
P.C. Xalxo
Parasitology Lab, School of Studies in Life Sciences, Pandit Ravishankar Shukla University, Raipur, Chhattisgarh, India
D. Karkun
Department of Botany,VYT Autonomous PG College, Durg, Chhattisgarh, India
A.N. Poddar
Parasitology Lab, School of Studies in Life Sciences, Pandit Ravishankar Shukla University, Raipur, Chhattisgarh, India
Research Journal of Microbiology
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 81-91







ABSTRACT
The root knot nematode (Meloidogyne spp.) is a polyphagous plant pest causing losses in a wide range of host crops. Due to environmental health concerns, the search for new microbial strains as nematode control agents is relevant. Soil samples were collected from the rhizoshere of nematode infested and non infested cucurbits plants and fungi isolated and identified using standard methods. Extraction of nematodes was done using Cobb’s sieving and decanting method and modified Baerman’s funnel technique. Cell free Fungal Culture Filtrates (FCF) were prepared from the most frequently occurring isolates and their nematicidal potentials assessed as the mean percentage mortality of nematodes at an interval of 24, 48 and 72 h. Sterile distilled water served as control. In all 18 fungal species were isolated from the rhizosphere soil of Momordica charantia control and 08 species from that of Momordica charantia infected and 09 species from Lagenaria siceraria infected. Among the nine fungi tested for nematicidal properties, Aspergillus flavus showed maximum nematode mortality (45%) after 24 h. After 48 h maximum nematode mortality was shown by Aspergillus species 2 and Trichoderma viride (68.75%). After 72 h, maximum nematode mortality was observed due to Trichoderma viride and Aspergillus flavus (93.75%). The results suggest that the various fungi associated with the rhizosphere soil of different vegetable crops could be used as a biocontrol agent of nematodes and the culture filtrates can serve as a source of novel nematicidal compound of fungal origin which is more environmental friendly.
PDF Abstract XML References Citation
Received: June 11, 2013;
Accepted: September 02, 2013;
Published: October 25, 2013
How to cite this article
P.C. Xalxo, D. Karkun and A.N. Poddar, 2013. Rhizospheric Fungal Associations of Root Knot Nematode Infested Cucurbits:
In vitro Assessment of Their Nematicidal Potential. Research Journal of Microbiology, 8: 81-91.
URL: https://scialert.net/abstract/?doi=jm.2013.81.91
URL: https://scialert.net/abstract/?doi=jm.2013.81.91
INTRODUCTION
The rhizosphere is an important biotic zone of interaction and is the residence of a large variety of microbes such as, bacteria, actinomycetes, fungi, protozoa and micro algae, besides presence of higher densities of nematodes and mites (Bazin et al., 1990). The root knot nematode (Meloidogyne spp.) is a cosmopolitan, polyphagous plant pest causing losses in a wide range of host crops. Since, indiscriminate use of nematicides is responsible for environmental and human health concerns; the search for new microbial strains as nematode control agents is relevant (Kerry, 2000; Regaieg et al., 2010). Since, fungi cohabit together with nematodes in the rhizosphere, their toxic metabolites may be responsible for keeping a low level of nematode populations (Siddiqui et al., 2001; Le et al., 2009; Zarina and Shahina, 2010). Thus, fungi may be considered as important in nematode control due to their ability for production of nematotoxic (Hallmann and Sikora, 1996; Kerry, 2000; Regaieg et al., 2010; Lopez-Llorca and Jansson, 2006) and nematicidal compounds (Hallmann and Sikora, 1996; Anke and Sterner, 1997; Chen et al., 2000; Meyer et al., 2000, 2004).
Specific secondary metabolites or enzymes show ing nematode antagonistic or toxic properties are a promising research field (Shinya et al., 2008; Regaieg et al., 2010). In recent years, the search for nematotoxic or antagonistic compounds in culture filtrates has been greatly intensified (Liu et al., 2008; Lopez-Llorca et al., 2008). Assays with culture filtrates show toxic and inhibitory effects towards plant parasitic nematodes (Nitao et al., 1999; Chen and Chen, 2002; Olivares-Bernabeu and Lopez-Llorca, 2002; Mukhtar and Pervaz, 2003; Khan et al., 2004). Toxic effects of fungal culture filtrates on M. incognita have been studied by several workers (Hallmann and Sikora, 1996; Anke and Sterner, 1997; Sharma, 1999; Wang et al., 1999; Costa et al., 2000; Randhawa et al., 2001; Meyer et al., 2004; Zareen et al., 2001; El-Shanshoury et al., 2005), although a broad range of fungi still remain unexplored. Hence, this work was undertaken with the objectives to isolate rhizospheric fungal associations of root knot nematode infected and uninfected Cucurbit plants and invitro assessment of their nematicidal potential against the Meloidogyne spp.
MATERIALS AND METHODS
Collection of samples: Soil samples (nematode infested and non infested) were collected from the rhizosphere of four species of cucurbits plants, namely Lagenaria siceraria, Cucumis sativus, Cucurbita pepo and Momordica charantia from a depth of 6 inches, at the rate of one unit sample per acre crop area. Each unit sample was a composite of 20 cores obtained from four corners and centre of the field. Two hundred gram of soil sub-samples prepared from the unit samples were stored in polythene bags and stored at 4°C in a refrigerator for not more than one week (Goodey, 1957).
Fungal isolation and identification: Rhizospheric fungi of the various vegetable crops were isolated by using soil plate method (Warcup, 1950) and Serial dilution method (Aneja, 2003). Materials from the pure culture were mounted in Lactophenol, stained with Cotton blue and the morphological observations of hyphae, sporangiophore/conidiophore and conidia were done with the help of a Compound light microscope, Labomed CXL Mono, at 100X, 400X and 1000X magnifications.
Preparation of cell free Fungal Culture Filtrate (FCF): To test the nematicidal potential of the cell free Fungal Culture Filtrate (FCF) the most frequently occurring isolates belonging to the genera of Aspergillus, Trichoderma and Fusarium were selected. These strains were inoculated on to petri plates containing Potato Dextrose Agar medium and incubated for 7 to 10 days at 27°C, which served as inocula. From these actively growing cultures, one disc each of 0.5 cm diameter was transferred to 250 mL Erlenmeyer flask containing 50 mL Potato Dextrose broth. These flasks were incubated in a rotary incubator at 27±1°C and 150 rpm for 15 days. After 15 days of incubation, the cell free culture filtrate was obtained by filtering the metabolized growth medium through pre weighed Whatmann filter paper. The filtrates were used in their original concentrations to test their toxicity against nematodes.
Extraction of root knot nematode from rhizosphere: Soil samples from the rhizosphere of nematode infected cucurbits were collected and processed for nematode extraction using Cobb’s sieving and decanting method and modified Baerman’s funnel technique (Southey, 1985). Populations of nematodes were counted microscopically in specially designed counting dishes.
Evaluation of nematicidal potential of fungal culture filtrates: The nematode suspension and fungal culture filtrate were pipetted into sterile multichambered counting dishes at the ratio of 1: 3 and number of nematodes counted at 0 h. The experiment was set in triplicates. The counting chambers were kept for incubation at room temperature in a moist chamber. The mortality was assessed as the mean percentage of dead (immobilized) nematodes at an interval of 24, 48 and 72 h. Sterile distilled water served as control (Radwan, 2007).
RESULTS
Fungal isolation and identification: The distribution of fungal species in the rhizosphere of Cucurbits uninfected and infected with root knot nematodes is shown in Table 1.
Twenty seven different fungal isolates belonging to various genera were isolated from six rhizosphere soil samples of four cucurbit plant species by using Serial dilution method and Warcup method.
| Table 1: | Distribution of fungal species in the rhizosphere of cucurbits uninfected and infected with Root Knot Nematodes (RKN) |
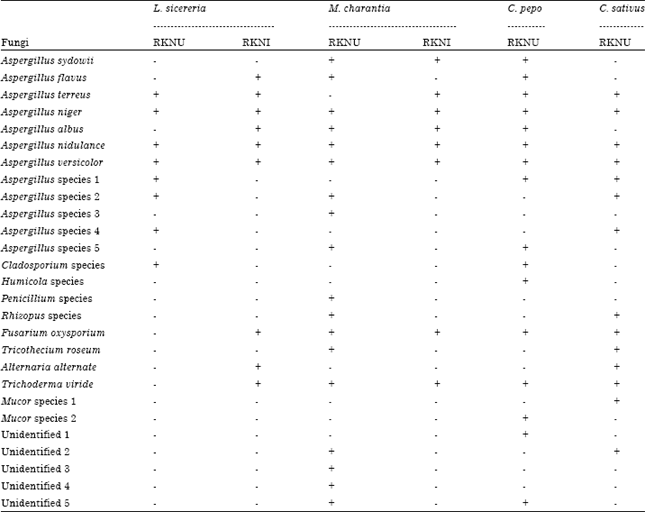 | |
| RKNU: Uninfected with root knot nematode, RKNI: Infected with root knot nematode, Present (+), Absent (-), L. sicereria (Lageneria sicereria), M. charantia (Momordica charantia), C. pepo (Cucurbita pepo), C. sativus (Cucumis sativus) | |
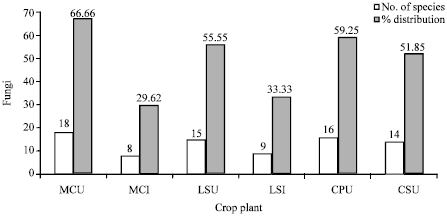 | |
| Fig. 1: | No. and % distribution of fungal species associated with rhizosphere of root knot nematode infected and uninfected Cucurbits, MC: M. charantia, LS: Lageneria sicereria, CP: Cucurbita pepo, CS: Cucumis sativus, Infected with Root Knot Nematode (I), Uninfected with Root Knot Nematode (U) |
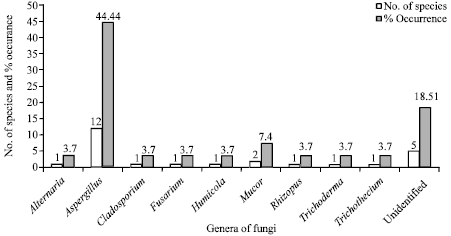 | |
| Fig. 2: | Percent occurrence of various fungal genera in the rhizosphere of cucurbits |
Twenty two species of moulds belonging to thirteen genera were identified. Five isolates could not be identified as they lacked sporulation in the conditions that were provided. Most were representatives of the class Deuteromycetes, although members of Zygomycetes and Ascomycetes were also found. The highest numbers of fungal species (18) were isolated from the rhizosphere soil of Momordica charantia uninfected with root knot nematodes, while 08 species were recorded from those of infected ones. Fifteen species were recorded from rhizosphere of uninfected Lagenaria siceraria as compared from 9 species from infected ones. Sixteen and fourteen species were respectively recorded from the rhizosphere of uninfected Cucurbita pepo and Cucumis sativus. Percent distribution of fungi was highest in M. charantia (66.66) followed by C. pepo (59.25), L. sicereria (55.55) and C. sativus (51.85) (Fig. 1). The most frequent genus was Aspergillus accounting 44.4% of the total mould isolates with Aspergillus niger being the most frequent and abundant species. The second most common genera were Trichoderma followed by Rhizopus and Fusarium (Fig. 2).
The various filamentous fungi isolated were identified, based on the following characteristics in accordance with the standard literature available.
Aspergillus sydowii: Colonies on Czapek’s agar were blue-green, with the bluish effect prominent; velvety, growing well, attaining 3-4 cm diameter after ten days. Reverse and substratum had shades of orange to red, becoming almost black. Conidiophores mostly arose from submerged hyphae, up to 500x4-8 μm, colourless, smooth and thick-walled. Heads radiate or globose; vesicles 12-20 μm in diameter. Phialides radiated in two series; conidia globose 2.5-3.5 μm.
Aspergillus flavus: Colonies on Czapek’s agar widely spreading, growing rapidly, 7.5-8.5 cm to slow growing, 3-4 cm in 10 days; conidial heads yellow when young, becoming dark yellow-green in age, in older cultures deep grey-green, reverse colourless to pale yellow brown. Conidiophores arose separately from the substratum, 400-700 or 1000 μm longx5-15 μm in diameter, broadening upward. Walls were pitted as to appear rough or spiny with low magnification, occasionally granular, gradually enlarging upward to form a vesicle 10-30 or 40 μm in diameter.
Aspergillus fumigatus: Colonies on Czapek’s agar spread rapidly, white at first becoming dull blue-green, velvety to floccose; reverse colourless. Conidial heads columnar, compact often densely crowded, up to 400x50 μm. Conidiophores short, smooth, light green, up to 300 μm in length and 5-8 μm in breadth, septate, gradually enlarging into a flask shaped vesicle. Vesicles , fertile on the upper 1/2 to 2/3, 20-30 μm in diameter, bearing a single series of phialides. Phialides closely packed, 6-8x2-3 μm; conidia globose to sub globose, 2.5-3 μm, green in mass.
Aspergillus niger: Colonies on Czapek’s agar rapidly growing with abundant submerged mycelium, attaining a diameter of 8-9 cm in 10 days. Conidial heads carbon black or sometimes deep brownish black. Reverse colourless to pale yellow. Conidial heads large and black, at first globose, then radiate or splitting, in well defined columns. Conidiophores arising directly from the substratum, smooth, non septate, thick walled, 1-2 mmx15-20 μm, vesicles globose, walls thick, commonly 45-75 μm in diameter, occasionally longer, bearing two series of fully packed phialides, conidia globose, at first smooth but later spinulose with colouring substance, black, 4-5 μm.
Aspergillus terreus: Colonies on Czapek’s agar form tints of pinkish-cinnamon through cinnamon to deeper brown shades in age, velvety, spreading well, attaining 3.5-5 cm in 10 days. Reverse pale yellow or fairly deep brown; exudates amber coloured; conidial heads long columnar, compact with uniform diameter, cinnamon brown, 150-500x30-50 μm, conidiophores smooth, colourless, 100-250x4.5-6 μm, vesicles hemispherical, dome like, 10-16 μm in diameter, phialides biseriate; conidia globose to subglobose, 1.8-2.4 μm.
Alternaria alternata: Colonies grew rapidly on PDA, reaching 7.5-8.0 in 10 days at 27°C, olive black without aerial mycelium. Conidiophores short, simple, straight, branched or unbranched up to 50 μm long Conidia often in long branched chains of 2-10 or more, uniform with 3-8 transverse septa, longitudinal or oblique septa present.
Fusarium oxysporum: Colonies reaching 4.5 cm diameter in 4 days at 25°C on PDA; white or peach but usually with a purple or violet tinge; reverse colourless, blue to dark purple; conidiophores unbranched, monophialidic. Stroma white, plectenchymatous, smooth, effuse; microconidia usually abundant, mostly zero septate, oval, ellipsoidal, kidney shaped or straight, produced on simple lateral phialides, solitary on free conidiophores, never formed chains. 5-12x2.5-3.5 μm; Macroconidia 2-5 septate, spindle to fusiform, curved or almost straight, pointed at both ends, 27-60x3-5 μm.
Humicola species: Colonies effuse, cottony, at first white, later grey brown with age. Mycelia, superficial and immersed, becoming 4 cm diameter in 10 days. Hyphae, hyaline, 0.5-2.5 μm wide, bearing masses of chlamydospores and conidia. Conidia white at first, rough, thick walled, typically globose, one celled.
Mucor species: Colonies fast growing, filling petridish in 3-5 days, often several cm high, white to yellow, becoming dark grey with the development of sporangia. Mycelia present in and on the substratum but without rhizoids or stolons. Sporangiophores forming a thick turf, springing from the mycelium, either unbranched with terminal sporangia or branched.
Rhizopus species: Colonies very fast growing, 1-2 cm high, white at first, pale, grey, dark brown-grey, reddish brown, brownish black. Mycelium branched, non septate, stolons and rhizoids present; sporangiophores arise from these points, sporangia globose, hemispherical, zygospores round or oval.
Trichoderma viride: Colonies grew rapidly up to 9 cm in 4 days, watery white, becoming hairy from the formation of loose scanty aerial mycelium, floccose to arachnoid; somewhat whitish; Conidiation effuse or in compact tufts; reverse uncolored; chlamydospores common, intercalary or terminal. Conidiophores much branched; arise in compact or loose tufts. Main conidiophores 4-5 μm, producing smaller side branches. Conidia globose or short obovoid, or broadly ellipsoidal, bluish green to dark green, 3.5-4.5 μm or 4-4.8x3.5-4 μm.
Trichothecium roseum: Colonies growing fast, reaching 7-8 cm diameter in 10 days, pinkish, zonate in diurnal rhythm, powdery due to conidial formation; reverse colourless to light pink; exudates lacking; conidiophores erect, produced singly or in groups, mostly simple, hyaline and septate. Conidia ellipsoidal to pyriform, two celled, 12-23x8-10 μm, smooth, thick walled, hyaline.
Cladosporium cladosporioides: Colonies reaching 3-5 cm diameter in 10 days at 27°C, velvety, light olive green to olive green; reverse olivaceous black; conidiophores macronematous, light olivacious brown, smooth, without swellings and sympodial elongations, branching acropleurogenously, producing conidia in chains below septum, 35-340x2-6 μm. Conidia formed in long branched chains, aseptate, ellipsoidal, olive green, olivacious brown, 3.5-10.2x2.6-3.8 μm.
Aspergillus nidulance: Colonies fast growing, reaching 6-7 cm diameter in 7 days, smooth walled conidiophore, small echinulate conidia 3-4 μm in diameter. Quickly ripening ascospores, purple red in color and with equatorial binding and large thick walled, globose bodies, termed “hulle cells” forming an irregular layer about the perithecia.
Aspergillus albus: Colonies on Czapek Dox agar fast growing, velvety, felted or floccose, plane, attaining a diameter of 4.0-8.5 cm in 4 to 12 days at 25-35°C. Reverse colorless to bright yellow, green, dark red, brown or even black in age. Conidiophore short, sometimes faintly yellow, upto 575.0 μm to 2.8 to 10.0 μm, conidia globose to subglobose.
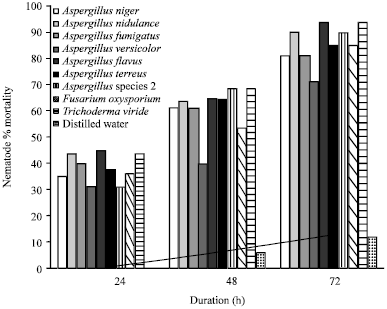 | |
| Fig. 3: | Percent mortality of nematodes in response to culture filtrates of various fungal genera |
Aspergillus versicolor: Colonies on Potato Dextrose Agar (PDA) at 25°C grew rapidly, reaching 21-23 mm in diameter after 7 days and 47-49 mm after two weeks, with sometimes centrally rising, velvety to rather floccose. Colour blue-green, exudates absent and reverse pale yellowish. It was interesting to find that a large number of heads were fragmentary and like penicillum. Conidiophores monoverticillate or biverticillate, hyaline, smooth-walled, 7.0-9.0x2.0-3.0 μm. Metulae 7.0-10.0x2.0-3.0 μm long. Phialides, 7.0-8.0x2.0-3.0 μm long, flask-shaped.
In vitro assessment of nematicidal property of fungal filtrates (Fig. 3): Among the nine fungi tested for nematicidal properties, the filtrate of Aspergillus flavus showed maximum mortality (45%) after 24 h. After 48 h maximum mortality was shown by Aspergillus species 2 and Trichoderma viride (68.75%). After 72 h, maximum mortality was reported by filtrates of Trichoderma viride and Aspergillus flavus (93.75%). Minimum activity was exhibited by Aspergillus species 2 and Aspergillus versicolor after 24 h (31.25%) and after 48 h minimum mortality in Aspergillus versicolor and Fusarium oxysporium (40 and 53.75%). After 72 h minimum mortality was reported by filtrate of Aspergillus versicolor (71.25%). The mortality rate of filtrate of Fusarium oxysporum was only 36.25% after 24 h but showed 85% mortality after 72 h. All the fungal filtrates tested showed some percentage of toxicity as compared to the control experiment using distilled water which showed zero mortality after 24 h and 6.25% mortality after 48 h and 12.5% mortality after 72 h.
DISCUSSION
Results show that the highest numbers of fungal species were isolated from the rhizosphere soil of Momordica charantia control (18 species). Lowest number was recorded from the rhizosphere of Momordica charantia infected (08 species) and Lagenaria siceraria infected (09 species). This result is in accordance with that of Altalhi (2004) and Abou-Zeid and El-Fattah (2007). Altalhi (2004) reported that the highest numbers of fungal colonies were recorded in the rhizosphere soil of Artemisia judaica and the lowest number in Desmostachya bipinnata. According to Abou-Zeid and El-Fattah (2007) the highest number of fungal populations were found in the rhizosphere of Asphodelus aestivus followed by Pulicaria crispa and the lowest numbers were reported on the rhizosphere of Argemone ochroleuce.
Twenty seven different fungal isolates belonging to various genera were isolated from six rhizosphere soil samples of four cucurbit plant species by using Serial dilution method and Warcup method. Most frequent genera was Aspergillus accounting 44.4% of the total mold isolates and Aspergillus niger, the most frequent and abundant species. This finding supports the observation made by Abdel-Hafez (1982a). According to their study, Aspergillus contributed the broadest spectrum of species in the rhizosphere and Aspergillus niger the most frequent fungi. These results are also agreeable with Abdel-Hafez (1981, 1982b) who isolated different fungal genera and species from the desert soils in Saudi Arabia. The present results also agree with El-Abyad et al. (1982) and Abdel-Hafez and Shoreit (1985) who isolated 70 species and 31 genera from 20 dust samples of Taif, Saudi Arabia and the most common genera were Aspergillus, Fusarium, Penicillium and Mucor.
In the present study the second most common genera was Mucor. Hence, it can be concluded that the frequency and type of fungal species vary in different cucurbit plants, both control and infected.
The effect of culture filtrates on nematodes differed according to nematode and fungal biological groups as well as exposure time. The nematicidal activity of culture filtrate from the most frequently occurring isolate nine fungal species the filtrate of Aspergillus flavus showed maximum mortality (45%) after 24 h. After 48 h maximum mortality Trichoderma viride and Aspergillus species showed (68.75%). These result also agreed with Siddiqui et al. (2004) and Zareen et al. (2001). After 72 h maximum mortality was reported by filtrates of Aspergillus flavus and Trichoderma viride (93.75%). Although, Trichoderma spp. are known to suppress plant-parasitic nematodes (Sharon et al., 2001; Jegathambigai et al., 2011), in this study its fungal culture filtrate had a toxic effect on M. incognita. Minimum activity was exhibited by Aspergillus species and Aspergillus versicolor after 24 h (31.25%) and after 48 h minimum mortality was reported by Fusarium oxysporium (53.75%). After 72 h minimum mortality was reported by filtrate of Aspergillus versicolor. Hence, it may be expressed undoubtedly that the culture filtrates of Aspergillus species different have a toxic effect on M. incognita similar to the results of Siddiqui et al. (2004) and Amer-Zareen et al. (1999). In contrast to the finding by Radwan (2007), filtrates of Aspergillus niger showed only 35% mortality after 24 h and 81.25% after 72 h. According to his study, filtrate of Aspergillus niger exhibited 100% mortality after 12 h in original concentration and Fusarium oxysporum showed 52% mortality after 12 h and 78.8% after 48 h. According to present investigation, the mortality rate of filtrate of Fusarium oxysporum was only 36.25% after 24 h but showed 85% mortality after 72 h. However, all the fungal filtrates tested showed some percentage of toxicity as compared to the control experiment using distilled water which showed zero mortality after 24 h and 6.25% mortality after 48 h and 12.5% mortality after 72 h (Fig. 3). The nematicidal efficacy of each fungal filtrate increased with duration of exposure.
CONCLUSION
The variations in the nematicidal action of the tested fungal filtrates may be attributed to the differences in number and nature of metabolites produced by different fungi and would be worth investigating. The result of the present study suggests that the various fungi associated with the rhizosphere soil of different vegetable crops could be used as a biocontrol agent of nematodes and the culture filtrates can serve as a source of novel nematicidal compound of fungal origin which is more environmental friendly. However, the effective components of the fungal metabolites, which are responsible for the nematicidal activity in the filtrate, need to be purified and characterized.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Prof. HS Gaur, Jt. Director, IARI, New Delhi, India for providing training in nematological techniques and Prof. SK Jadhav, Head , SoS in Biotechnology, Pandit Ravishankar Shukla University, Raipur, Chhattisgarh, India for help in fungal identification. Sincere thanks are due to Prof. V.Rai, Ex-Head of the Department, School of Studies in Life Sciences, Pandit Ravishankar Shukla University, Raipur, for providing laboratory facilities. Thanks are also due to the UGC, New Delhi for providing RGN Fellowship to P.C. Xalxo.
REFERENCES
- Abdel-Hafez, S.I.I. and A.A.M. Shoreit, 1985. Mycotoxins producing fungi and mycoflora of air-dust from Taif, Saudi Arabia. Mycopathologia, 92: 65-71.
CrossRef - Abdel-Hafez, S.I.I., 1981. Halophilic fungi of desert soils in Saudi Arabia. Mycopathologia, 75: 75-80.
CrossRef - Abdel-Hafez, S.I.I., 1982. Rhizosphere and rhizoplane fungi of Triticum vulgare cultivated in Saudi Arabia. Mycopathologia, 78: 79-86.
CrossRefDirect Link - Abdel-Hafez, S.I.I., 1982. Osmophilic fungi of desert soils in Saudi Arabia. Mycopathologia, 80: 9-14.
CrossRef - Abou-Zeid, A.M. and R.I.A. El-Fattah, 2007. Ecological studies on the rhizospheric fungi of some halophytic plants in taif governorate, Saudi Arabia. World J. Agric. Sci., 3: 273-279.
Direct Link - Amer-Zareen, M.J. Zaki and A. Ghaffar, 1999. Effect of culture filtrate of fungi in the control of Meloidogyne javacnica, root knot nematodes on okra and broad bean. Pak. J. Biol. Sci., 2: 1441-1444.
CrossRef - Chen, F. and S. Chen, 2002. Mycofloras in cysts, females and eggs of the soybean cyst nematode in Minnesota. Applied Soil Ecol., 19: 35-50.
CrossRefDirect Link - Chen, S.Y., D.W. Dickson and D.J. Mitchell, 2000. Viability of Heterodera glycines exposed to fungal filtrates. J. Nematol., 32: 190-197.
Direct Link - Costa, M.J.N., V.P. Campos, L.H. Pfenning and D.F Oliveira, 2000. Pathogenicity and reproduction of Meloidogyne incognita in tomato plants (Lycopersicon esculentum) with application of fungal filtrates or plant and animal manure extracts. Nematol. Bras., 24: 219-226.
Direct Link - EL-Shanshoury, A.E.R.R., S.A. El-Sayed, Y.A.G. Mahmoud and D.M. Khalefa, 2005. Evaluation of Pochonia chlamydosporia, Paecilomyces lilacinus and Arthrobotrys dactyloide as biocontrol agents for Meloidogyne incognita under green house condition. Pak. J. Biol. Sci., 8: 1511-1516.
CrossRefDirect Link - Hallmann, J. and R.A. Sikora, 1996. Toxicity of fungal endophyte secondary metabolites to plant parasitic nematodes and soil-borne plant pathogenic fungi. Eur. J. Plant Pathol., 102: 155-162.
Direct Link - Jegathambigai, V., R.S.W. Wijeratnam and R.L.C. Wijesundera, 2011. Effect of Trichoderma viride Strain NRRL 6418 and Trichoderma harzianum (Hypocrea lixii TWC1) on Livistona rotundifolia root knot nematode Meloidogyne incognita. J. Entomol., 8: 229-239.
CrossRefDirect Link - Kerry, B.R., 2000. Rhizosphere interactions and the exploitation of microbial agents for the biological control of plant-parasitic nematodes. Ann. Rev. Phytopathol., 38: 423-441.
CrossRefPubMedDirect Link - Khan, A., K.L. Williams and H.K.M. Nevalainen, 2004. Effects of Paecilomyces lilacinus protease and chitinase on the eggshell structures and hatching of Meloidogyne javanica juveniles. Biol. Control, 31: 346-352.
CrossRefDirect Link - Le, H.T.T., J.L. Padgham and R.A. Sikora, 2009. Biological control of the rice root-knot nematode Meloidogyne graminicola on rice, using endophytic and rhizosphere fungi. Int. J. Pest Manage., 55: 31-36.
CrossRef - Liu, T., L. Wang, Y.X. Duan and X. Wang, 2008. Nematicidal activity of culture filtrate of Beauveria bassiana against Meloidogyne hapla. World J. Microbiol. Biotechnol., 24: 113-118.
CrossRefDirect Link - Meyer, S.L.F., R.N. Huettel, X.Z. Liu, R.A. Humber, J. Juba and J.K. Nitao, 2004. Activity of fungal culture filtrates against soybean cyst nematode and root-knot nematode egg hatch and juvenile motility. Nematology, 6: 23-32.
Direct Link - Meyer, S.L.F., S.I. Massoud, D.J. Chitwood and D.P. Roberts, 2000. Evaluation of Trichoderma virens and Burkholderia cepacia for antagonistic activity against root-knot nematode, Meloidogyne incognita. Nematology, 2: 871-879.
Direct Link - Mukhtar, T. and I. Pervaz, 2003. In vitro evaluation of ovicidal and larvicidal effects of culture filtrate of Verticillium chlamydosporium against Meloidogyne javanica. Int. J. Agric. Biol., 5: 576-579.
Direct Link - Nitao, J.K., S.L. Meyer and D.J. Chitwood, 1999. In-vitro assays of Meloidogyne incognita and Heterodera glycines for detection of nematode-antagonistic fungal compounds. J. Nematol., 31: 172-183.
PubMedDirect Link - Olivares-Bernabeu, C.M. and L.V. Lopez-Llorca, 2002. Fungal egg-parasites of plant-parasitic nematodes from Spanish soils. Rev. Iberoam. Micol., 19: 104-110.
Direct Link - Radwan, M.A., 2007. Comparative effects of culture filtrate of soil-borne fungi on mortality and infectivity of juveniles of Meloidogyne incognita. Indian J. Nematol., 37: 109-114.
Direct Link - Regaieg, H., A. Ciancio, N.H. Raouani, G. Grasso and L. Rosso, 2010. Effects of culture filtrates from the nematophagous fungus Verticillium leptobactrum on viability of the root-knot nematode Meloidogyne incognita. World J. Microbiol. Biotechnol., 26: 2285-2289.
CrossRefDirect Link - Sharon, E., M. Bar-Eyal, I. Chet, A. Herrera-Estrella, O. Kleifeld and Y. Spiegel, 2001. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Phytopathology, 91: 687-693.
CrossRef - Siddiqui, I.A., S.S. Shaukat and A. Khan, 2004. Differential impact of some Aspergillus species on Meloidogyne javanica biocontrol by Pseudomonas fluorescens strain CHA0. Lett. Applied Microbiol., 39: 74-83.
Direct Link - Siddiqui, I.A., Amer-Zareen, M.J. Zaki and S.S. Shaukat, 2001. Use of Trichoderma species in the control of Meloidogyne javanica root knot nematode in Okra and Mungbean. Pak. J. Biol. Sci., 4: 846-848.
CrossRefDirect Link - Wang, L., B. Yang and C. Li, 1999. Evaluation of pathogenicity of parasitic fungi to root-knot nematodes. Scientia Silvae Sinicae, 35: 41-47.
Direct Link - Warcup, J.H., 1950. The soil-plate method for isolation of fungi from soil. Nature, 166: 117-118.
PubMedDirect Link - Zarina, B. and F. Shahina, 2010. Research work carried out on the management of root-knot nematode diseases in Pakistan. Pak. J. Nematol., 28: 153-239.
Direct Link