
Mutlag M. Al-Otaibi
Department of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, P.O. Box 400, Al-Ahsa, Saudi Arabia
Najeeb S. Al-Zoreky
Department of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, P.O. Box 400, Al-Ahsa, Saudi Arabia
Hassan A. El-Dermerdash
Department of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, P.O. Box 400, Al-Ahsa, Saudi Arabia
Research Journal of Microbiology
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 70-80







ABSTRACT
Generally, fresh milk is taken as raw product with or without proper preservation. Fresh milk is subjected to bacterial contamination irrespective of its source and can cause health problems if consumed as fresh product. Camel is one of the favorite animals in the Middle East Countries. Studies on isolating active LAB from camel’s milk in Saudi Arabia are scarce. The main objective of this study was to isolate potential probiotic Lactic Acid Bacteria (LAB). Phenotypic characteristics were determined by phenotypical and biochemical tests, carbohydrate assimilation by using API 50CH strips and API 50 CHL medium and probiotic activity of isolates was determined by tolerating bile salts. The Cell Free Supernatants (CFS) were sensitive to proteolytic enzymes indicating the proteinaceous nature (i.e., bacteriocins) of inhibitors. There was no significant difference (p>0.05) in weight gain among control and treated mice. The counts (log10 CFU mL-1) of test isolates were not significantly (p>0.05) affected by bile slats in MRS broth. In conclusion, LAB isolates proved useful and can exhibit probiotic activities in albino mice which are able to prevent the adhesion of Staphylococcus aureus and to inhibit the E. coli in Gastrointestinal (GI) tract.
PDF Abstract XML References Citation
Received: April 30, 2013;
Accepted: May 15, 2013;
Published: October 25, 2013
How to cite this article
Mutlag M. Al-Otaibi, Najeeb S. Al-Zoreky and Hassan A. El-Dermerdash, 2013. Camel's Milk as a Natural Source for Probiotics. Research Journal of Microbiology, 8: 70-80.
URL: https://scialert.net/abstract/?doi=jm.2013.70.80
URL: https://scialert.net/abstract/?doi=jm.2013.70.80
INTRODUCTION
Among the various food preservation methods, fermentation is commonly used by the food industries for the production of infection free products. Indigenous raw and fermented foods are strongly linked to culture and tradition. Among the most commonly used microorganisms, Lactic Acid Bacteria (LAB) play an important role in preserving milk and milk products. Their common occurrence in foods coupled with their long historical use contribute to their acceptance as Generally Recognized As Safe (GRAS) for human consumption and also are approved for Qualified Presumption of Safety (QPS) (Liu et al., 2011; Jans et al., 2012). Lactococcus, Lactobacillus, Pedicoccus, Leuconostoc, Streptococcus, Enterococcus and Bifidobacterium are important LAB genera (Liu et al., 2011). Some researchers reported that LAB exist in many food products and are part of natural population of Gastrointestinal (GI) tract (De Vuyst and Leroy, 2007).
Presently, it is considered that LAB act against the pathogenetic organisms to improve the quality of preserved foods for human consumption (De Vuyst and Leroy, 2007; Settanni and Corsetti, 2008). Those antimicrobials produced by Gram-positive bacteria, particularly LAB, have been largely studied with the perspective of food protection against pathogenic and spoilage microorganisms (Malheiros et al., 2012). Many factors such as varying pH, growth of bacteriocins, nutrients competition and displacing pathogens affect the protective mechanism of LAB for the control of various pathogens of food in the GI. Bacteriocins are antimicrobial peptides produced among bacteria that may show varied antimicrobial spectra. Also, bacteriocin producers have the immunity to their own bacteriocins (Chen and Hoover, 2003; De Vuyst and Leroy, 2007; Settanni and Corsetti, 2008). They are also lethal to some food-borne pathogens and spoilage bacteria (Chen and Hoover, 2003; Cheikhyoussef et al., 2008). Food-borne pathogens and spoilage microorganisms such as Listeria monocytogenes, Bacillus cereus, Staphylococcus aureus and Clostridium tyrobutyricum were inhibited by bacteriocins of LAB (Alegria et al., 2010).
Among the various LAB present in preserved foods, some LAB such as probiotics Lactobacillus rhamnosus tended to reduce the adhesion and viability of adherent Staphylococcus aureus (Vesterlund et al., 2006). The health benefits of the consumption of fermented milk containing viable or non-viable LAB have been extensively documented. Particularly, it has been reported that the administration of fermented dairy products can confer enhanced resistance against infections by enteric pathogens (Millette et al., 2008).
The raw milk microbiota are an essential components of many traditional fermented milk products and play important roles during manufacturing and ripening of fermented products such as cheeses. The quantitative evolution of lactococci, lactobacilli, leuconostocs and enterococci from milk to finished products like cheese followed similar trends. In traditional fermentation, microflora dominated by LAB probably originating from the vessel surface are used in fermentation (Jans et al., 2012). In Europe, Africa and other regions of the world, traditional food such camel’s milk were explored as natural sources for LAB. However, studies on isolating active LAB from camel’s milk are scarce or lacking. Therefore, the present study was aimed to isolate active LAB from raw camel’s milk.
MATERIALS AND METHODS
Milk samples: A total of thirty five samples of raw camel's milk were obtained by manual milking of camels. The milk samples were analyzed immediately or, when necessary, stored refrigerated overnight prior to experiments.
Isolation of lab from traditional food: A 10 mL portion from each experimental milk sample was stomached with 90 mL sterile peptone water (Oxoid, UK). Serial dilutions were further prepared using the same diluent. MRS agar plates (Oxoid, UK) were spread with 0.1 mL diluted samples with a sterile glass rod. Plates were incubated anaerobically (Gas Pak) at 37°C for 48-72 h. Representative colonies were aseptically picked from plates containing 20-100 colonies. Gram-positive coccids/rods and catalase negative bacteria were considered as presumptive LAB. Isolates and indicators were maintained in the respective broth with 15% glycerol at -20°C.
Antibacterial activity of isolates: Well diffusion assay Cell-Free Supernatants (CFS) of isolates were obtained by centrifugation (9000 g, 10 min at 5°C) of cultures grown in broths (MRS) for 48 h followed by 0.45 μm membrane filtration (Nalgene, USA). MRS agar is often abbreviated to MRS, this type of bacterial growth medium is so-named by its inventors: de Man, Rogosa and Sharpe (MRS). Eight milliliters of soft agar media (MRS or BHI plus 0.75% agar) seeded with 105 CFU mL-1 of indicator bacteria was overlaid onto MRS agar plates (Nikolic et al., 2008). Wells (5 mm) were made in the agar media and filled with 100 μL of CFS. Plates were incubated at 37°C for 18-48 h. Clear inhibition zones around wells in the lawn of indicators were measured in mm.
Effect of proteases: The CFS producing inhibition zones were tested for susceptibility to α-Chymotrypsin (Sigma), Trypsin (Sigma) and Lipase (Sigma) using the well assay mentioned earlier. L. monocytogenes ATCC 7644 was the indicator. Each enzyme (5 mg mL-1) was separately spotted (20 μL) adjacent to the edge of well prior to incubation at 37°C for 20-48 h. Untreated CFS were used as control. The activity of isolates Enterococcus (LG), Lactococcus (C8) and Lactobacillus (F3) was determined (well assay) to evaluate effects of proteolytic enzymes on the antagonistic activity of their CFS.
Activity of isolates in packaging materials: The method of Ercolini et al. (2010) with some modifications was adopted for evaluating the inhibition activity of CFS of isolates (LG, F3 and C8) against the pathogen L. monocytogenes ATCC 7644. Plastic discs of 1 cm diameter were aseptically dipped in CFS of isolates and left to dry under ambient temperatures. Discs were aseptically put onto agar media (Tryptic soy agar +0.6% yeast extract) and plates were incubated at 37°C for up to 24 h. The activity of CFS against L. monocytogenes was also determined as described by Ercolini et al. (2010).
Phenotypic characterization of active isolates: The isolates (presumptive LAB) which produced CFS sensitive to proteolytic enzyme (bacteriocins) were characterized using the phenotypical and biochemical tests (Holt, 1977; Morandi and Brasca, 2012).
Carbohydrate assimilation: Carbohydrate fermentation tests of the above isolates were carried out by using API 50CH strips and API 50 CHL medium according to manufacturer's instructions (BioMerieux, Marcy-l'Etoile, France).
Probiotic activity of isolates
Bile tolerance: The isolates were tested for their ability as probiotics by tolerating bile salts following the procedures of some previous publications (Ricci et al., 2011; Ripamonti et al., 2011). Tolerance to bile salts was tested at 37°C by inoculation of fresh cultures in MRS broth enriched with 0.3% Oxgall (Oxoid, UK). Resistance to bile salts (Oxygall) was assessed in terms of viable count, enumerated (in MRS agar, Gas Pak at 37°C) after incubation with bile salts for 0 and 2 h.
Antibiotic resistance: Antimicrobial susceptibility was determined by the standardized agar diffusion test on MRS agar (Morandi and Brasca, 2012). Sterilized and tempered (50°C) agar media were seeded with test isolates at 105 CFU mL-1. Antibiotic discs were placed on agar surfaces. Plates were incubated for 12-48 h at 37°C. LAB isolates were screened for their susceptibility to amikacin (30 μg/disc), ampicillin (10 μg/disc), amoxicillin (30 μg/disc), bacitracin (10 IU disc-1), cefuroxime (30 μg/disc), chloramphenicol (30 μg/disc), colistin (10 μg/disc) and gentamicin (10 μg/disc). After incubation, inhibition zones were measured in mm.
Animal experiments: The impact of ingested probiotics (in vivo trails) on fecal population was evaluated by the oral administration (bottle feeding) of probiotics to animal models (albino mice). The experimental trails using mice and other necessary items for feeding were conducted in the animal laboratory of the College of Veterinary Medicine, King Faisal University (KFU). The modulation of intestinal microbiota was assessed according to the procedures of Medici et al. (2005) and Millette et al. (2008). Animal experiments (feeding trials for 3-16 days) were conducted to evaluate the health benefits (fecal microbiota modulation) of probiotic LAB (108 CFU mL-1 milk) isolated from samples. Prior to feeding experiments, mice were kept for a 14-day acclimatization period (day zero) as recommended by Shu et al. (1999).
Albino mice (25 g) were obtained from the random bred colony kept by the College of Veterinary Medicine (KFU). The treated and control mice were fed ad libitum (barley, corn, alfalfa, wheat bran, soybean meal, date syrup, vitamins and minerals) from ARASCO, Saudi Arabia. The diet contained 13% crude protein, 9% crude fiber, 1% fat (crude) and 0.7% minerals. Mice were housed in groups of five per plastic cage and kept under ambient conditions (20-23°C) in the animal laboratory.
Experimental mice were divided into six groups: (1): Control group were fed on normal diet and water (2): three groups were fed on normal diet plus diluted experimental probiotic yoghurt (108 CFU mL-1 of F3, LG or C8). Meanwhile, two additional groups of mice were fed with UHT milk (blank without probiotics) or a commercial probiotic dairy product (as reference). From preliminary trails, it was observed that one isolate from camel's milk (coded LG) caused aggressive behaviors in mice and thus was excluded from further animal experiments. LG was later identified as Enterococcus.
Experimental mice were examined at regular intervals for any signs of behavioral perturbation during the experiments. After defecation, the stool (controls, treatments and reference) was aseptically collected and tested for selected microorganisms; LAB (MRS agar), E. coli (EMB agar), Staphylococcus aureus (Baird-Parker agar with egg yolk-tellurite supplement) (Medici et al., 2005; Millette et al., 2008).
Statistical analysis: Data from triplicate experiments were evaluated for statistical significance at 5% level of significance using ANOVA according to SAS Institute (2001).
RESULTS AND DISCUSSION
The isolation and screening of microorganisms from natural sources have always been the most powerful means for obtaining useful and genetically-stable strains for industrially-important products. LAB are important in the food and dairy industries because their metabolites (bacteriocins) act as natural preservatives as well as flavor enhancers. Meanwhile, LAB find increasing acceptance as probiotics which aid in stimulating immune responses, preventing infection by enteropathogenic bacteria and treating and preventing diarrhea (Adnan and Tan, 2007; Giraffa, 2011; Jiang et al., 2012).
Presumptive LAB isolated from camel's milk samples from local farms were screened for antagonistic activity against pathogenic microorganisms (e.g., L. monocytogenes ATCC 7644, Staphylococcus aureus ATCC 29213).
Well assay: The three LAB isolates (Table 1) gave clear inhibition zones (14-25 mm) against indicators (gram positive and negative bacteria) in the well assay using the CFS. It is known that some LAB produce bacteriocins which are inhibitory for closely related gram positive bacteria (i.e., LAB) and well-known pathogens such as L. monocytogens. Up to date, nisin is a bacteriocin accepted as a safe food preservative by over 45 countries. It is the most widely used commercial bacteriocin and remains the only bacteriocin that may be added to U.S. foods. Microbes produce an extraordinary array of microbial defense systems. Metabolic by-products such as bacteriocins are loosely defined as biologically active protein moieties with a bactericidal mode of action.
| Table 1: | Inhibition zones in mm (well assay) of presumptive LAB isolates against pathogens and reference LAB |
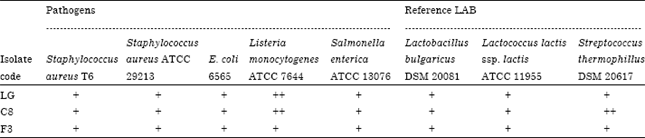 | |
| +: 14<zone<20 mm, ++: >20-25 mm | |
| Table 2: | Effect of proteolytic enzymes and lipase on inhibitive activity (well assay) of food isolates against L. monocytogenes ATCC 7644 |
 | |
| +: Inhibition zones>14-25 mm, -: No inhibition zones | |
The bacteriocin-producing strains have a natural immunity to their own bacteriocins. LAB can inhibit the growth of other bacteria by synthesizing a wide array of antimicrobials and bacteriocins. Their bio-preservation value is reflected in their inhibition of food spoilage and pathogenic microorganisms (Liu et al., 2011).
Effect of proteases: CFS of active isolates were subjected to various proteases and a lipase. The results in Table 2 show that CFS were sensitive to proteolytic enzymes indicating the proteinaceous nature (i.e., bacteriocins) of inhibitors. As expected, lipase did not affect the inhibition activity of CFS against L. monocytogenes ATCC 7644. It is stated that LAB bacteriocins are proteins and thus were degraded by proteases such as α-Chymotrypsin (Alegria et al., 2010; Chen and Hoover, 2003).
Activity of CFS in food packaging materials: Many studies revealed the antimicrobial activities of packaging films containing bacteriocins (e.g., nisin) or plant materials (e.g., cinnamaldehyde) against E. coli O157:H7, S. enterica and L. monocytogenes in poultry and meat (Ercolini et al., 2010; Ravishankar et al., 2009). Therefore, CFS of some active isolates were tested for activity in food-contacting surfaces (packaging) against L. monocytogenes ATCC 7644. As shown in Table 3, inhibitory substances for L. monocytogenes ATCC 7644 of CFS from isolates (F3, C8 and LG) gave clear zones under the plastic discs (plastic wrapping of food). Meanwhile, no inhibition halos were detected against the pathogen around the plastic discs. Evidently, this finding indicated that inhibitory substances present in CFS did not migrate from the discs (plastic packages) and thus the adsorbed components imposing only their inhibitory activity against L. monocytogenes ATCC 7644 in the area underneath the plastic films. This finding was in a complete agreement with results reported by Ercolini et al. (2010).
Phenotypic characteristics of active isolates: Some of the phenotypic characteristics of the LAB isolates are presented in Table 4. As per assumption, the LAB isolates were gram-positive, catalase negative, coccids or rods bacteria. They were homo-fermentative LAB producing only lactic acid from glucose. Lactobacillus plantarum and Lactococcus lactis were isolated from camel's milk in Kuwait (Yateem et al., 2008).
| Table 3: | Inhibitory activity against L. monocytogenes ATCC 7644 of CFS (LAB isolates) adsorbed to food packaging plastic films |
 | |
| +: Inhibition, -: No inhibition, LG: Enterococcus, F3: Lactobacillus, C8: Lactococcus | |
| Table 4: | Some characteristics of presumptive LAB isolated from camel's milk |
 | |
| -: Negative, +: Positive, c: Cocci, r: Rods, W: Weak. LG: Enterococcus, C8: Lactococcus, F3: Lactobacillus | |
Isolates lowered the pH of MRS broths and skim milk to below 5 when incubated at 22°C as a result of producing lactic acid. The three isolates (LG, F3 and C8) had no β-hemolytic activity (Table 4).
Utilization of carbohydrates: The carbohydrate assimilation (API 50) pattern of test isolates is presented in Table 5. Some isolates were identified (89-95%) as Lactococcus (C8) and Lactobacillus (F3) using the API software (BioMerieux Model Mini API).
Probiotic properties of LAB isolates: Probiotic traits (e.g., bile salt tolerance) of selected isolates showing better inhibition against the pathogen L. monocytogenes (Table 1) were evaluated as described below.
Bile tolerance of isolates: As seen in Fig. 1 and 2, counts (log10 CFU mL-1) of test isolates were not significantly (p>0.05) affected by bile slats in MRS broth. Similarly, other LAB (i.e., L. helveticus, L. paracasei and L. rhamnosus) isolated from artisanal Italian cheeses were resistant to 0.3% bile salts (Ricci et al., 2011). These results highlight the potential of the strains of dairy origin to survive under gastrointestinal conditions. Bile salts did not significantly affect the viability of Bifidobacterium longum and Lactobacillus paracasei subsp. paracasei (Ripamonti et al., 2011). Bile tolerance, being an important character, enables the probiotic strains to survive, grow and exert their beneficial effects in the host (Ricci et al., 2011; Ripamonti et al., 2011).
Antibiotic sensitivity of isolates: Table 6 lists the inhibition zones produced by the antibiotic discs. Lactococcus lactis subsp. Lactis ATCC was used as a reference.
| Table 5: | Carbohydrate assimilation of Isolates using the API 50 CH |
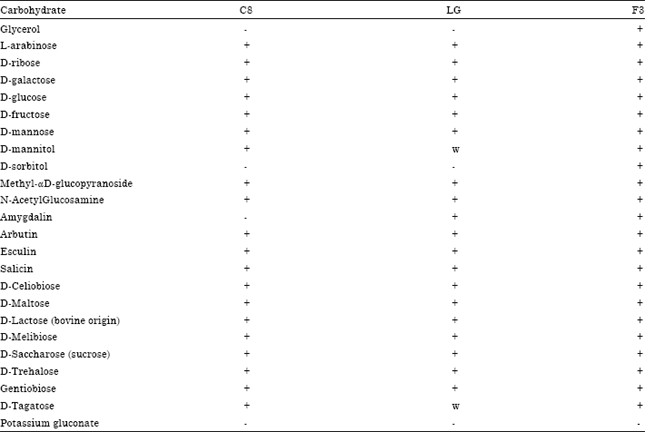 | |
| +: Positive, -: Negative, w: weak reaction, Isolates. LG: Enterococcus, F3: Lactobacillus, C8: Lactococcus | |
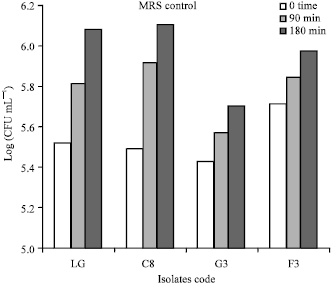 | |
| Fig. 1: | Growth (log10 CFU mL-1) of LAB isolates in MRS broth (without bile salt) during incubation at 37°C for 0 to 180 min |
All isolates were sensitive to ampicillin, amoxicillin, bacitracin and chloramphenicol. The range of some important isolates was 40, 30 and 26 of LG, C8 and F3, respectively for Ampicillin; 45, 34 and 28 of LG, C8 and F3, respectively for Amoxicillin; 25, 13 and 12 of LG, C8 and F3, respectively for Bacterian whereas as it was 30, 25 and 24 of LG, C8 and F3, respectively for Chloemphenicol.
| Table 6: | Diameter of inhibition zones (mm) for presumptive LAB isolates using disc diffusion test of 8 antibiotics |
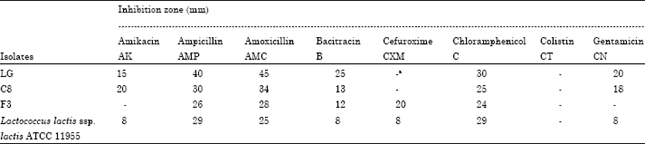 | |
| Lactococcus lactis ssp. lactis ATCC 11955, a-: No inhibition. Isolates: LG: Enterococcus, F3: Lactobacillus, C8: Lactococcus | |
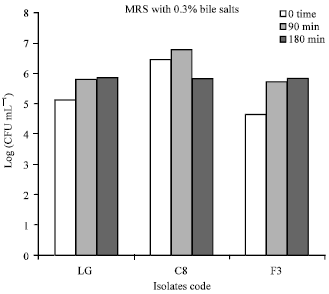 | |
| Fig. 2: | Survival (log10 CFU mL-1) of LAB isolates in MRS broth with 0.3% bile salts during incubation at 37°C for 0 to 180 min |
On the other hand, colistin had no activity against isolates. The LAB displays a wide range of natural antibiotic resistances but in most cases antibiotic resistance is not of the transmissible type. Lactobacillus strains with non-transmissible antibiotic resistances are not usually of a safety concern. It has to be considered that antibiotic resistance observed in LAB strains is often intrinsic and non-transmissible because it is chromosomally encoded (Ripamonti et al., 2011). Due to the indiscriminate use of antibiotics in human and veterinary medicine as well as in animal growth promoters, antibiotic resistance has become an increasingly common characteristic in microorganisms. However, checking the ability of a proposed probiotic strain to act as a donor of antibiotic resistance genes may be a further prudent precaution (Ricci et al., 2011).
Animal feeding experiments: Now a days, there is a growing interest in using some LAB as probiotics in food products to enhance immunity and prevent gastrointestinal infections. According to the WHO, probiotics are live microorganisms and when administered in adequate amounts confer a health benefit on the host (Giraffa, 2011; Millette et al., 2008; Vesterlund et al., 2006). Since, January 2006, the European ban of Growth Promoter Antibiotic (GPA) has resulted in an increased interest in the use of probiotics as feed additives to optimize gut health and animal performance (Ripamonti et al., 2011).
| Table 7: | Counts (log10 CFU g-1 feces) of microorganisms in mice groups fed daily with LAB isolates used as probiotics for sixteen days |
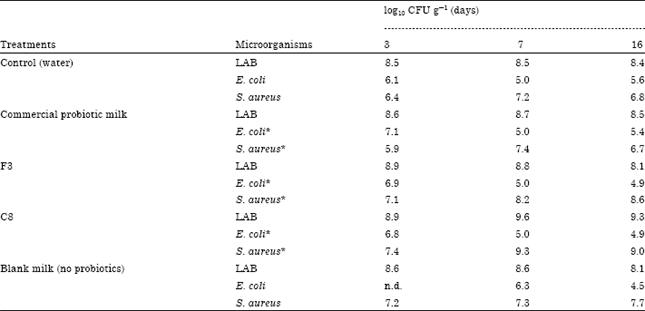 | |
| Zero day counts (log10 CFU g-1), LAB 8.2, E. coli 6.6, S. aureus 6.9. n.d.: Not done. *p<0.05. Isolates: F3: Lactobacillus, C8: Lactococcus | |
In the present investigation, there were no abnormal signs in both the control and treated mice during the 16 day period of experiments. Although F3-treated mice consumed more milk (100 mL daily), yet there were no significant differences (p>0.05) in consumption of liquids between untreated (water or blank milk) and treated (F3 or C8 probiotic milks) mice. This finding completely agreed with data reported by Shu et al. (1999).
As depicted in Table 7, F3 and C8 possessed antagonistic activities against some of GI microflora of mice. In fact, there are several reports showing that specific probiotic strains protect against gastrointestinal infections (Millette et al., 2008; Vesterlund et al., 2006). The safety of different probiotic LAB, such as Lactobacillus acidophilus and Bifidobacterium was studied in mice fed with doses of test strains.
Although the safety status of traditional LAB strains used in food products has been defined by extensive studies, it is nevertheless important to confirm the safety of any newly identified probiotic strains (Shu et al., 1999). Based on the above reports, the safety and efficacy of some isolated LAB from test food (camel’s milk) were carried out using mice feeding trails.
In the present study, there was no significant difference (p>0.05) in weight gain among control and treated mice. The average gain was 1.2 g during the trail periods of 16 days. These findings were in complete agreement with a previous investigation by Vesterlund et al. (2006) regarding detachment of pathogens to GI, F3 and C8 prevented adhesion of staphylococci in the intestine of mice. The counts of staphylococci in feces of mice fed test isolates were significantly (p<0.05) higher comparing to the control or blank milk (Table 7). Additionally, probiotic isolates antagonized growth of E. coli (Table 7) since the counts of those enterics were less in feces of mice fed test isolates (F3 or C8). The commercial probiotic yoghurt used in experiments contained Bifidobacterium acitiregularis (as indicated in label) had lesser activity, compared to control or blank, against E. coli (Table 7).
Probiotic-containing foods can be categorized as functional foods and are often associated with prebiotics which are non-digestible carbohydrates that act as food for probiotics. Functional foods contain beneficial properties over and above their normal nutritional value. In this framework, probiotics are actually being functional products. Probiotics are obtained by the action of microorganisms, usually LAB and yeasts. Those microorganisms are useful in assisting the gastrointestinal tract by breaking down sugars and carbohydrates to promote good digestion, boost the immune system and maintain proper intestinal pH. When probiotics and prebiotics are combined, they form a synbiotic. Yoghurt is considered a synbiotic food because it contains live bacteria (Giraffa, 2011). When choosing a probiotic strain, it must meet different requisites, including product safety for human and animal consumption (GRAS) and survival in the Gastrointestinal (GI) tract. Probiotics are chosen from the European list of accepted additives; the reference law for the permission to introduce and employ additives in feedstuff or pre-mixtures for feedstuff is the EU Regulation 1831/2003. For this reason, the development of new probiotic products that could be licensed for animal use is receiving considerable interest (Ripamonti et al., 2011).
CONCLUSION
The present study showed varying levels of probiotics in the camel’s milk. The CFS were sensitive to proteolytic enzymes indicating the proteinaceous nature (i.e., bacteriocins) of inhibitors. There was no significant difference (p>0.05) in weight gain among control and treated mice. The counts (log10 CFU mL-1) of test isolates were not significantly (p>0.05) affected by bile slats in MRS broth. The LAB isolates exhibited probiotic activities in albino mice by preventing the adhesion of Staphylococcus aureus and inhibiting E. coli in GI.
ACKNOWLEDGMENTS
The authors thank the King Abdul-Aziz City for Science and Technology (KACST) for the partial support of this research project No. ARP-29-141.
REFERENCES
- Adnan, A.F.M. and I.K. Tan, 2007. Isolation of lactic acid bacteria from Malaysian foods and assessment of the isolates for industrial potential. Bioresour. Technol., 98: 1380-1385.
CrossRefPubMedDirect Link - Alegria, A., S. Delgado, C. Roces, B. Lopez and B. Mayo, 2010. Bacteriocins produced by wild Lactococcus lactis strains isolated from traditional, starter-free cheeses made of raw milk. Int. J. Food Microbiol., 143: 61-66.
CrossRef - Cheikhyoussef, A., N. Pogori, W. Chen and H. Zhang, 2008. Antimicrobial proteinaceous compounds obtained from bifidobacteria: From production to their application. Int. J. Food Microbiol., 125: 215-222.
CrossRefPubMedDirect Link - Chen, H. and D.G. Hoover, 2003. Bacteriocins and their food applications. Comprehen. Rev. Food Sci. Food Saf., 2: 82-100.
CrossRefDirect Link - De Vuyst, L. and F. Leroy, 2007. Bacteriocins from lactic acid bacteria: Production, purification and food applications. J. Mol. Microbiol. Biotechnol., 13: 194-199.
CrossRefDirect Link - Giraffa, G., 2011. Probiotics, health claims and consumer needs: Do they always overlap? Ferment. Technol., Vol. 1.
CrossRefDirect Link - Jans, C., J. Bugnard, P.M.K. Njage, C. Lacroix and L. Meile, 2012. Lactic acid bacteria diversity of African raw and fermented camel milk products reveals a highly competitive, potentially health-threatening predominant microflora. LWT Food Sci. Technol., 47: 371-379.
CrossRefDirect Link - Jiang, J., B. Shi, D.Q. Zhu, Q.X. Cai and Y.R. Chen et al., 2012. Characterization of a novel bacteriocin produced by Lactobacillus sakei LSJ618 isolated from traditional Chinese fermented radish. Food Control, 23: 338-344.
CrossRefDirect Link - Liu, S.N., Y. Han and Z.J. Zhou, 2011. Lactic acid bacteria in traditional fermented Chinese foods. Food Res. Int., 44: 643-651.
CrossRef - Malheiros, P.S., V. Sant'Anna, M. Utpott and A. Brandelli, 2012. Antilisterial activity and stability of nanovesicle-encapsulated antimicrobial peptide P34 in milk. Food Control, 23: 42-47.
CrossRefDirect Link - Medici, M., C.G. Vinderola, R. Weill and G. Perdigon. 2005. Effect of fermented milk containing probiotic bacteria in the prevention of an enteroinvasive Escherichia coli infection in mice. J. Dairy Res., 72: 243-249.
PubMedDirect Link - Millette, M., G. Cornut, C. Dupont, F. Shareck, D. Archambault and M. Lacroix, 2008. Capacity of human nisin and pediocin-producing lactic acid bacteria to reduce intestinal colonization by vancomycin-resistant enterococci. Applied Environ. Microbiol., 4: 1997-2003.
CrossRef - Morandi, S. and M. Brasca, 2012. Safety aspects, genetic diversity and technological characterisation of wild-type Streptococcus thermophilus strains isolated from north Italian traditional cheeses. Food Control, 23: 203-209.
CrossRef - Nikolic, M., A. Terzic-Vidojevic, B. Jovcic, J. Begovic, N. Golic and L. Topisirovic, 2008. Characterization of lactic acid bacteria isolated from Bukuljac, a homemade goat's milk cheese. Int. J. Food Microbiol., 122: 162-170.
CrossRef - Ravishankar, S., L. Zhu, C.W. Olsen, T.H. McHugh and M. Friedman, 2009. Edible apple film wraps containing plant antimicrobials inactivate foodborne pathogens on meat and poultry products. J. Food Sci., 74: M440-M445.
CrossRefPubMedDirect Link - Ricci, G., F. Borgo, C. Ferrario and M. Fortina, 2011. Cocoa powder as delivery medium for probiotic Lactobacillus strains. Adv. Microbiol., 1: 1-6.
CrossRef - Ripamonti, B., A. Agazzi, C. Bersani, P. de Dea and C. Pecorini et al., 2011. Screening of species-specific lactic acid bacteria for veal calves multi-strain probiotic adjuncts. Anaerobe, 17: 97-105.
CrossRef - Settanni, L. and A. Corsetti, 2008. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol., 121: 123-138.
CrossRefDirect Link - Shu, Q., S.J. Zhou, K.J. Rutherfurd, M.J. Birtles, J. Prasad, P.K. Gopal and H.S. Gill, 1999. Probiotic lactic acid bacteria (Lactobacillus acidophilus HN017, Lactobacillus rhamnosus HN001 and Bifidobacterium lactis HN019) have no adverse effects on the health of mice. Int. Dairy J., 9: 831-836.
CrossRef - Vesterlund, S., M. Karp, S. Salminen and A.C. Ouwehand, 2006. Staphylococcus aureus adheres to human intestinal mucus but can be displaced by certain lactic acid bacteria. Microbiology, 152: 1819-1826.
CrossRef - Yateem, A., M.T. Balba, T. Al-Surrayai, B. Al-Mutairi and R. Al-Daher, 2008. Isolation of lactic acid bacteria with probiotic potential from camel milk. Int. J. Dairy Sci., 3: 194-199.
CrossRefDirect Link