotic, antifungal compounds (Afifi et al., 2012a, b; Atta et al., 2011). The Streptomyces griseoviridis, KSA-20 was isolated from Al-Khurmah governorate and produce a wide spectrum antimicrobial agent against Gram positive and Gram negative bacteria and unicellular and filamentous Fungi. Streptomyces griseoviridis, KSA-20 isolate was growing on SNA to investigate its potency to produce antimicrobial agents.
The Streptomyces species under investigation exhibited a wide spectrum antibacterial agent as previously reported (Dhananjeyan et al., 2010; Kavanagh, 1972; Zamanian et al., 2005). Due to the selective isolation of soil actinomycetes for finding novel strains which can produce useful bioactive compounds, thus various culture media and techniques have been deveoped (Dhananjeyan et al., 2010; Hozzein et al., 2008). Identification process had been performed (Hensyl, 1994; NTP, 1989; Williams, 1989). The morphological characteristics and microscopic examination emphasized that the spore chain is spiral. Spore mass is red, while spore surface is smooth, substrate mycelium is light yellow-brown and diffusible pigment grayish red orange. The results of physiological, biochemical characteristics (Table 2) and cell wall hydrolysate of actinomycetes isolate, exhibited that the cell wall containing LL-diaminopimelic acid (DAP). These results emphasized that the actinomycetes isolate related to a group of Streptomyces as previously studied (Afifi et al., 2012a, b; Muharram et al., 2013; Raja et al., 2010; Zamanian et al., 2005).
The phylogenetic tree (diagram) revealed that the local isolate is closely related Streptomyces griseoviridis, similarity matrix is 95% as identified strain of Streptomyces plicatus (strain 101) by Zamanian et al. (2005) and Streptoverticillium sp. and two Streptomyces sp. by Raja et al. (2010). In view of all the previously recorded data, the identification of actinomycete isolate KSA-20 was suggestive of being belonging to Streptomyces griseoviridis, KSA-20, which can produce a broad-spectrum antibacterial agents as previously reported (Ghadin et al., 2008; Ubukata et al., 2007).
The species under investigation belonging to the genus Streptomyces constitute 50% of the total population of soil actinomycetes and 75-80% of the commercially and medicinally useful antibiotics have been derived from this genus (Mellouli et al., 2003). The organic phase was collected and evaporated under reduced pressure using a rotary evaporator. The extract was concentrated and treated with petroleum ether (b.p. 40-60°C) for precipitation process where only one fraction was obtained in the form of yellowish ppt. and then tested for their antimicrobial activity. Separation of antibiotic into individual components has been tried by TLC using a solvent system composed n-butanol-acetic acid-water (3:1:1, v/v); as developing solvent (Atta et al., 2009; Zhang et al., 2007).
For the purpose of purification process, the antibiotic were allowed to pass through a column chromatography packed with silica gel and eluting solvent was composed of chloroform and methanol (10:2 v/v), fifty fractions were collected and tested for their activities. The most active fractions against the tested organisms ranged between 24 to 30. Similarly, many workers used a column chromatography packed with silica gel and an eluting solvent composed of various ratios of chloroform and methanol (Sekiguchi et al., 2007). The compound under investigation is freely soluble in chloroform, ethyl acetate, n-butanol, acetone, ethyl alcohol, methanol and 10% isopropyl alcohol but insoluble in petroleum ether, hexan and benzene; similar results were recorded (Atta, 2010; El-Tayeb et al., 2004; Mellouli et al., 2003).
Isolation of antimicrobial agents from Streptomyces griseoviridis isolated from Al-Khurmah governorate showed remarkable antimicrobial activity against tested organisms. This isolate was identified according to the recommended keys as Streptomyces griseoviridis KSA-20 as antimicrobial agents isolated from actinomycetes from EL-Taif area, Kingdom of Saudi Arabia (Al-Humiany, 2011).
In this investigation, a study of the elemental analysis of the antibiotic showed C = 55.33%, H = 5.70%, N = 8.80%, O = 23.45% and S = 6.71%. This analysis indicates a suggested empirical formula of C22H27N3O7S. The spectroscopic characteristics of antibiotic revealed the presence of the maximum absorption peak in UV at 221 nm, infra-red absorption spectrum showed characteristic band corresponding to 26 peaks. Mass-spectrum showed that the molecular weight is 477.4. The MIC of antibiotic under study exhibited fairly active against both Gram positive and Gram negative bacteria. Similar investigations and results were attained (Atta et al., 2009; Inagaki et al., 2006; Sekiguchi et al., 2007). Identification of antibiotic according to recommended international keys indicated that the antibiotic is suggestive of being belonging to griseoviridin antibiotic (Berdy, 1974; Umezawa, 1977).
CONCLUSION
The present study mainly involved in the isolation of actinomycetes based on the cultural, morphological, physiologcial and biochemical characteristics, as well as 16S rRNA methodology. Further work should be focused in most potent Streptomyces isolate for production the antibacterial activities against Gram positive and Gram negative bacteria and studies parameters controlling the biosynthetic process of antibacterial agent formation. The bioactive substance was suggestive of being belonging to griseoviridin antibiotic.
ACKNOWLEDGMENT
This research was supported by a program to support research and researchers at King Khalid University, Saudi Arabia No. (KKU_S133_33).
REFERENCES
- Afifi, M.M., H.M. Atta, A.A. Elshanawany, U.M. Abdoul-Raouf and A.M. El-Adly, 2012. Biosynthesis of hygromycin-B antibiotic by Streptomyces crystallinus AZ151 isolated from Assuit, Egypt. Bacteriol. J., 2: 46-65.
CrossRefDirect Link - Afifi, M.M., H.M. Atta, A.A. Elshanawany, U.M. Abdoul-Raouf and A.M. El-Adly, 2012. Identification of luteomycin like antibiotic produced by Streptomyces tanashiensis AZ-C442 isolated from luxor governorate at Upper Egypt. Microbiol. J., 2: 1-22.
CrossRefDirect Link - Ahmed, F. and W.A. Donaldson, 2007. Chemistry and biology of the streptogramin a antibiotics. Mini-Rev. Org. Chem., 4: 159-181.
Direct Link - Al-Humiany, A.A.A., 2011. Taificidin1 and taificidin2, two anti-microbial agents isolated from the fermentation broth of Streptomyces roseodistaticus TA15 and Streptomyces lavendofoliae TA17. Res. J. Microbiol., 6: 328-342.
CrossRef - Atta, H.M., 2010. Production, purification, Physico-chemical characteristics and biological activities of antifungal antibiotic produced by Streptomyces antibioticus, AZ-Z710. Am.-Eur. J. Sci. Res., 5: 39-49.
Direct Link - Atta, H.M., M.H. El-Sehrawi, N.M. Awny and N.I. El-Mesady, 2011. Cirramycin-B antibiotic production by Streptomyces cyaneus-AZ-13Zc: Fermentation, purification and biological activities. N. Y. Sci. J., 4: 35-42.
Direct Link - Barriere, J.C., N. Berthaud, D. Beyer, S. Dutka-Malen, J.M. Paris and J.F. Desnottes, 1998. Recent development in Streptogramin research. Curr. Pharm. Dis., 4: 155-180.
PubMed - Becker, B., M.P. Lechevalier, R.E. Gordon and H.A. Lechevalier, 1964. Rapid differentiation between Nocardia and Streptomyces by paper chromatography of whole-cell hydrolysates. Applied Microbiol., 12: 421-423.
PubMed - Berdy, J., 1974. Recent developments of antibiotic research and classification of antibiotics according to chemical structure. Adv. Applied Microbiol., 18: 309-406.
PubMed - Chapman, G.H., 1952. A simple method for making multiple tests of a micro-organism. J. Bacteriol., 63: 147-149.
PubMed - Cuppels, D.A., J. Higham and J.A. Traquair, 2013. Efficacy of selected streptomycetes and a streptomycete + pseudomonad combination in the management of selected bacterial and fungal diseases of field tomatoes. Biol. Control, 67: 361-372.
CrossRef - Dhananjeyan, V., N. Selvan and K. Dhanapal, 2010. Isolation, characterization, screening and antibiotic sensitivity of actinomycetes from locally (Near MCAS) collected soil samples. J. Biol. Sci., 10: 514-519.
CrossRefDirect Link - Edwards, U., T. Rogall, H. Blocker, M. Emde and E.C. Bottger, 1989. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucl. Acids Res., 17: 7843-7853.
CrossRefPubMedDirect Link - El-Tayeb, O.M., A.A. Salama, M.M.M. Hussein and H.F. El-Sedawy, 2004. Optimization of industrial production of rifamycin B by Amycolatopsis mediterranei I. The role of colony morphology and nitrogen sources in productivity. Afr. J. Biotechnol., 3: 266-272.
Direct Link - Ghadin, N., N.M. Zin, V. Sabaratnam, N. Badya, D.F. Basri, H.H. Lian and N.M. Sidik, 2008. Isolation and characterization of a novel endophytic Streptomyces SUK 06 with antimicrobial activity from Malaysian plant. Asian J. Plant Sci., 7: 189-194.
CrossRefDirect Link - Gordon, R.E., 1966. Some criteria for the recognition of Nocardia madurae (vincent) blanchard. J.Gen. Microbiol., 45: 355-364.
CrossRefPubMedDirect Link - Gordon, R.E., D.A. Barnett, J.E. Handerhan and C.H.N. Pang, 1974. Nocardia coeliaca, Nocardia autotrophica and the Nocardin strain. Int. J. Syst. Evolut. Microbol., 24: 54-63.
CrossRef - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hankin, L., M. Zucker and D.C. Sands, 1971. Improved solid medium for the detection and enumeration of pectolytic bacteria. Applied Microbiol., 22: 205-209.
Direct Link - Hozzein, W.N., M.I.A. Ali and W. Rabie, 2008. A new preferential medium for enumeration and isolation of desert actinomycetes. World J. Microbiol. Biotechnol., 24: 1547-1552.
CrossRefDirect Link - Jones, K.L., 1949. Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol., 57: 141-145.
Direct Link - Khucharoenphaisan, K., N. Sripairoj and K. Sinma, 2012. Isolation and identification of actinomycetes from termite's gut against human pathogen. Asian J. Anim. Vet. Adv., 7: 68-73.
CrossRefDirect Link - Lechevalier, M.P. and H. Lechevalier, 1970. Chemical composition as a criterion in the classification of aerobic actinomycetes. Int. J. Syst. Evol. Microbiol., 20: 435-443.
CrossRefDirect Link - Mellouli, L., R.B. Ameur-Mehdi, S. Sioud, M. Salem and S. Bejar, 2003. Isolation, purification and partial characterization of antibacterial activities produced by a newly isolated Streptomyces sp. US24 strain. Res. Microbiol., 154: 345-352.
CrossRefDirect Link - Muharram, M.M., M.S. Abdelkader and S.I. Alqasoumi, 2013. Antimicrobial activity of soil actinomycetes isolated from Alkharj, KSA. Int. Res. J. Microbiol., 4: 12-20.
Direct Link - Nitsch, B. and H.J. Kutzner, 1969. Egg-yolk agar as a diagnostic medium for streptomycetes. Experientia, 25: 220-221.
CrossRefDirect Link - Pridham, T.G. and D. Gottlieb, 1948. The utilization of carbon compounds by some Actinomycetales as an aid for species determination. J. Bacteriol., 56: 107-114.
Direct Link - Raja, A., P. Prabakaran and P. Gajalakshmi, 2010. Isolation and screening of antibiotic producing psychrophilic actinomycetes and its nature from rothang hill soil against viridans Streptococcus sp. Res. J. Microbiol., 5: 44-49.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sanger, F., S. Nicklen and A.R. Coulson, 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A., 74: 5463-5467.
CrossRefPubMedDirect Link - Sekiguchi, J.I., T. Miyoshi-Akiyama, E. Augustynowicz-Kopec, Z. Zwolska and F. Kirikae et al., 2007. Detection of multidrug resistance in Mycobacterium tuberculosis. J. Clin. Microbiol., 45: 179-192.
CrossRef - Ubukata, M., N. Shiraishi, K. Kobinata, T. Kudo and I. Yamaguchi et al., 2007. RS-22A, B and C: New macrolide antibiotics from Streptomyces violaceusniger. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot (Tokyo), 48: 289-292.
PubMed - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Umezawa, H., 1977. Recent advances in bioactive microbial secondary metabolites. Jpn. J. Antibiot., 30: 138-163.
PubMed - Williams, S.T. and F.L. Davies, 1965. Use of antibiotics for selective isolation and enumeration of actinomycetes in soil. J. Gene. Microbiol., 38: 251-261.
CrossRefDirect Link - Zamanian, S., G.H. Shahidi Bonjar and I. Saadoun, 2005. First report of antibacterial properties of a new strain of Streptomyces plicatus (strain 101) against Erwinia carotovora from Iran. Biotechnology, 4: 114-120.
CrossRefDirect Link - Zhang, L.J., Z.H. Jin,, X.G. Chen, Q.C. Jin and M.G. Feng, 2012. Glycine feeding improves pristinamycin production during fermentation including resin for in situ separation. Bioprocess Biosyst. Eng., 35: 513-517.
CrossRef