
Jahwarhar Izuan Abd Rashid
Department of Chemistry and Biology, Centre for Defense Foundation Studies, National Defense University of Malaysia, Sungai Besi Camp, 57000, Kuala Lumpur, Malaysia
Noraini Samat
Strategic Livestock Research Centre, Malaysian Agriculture Research and Development Institute, P.O. Box 12301, 50774 Kuala Lumpur, Malaysia
Wan Mohtar Wan Yusoff
School of Bioscience and Biotechnology, Faculty of Science and Technology, National University of Malaysia, 43600, Bangi, Malaysia
Research Journal of Microbiology
Year: 2012 | Volume: 7 | Issue: 5 | Page No.: 242-255







ABSTRACT
In this study, two statistical experimental methods including Plackett Burman design and Response Surface Methodology (RSM) were employed to optimize the medium composition for mannanase production by Aspergillus terreus SUK-1, a locally isolated fungus using Palm Kernel Cake (PKC) as the sole carbon source. A total of 19 media were screened in order to determine the optimum media for mannanase production using Plackett-Burman design. Soya bean, proteose peptone, urea and NH4NO3 were identified as the supplement to give positive effect on mannanase production by A. terreus SUK-1. Proteose peptone was more significant in terms of its effect on mannanase production compared to urea. The optimum of the medium composition was predicted when the concentrations of proteose peptone and urea were set at 0.5 and 1% (g g-1), respectively. The optimum mannanase activity was closed to the predicted value 30.24±1.32-32.60 IU g-1 when the optimum medium composition was employed. Under the optimized medium (0.5% (g g-1) of proteose peptone and 1% (g g-1) of urea), Mannanase production was 3-fold higher than the activity obtained under original medium (9.97±1.34 IU g-1).
PDF Abstract XML References Citation
Received: January 10, 2012;
Accepted: July 10, 2012;
Published: August 31, 2012
How to cite this article
Jahwarhar Izuan Abd Rashid, Noraini Samat and Wan Mohtar Wan Yusoff, 2012. Screening and Optimization of Medium Composition for Mannanase Production by Aspergillus terreus SUK-1 in Solid State Fermentation Using Statistical Experimental Methods. Research Journal of Microbiology, 7: 242-255.
URL: https://scialert.net/abstract/?doi=jm.2012.242.255
URL: https://scialert.net/abstract/?doi=jm.2012.242.255
INTRODUCTION
Hemicellulose are complete polysaccharides consisting of linear and branched chain in the cell walls of higher plants which is closely associated to the cellulose and lignin forming lignocelluloses biomass (Moreira and Filho, 2008). Mannan is the most abundant of polysaccharide present in softwood hemicellulose. Manno-oligosaccharides are generated when mannan is hydrolysed by the combination of β-mannanase (EC 3.2.1.78), β-mannosidase (EC 3.2.1.25) and β-glucosidase (EC 3.2.1.21) with debranching enzymes such as galactosidase (EC 3.2.1.22) and acetyl esterase (EC 3.1.1.6) (Gubitz et al., 1996; Singh et al., 2003; Petkowicz et al., 2007).
Mannanase enzyme is important in paper industry including bioleaching pulp (Gubitz et al., 1996) waste bioconversion of biomass to fermentable sugars (Chandrakant and Bisaria, 1998), increasing the quality of feed quality (Marini et al., 2006) and reduce viscosity of coffee extracts (Hagglund et al., 2003).
Filamentous fungi are commonly used for enzyme production in SSF since have the ability to secrete large amount of protein into the growth medium (Van Zyl et al., 2009). Aspergillus species have been known as potential fungi in the production of a wide range of microbial enzymes (Gao et al., 2008). Therefore, in this study we used our local strain, Aspergillus terreus SUK-1 which was isolated from palm oil mill sludge and capable to enhance mannanase production (Rashid et al., 2011a, b).
Solid State Fermentation (SSF) defined as any fermentation process performed in the absence or near-absence of free water, employing a natural substrate or an inert support (Sabu et al., 2006). As it was reported that SSF technique is yet to be explored for mannanase production (Noraini et al., 2004; Ong et al., 2004; Regulapati et al., 2007; Abd-Aziz et al., 2008; Norita et al., 2010) despite its many advantages such as lower capital investment, improved product recovery and easy downstream processing etc. over the conventional submerged process (Mitchell and Lonsane, 1992; Pandey et al., 2001; Naveena et al., 2003). Improvement in productivity of mannanase by A. terreus SUK-1 is done by manipulating the nutritional and physical parameters in solid state fermentation.
Therefore, development of an economical production medium requires selection of carbon, nitrogen, phosphorus, potassium and trace element sources. Medium optimization by single dimensional search is laborious and time consuming, especially for a large number of variables and it does not ensure desirable conditions. Plackett-Burman design is widely used in screening experiment as the number of experiment run required are very few, leading to saving of time, chemicals, glass ware and man power (Srinivas et al., 1994; Naveena et al., 2003; Sabra and Hassan, 2008; Jamal et al., 2009). Hence optimization of fermentation media is done at two step level. According to Pareto’s law, initial screening of the ingredients is done to understand the significance of their effect on the product formation and then a few better ingredients are selected for further optimizations (Ganapathy et al., 1998; Naveena et al., 2003). RSM which has been extensively applied in optimization of medium composition, conditions of enzymatic hydrolysis, fermentation and food manufacturing process as well as process parameters for enhanced production (Wang and Lu, 2005; Jaiswal et al., 2011).
No data on mannanase production by A. terreus SUK-1 in SSF for optimization of medium nutrients has been published yet. However, there was a study of medium optimization for endoglucanase production by A. terreus SUK-1 using these statistical models (Youssef and Berekaa, 2009). In the present study, we report screening of medium to supplement with PKC as sole carbon sources using Plackett-Burman design for mannanase production in SSF by A. terreus SUK-1. The critical medium nutritionals from screening experiment were further optimized the medium composition using response surface methodology.
MATERIALS AND METHODS
Microorganism: A local isolate Aspergillus terreus SUK-1 from School of Biosciences and Biotechnology, Universiti Kebangsaan Malaysia, was grown and maintained on Potato Dextrose Agar (PDA). Spore suspension of 107 spore mL was prepared by harvesting from 7-days old cultures mold with 15 mL sterile distilled water and 5.5% (w/v) of inoculum was used in all experiments.
Substrate and medium: Palm kernel cake was supplied by Malaysian Agriculture Research Development Institute (MARDI) and used as a solid substrate and PKC was ground to a particle size of 2 mm. The medium employed (Sternberg, 1976) is shown in Table 1.
| Table 1: | SSF medium for mannanase production by A. terreus SUK-1 |
 | |
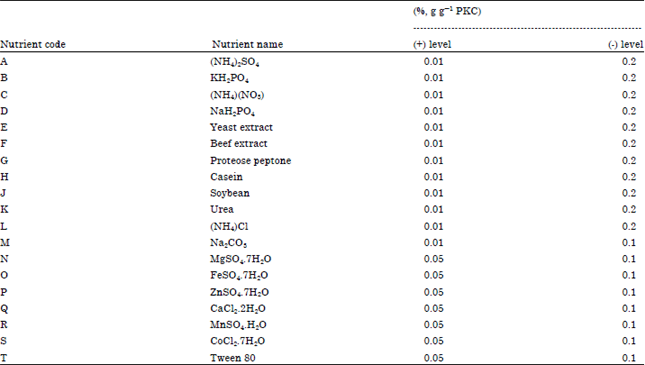
| Table 2: | Nutrients supplements for screening using Plackett-Burman design |
 | |
Solid state fermentation: Fermentation was carried out in Erlenmeyer flask 250 mL containing 25 g of (PKC) and Sternberg’s medium (Sternberg, 1976) with ratio initial moisture 1:0.75 (w/v). The mixtures were autoclaved at 121°C for 15 min. Each flask was inoculated with 5.5% (w/v) inoculum size of spore suspension of A. terreus SUK-1. All flasks were incubated for 4 days under 30°C. Duplicate flasks were set up under various experiments according to the experimental design as described in the following paragraphs.
Experimental design
Plackett-Burman design: Plackett-Burman factorial design was employed for screening the important nutrients for mannanase production by A. terreus SUK-1. Nineteen medium nutrients (Table 2) were used to determine the key ingredients significantly affecting the mannanase production. Based on Plackett-Burman factorial design, each factor was examined at two levels: -1 for low level and +1 for high level and a center point was run to evaluate the linear and curvature effects of the variables. Table 2 shows the Plackett-Burman experimental design with 19 factors under investigation as well as levels of each factor used in the experimental design and the response. Plackett–Burman experimental design is based on the first-order polynomial model:
Y = B0+ΣB1X1
where, Y is the response (enzyme activity), B0 is the model intercept and B1 is the linear coefficient and X1 is the level of the independent variable. This model does not describe interaction among factors and it is used to screen and evaluate the important factors that influence the response. In the present work, 19 assigned variables were screened in 30 experimental runs in addition to the three runs at their center points. Enzyme activity assays were carried out in duplicate and the averages of the mannanase activity was taken as response Y. From the regression analysis of the variables, the factors significant at 90% level (p<0.1) were considered to have significant effect on mannanase production and were further optimized by RSM.
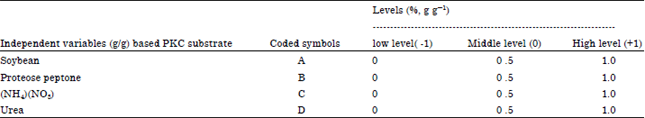
Optimization of critical medium nutrients using RSM: Based on the results of screening of medium nutrient from Plackett-Burman design, the medium nutrients that significantly had positive affect on mannanase production were identified and optimized further using the RSM). Each variable in the design was studied at three different levels (Table 3).
| Table 3: | Nutrients for the production of mannanase by Aspergillus terreus SUK 1 using Plackett Burman design criterion |
 | |
| A: (NH4)2SO4, B: KH2PO4, C: (NH4)(NO3), D: NaH2PO4, E: Yeast Extract, F: Beef Extract, G: Proteose peptone, H: Casein, J: Soybean, K: Urea, L: (NH4)Cl, M: Na2CO3, N: MgSO4.7H2O, O: FeSO4.7H2O, P: ZnSO4.7H2O, Q: CaCl2.2H2O, R: MnSO4.H2O, S: CoCl2.7H2O, T: Tween 80 | |
A 24 factorial design, with eight axial points and six replicates at the central points with a total number of 30 experiments were employed according to statistical package, software design expert 6.0 (StatEase Inc. Minneapolis, USA) to fit a second order polynomial response surface methodology according to the following equation:
Y = X0+X1A+X2B+X3C+X4D+X11A2X22B2X33C2+X44D2+X1X2AB+X1X3AC+X1X4AD+X2X3BC+X2X4BD+X3X4CD
Y is the mannanase activity (U g-1), X0, X1,......,X23 represent the estimated regression coefficients, X1, X2, X3, X4 represent the linear effect, X11, X22, X33, X44, the quadratic effect and X1X2, X1X3,X1X4, X2X3, X2X4, X3X4, cross product coefficient A, B, C, D represent variables of selected nutrient.
Crude enzyme extraction: Mannanase enzyme was extracted by adding 100 mL of distilled water into the flask containing 10 g of fermented PKC and agitated at 150 rpm for 24 h at 10°C. Solids were removed by filtration using Whatman filter paper No.1 followed by centrifugation. The supernatant was used as the source of mannanase enzyme.
Enzyme assay: Mannanase activity was carried out according to the method described by McCleary (1978) using azo carob galactomannan as substrate. One unit (U) of mannanase activity was defines as mannose released/min/g of substrate.
RESULTS AND DISCUSSION
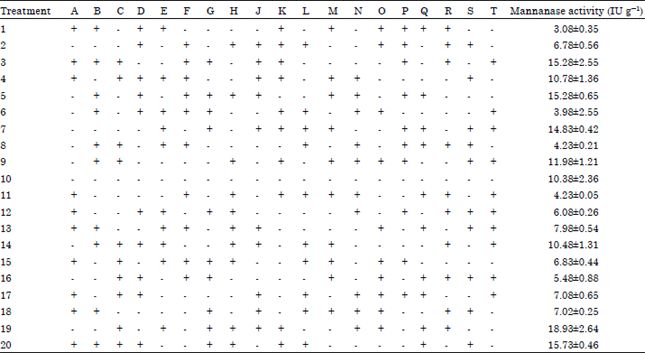
Screening of medium nutrient by Plackett-Burman: Plackett-Burman factorial design was employed for screening the important nutrients to supplement with PKC in solid state fermentation (SSF) for mannanase production. Table 4 shows that treatment 19 with supplement of medium nutrient of C, E, G, H, I, J, M, N, P and Q gave a better value of mannanase production (18.93±2.64 IU g-1) than the others. Out of 19 variables, 11 namely (NH4)2SO4 (NH4)(NO3), NaH2PO4, beef extract, proteose peptone, casein, soybean, urea (NH4)Cl, FeSO4.7H2O and MnSO4.H2O influenced mannanase production significantly (Fig. 1).
| Table 4: | Variables and experimental design levels for response surface methodology |
 | |
 | |
| Fig. 1: | The main effect of the fermentation nutrient on mannanase production according to Plackett-Burman design results. A: (NH4)2SO4, C: (NH4)(NO3), D: NaH2PO4, F: Beef extract, G: Proteose peptone, H: Casein, J: Soybean, K: Urea, L: (NH4)Cl, O: FeSO4.7H2O, R: MnSO4.H2O |
The addition of mineral salts and trace elements (KH2PO4, ZnSO4.7H2O, Na2CO3, MgSO4.7H2O, CaCl2.2H2O and CoCl2.7H2O) into the PKC substrate had no significant effect on mannanase production by A. terreus SUK-1. This result may be explained by the fact that mineral salts and trace elements such as zinc, copper, calcium, magnesium and phosphorus are present in the PKC substrate (Marini et al., 2006) and thus support the growth of Aspergillus as well as to induce mannanase production. The size of each coefficient gives a direct measurement of the importance of each effect (Psomas et al., 2007). Figure 1 clearly shows that supplementation of soybean meal into the PKC substrate was the most influential in affecting the mannanase production followed by proteose peptone, FeSO4.7H2O and NH4NO3. Haaland (1989) reported that the positive sign of the coefficient explaining the positive impact that the production of reaction would increase if the concentration factor increased and vice versa. It was postulated that the positive effects of soybean, proteose peptone (NH4)(NO3), casein and urea could be attributed to the requirement for nitrogen sources during fermentation (Fig. 1). In contrast, negative effects on mannanase production was observed when the level (NH4)2SO4, NaH2PO4, extract beef, casein (NH4)Cl, FeSO4.7H2O and MnSO4.H2O increased.
Earlier studies reported that the addition of inorganic nitrogen sources such as (NH4)Cl (NH4)2SO4 and NaH2PO4 reduced the rate of growth of Aspergillus and thus inhibited the production of enzymes (Jin et al., 1998; Sivaramakrishnan et al., 2007; Mohamad et al., 2011). It was similar results in this study that the supplementation of (NH4)Cl (NH4)2SO4 and NaH2PO4 reduced the synthesis of mannanase enzyme.
According to Marini et al. (2006), PKC substrate contained higher Iron and manganese mineral compared to other minerals (Zinc, Copper). Thus it was postulated that the addition of MnSO4.H2O and FeSO4.7H2O into PKC substrate inhibited mannanase activity. The results of this study are in consistent with the statement that the presence of excess concentration of minerals in medium lead to the toxicity and inhibition of fungal growth and sporulation as well as synthesis of enzymes (Jaworska and Dluzniewska, 2007).
Our findings demonstrated that the supplementation of organic nitrogen sources to PKC substrate was sufficient to enhance the production of mannanase enzyme by A. terreus SUK-1 (Fig. 1). This argument is strengthened by the previous report that the addition of organic nitrogen sources capable of providing stability in the substrate in terms of the pH range of 5.5-6.0 (Lin and Chen, 2004; Noraini et al., 2004). This pH ranges have been report previously as the optimal pH for mannanase production (Arisan-Atac et al., 1993; Ademark et al., 1998; Mudau and Setati, 2008). Nevertheless, when the concentration of nitrogen level is increased, the mannanase activity would decline due to the presence of high concentration of nitrite of fungal cells. High concentration of nitrite in the fungal cell leads to fungal toxicity, thus reducing the production of fungal enzymes mannanase (Raimbault, 1998). From the screening results, we can conclude that the growth of A. terreus SUK-1 in SSF requires the optimum medium of nitrogen sources for the production of enzymes as it was reported that nitrogen sources is a major limiting factor (Tigue et al., 1994; Pedersen and Nielsen, 2000). The medium nutrient that resulted in a negative effect on mannanase production was no longer taken into account, while the positive factors were included in the next optimization process.
The optimization of medium nutrients for mannanase production using RSM method: Soybean, proteose peptone (NH4)NO3 and urea had the most significant positive effect on mannanase production. These supplements were further optimized by RSM using Central Composite Design (CCD).
| Table 5: | Experimental design and results of CCD of response surface methodology |
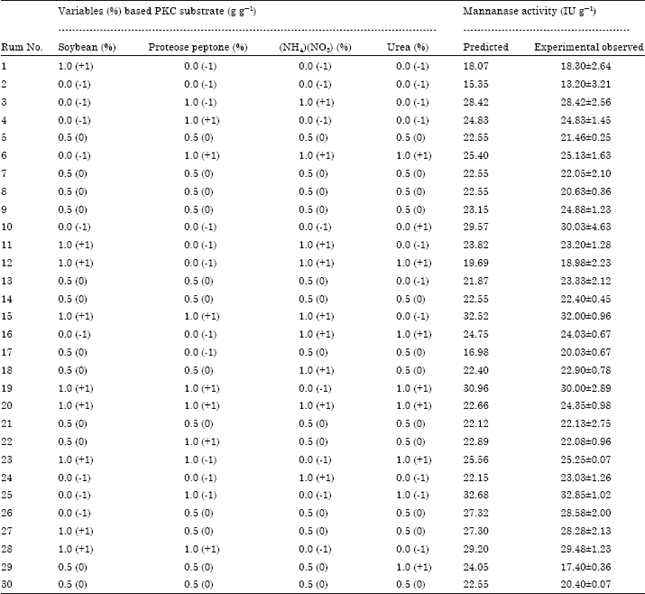 | |
As indicated in Table 5, treatment 25 gave the maximal mannanase enzyme activity (32.85±0.2 IU g-1) when 1% proteose peptone (g g-1 PKC) and 1% urea (g g-1 PKC) substrate were added into PKC substrate. However, the minimum of mannanse enzyme activity (13.20±3.21 IU g-1) was obtained in treatment 2 when there was no addition of nitrogen source in PKC substrate. The responses of the CCD design were fitted with a second-order polynomial equation (Eq. 1). In second-order polynomial equation derived for the optimization of medium components, the mannanase activity (Y) was shown as a function of these variables. By applying multiple regression analysis on the experimental data, the following equation was found to explain the production of mannanase:
Y = 22.56-0.013 A+2.95 B-0.37 C+1.09 D+4.76 A2-2.62 B2+0.22 C2+0.41 D2+0.57 AB-0.26 AC-1.68 AD-0.61 BC-1.43 BD-2.90 CD | (1) |
where, Y represents the response of mannanase production and A, B, C, D, A2, B2, C2, D2, AB, AC, AD, BC, BD and CD are the variables. A, B, C, D represent the coded levels of soybean, proteose peptone, ammonium nitrate (NH4)NO3 and urea, respectively.
| Table 6: | Analysis of variance (ANOVA) for regression model to optimize mannanase production |
 | |
| R2: 0.9234, CV: 6.90, Adjusted R2: 0.8518 | |
| Table 7: | Model coefficient estimated by multiplies linear regression |
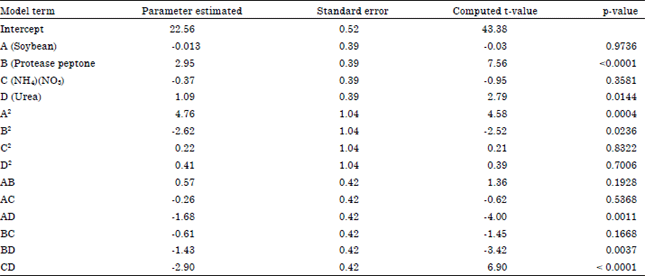 | |
The statistical significance of the regression model was checked by F-test and the analysis of variance for the response surface quadratic model is shown in Table 6. The model was highly significant, as manifested by the F-value and the probability value [(p total model>F) = 0.0007]. The data from the analysis of variance showed that the second-order model was well fitted to the experimental data. The goodness of fit was manifested by the determination coefficient (R2). The Coefficient of determination, R2 gives the information how much of the variability in the observed data could be explained by the experiment and their interaction (Kawaguti et al., 2006). In this case, the R2 value of 0.9234 indicated that the response model can explain 92.3% of the total variations in the model could be explained and about 7.7% of the total variation cannot be explained by the model. In general, a regression model having an R2 value higher than 0.9 is considered to have a very high correlation (Haaland, 1989). The value of the adjusted determination coefficient (R2 adj. = 85.18%) was also high enough to indicate the significance of the model.
The Student’s t-distribution and the corresponding p-value, along with the parameter, are given in Table 7. The corresponding p-value suggested that the linear coefficient (B, C, D), quadratic terms coefficient (A2, B2) and cross product coefficient (AD, BD, CD) whereas the other terms coefficient (A, C2, D2, AB, AC, BC) were not significant(p>0.05).
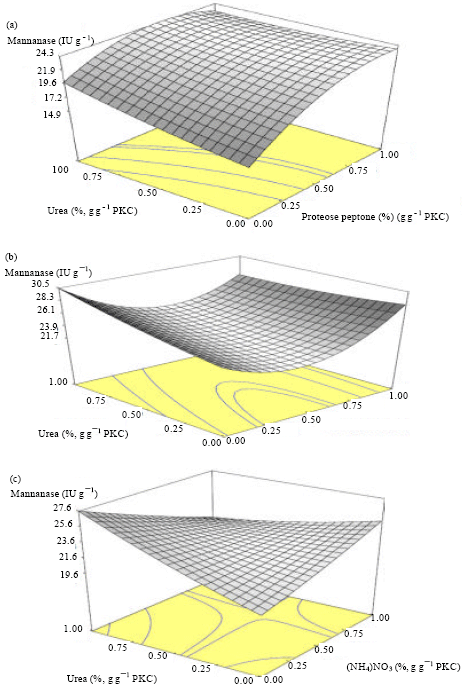 | |
| Fig. 2(a-c): | A. Terreus SUK-1 mannanase production effected by (a) Urea+proteose peptone, (b) Urea+soybean and (c) Urea+NH4NO3 |
Table 7 clearly demonstrated that the supplementation of proteose peptone (B) with PKC substrate was the most influential nutrients on mannanase production.
The 3D response surfaces plots were employed to determine the interaction of the basal medium components and the optimum levels that have the most significant effect on mannanase production. Figure 2 illustrates the relationship between the response and the experimental data. Figure 2a describes the effects of urea and soybean on mannanase production, when proteose peptone and ammonium nitrate were fixed at its middle level (0.5% (g g-1) base substrate PKC, respectively. From Fig. 2a, it can be seen that the mannanase activity increased gradually when urea concentration increased at a high level while soybean concentration decreased at a low level. Out of range 0.5-0.6% concentration of soybean shows that the yield of mannanase production increased gradually.
Figure 2b demonstrates the effects of urea and proteose peptone on mannanase production when the soybean and ammonium nitrate were fixed at its middle level (0.5% (g-1) base substrate PKC, respectively. The mannanase production was predominantly affected by the Proteose Peptone concentration and the mannanase production increased gradually when both of Proteose Peptone concentration and urea concentrations were increased at high level.
Figure 2c shows the effects of urea and ammonium nitrate on the mannanase production when the concentration of soybean and proteose peptone was set at the middle level (0.5% (g-1) base substrate PKC, respectively. It can be seen that the mannanase production could be increased when the Urea concentration increased at a high level while ammonium concentration was fitted a low level.
The maximum activity of mannanase predicted from the model RSM was 32.60 IU g-1 under the optimal media when the concentration of Proteose Peptone and Urea were set at 0.5% (g g-1) and 1% (g g-1), respectively. Verification of the predicted values was conducted by using this value of optimum medium fermentation. The practical corresponding response was 30.24±1.32 IU g-1 which corroborated the validity and the effectiveness of the current model.
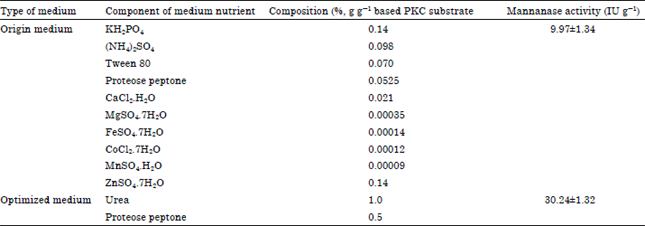
Effect of nitrogen sources on mannanase production: Mannanase production by A. terreus SUK-1 using optimum medium was 3-fold higher (30.24±1.32 IU g-1) than the activity obtained under original medium (9.97±1.34 IU g-1) (Table 8). This is probably because the quantity of original medium components is insufficient to induce the production of mannanase enzyme and also the presence of inhibitor as described in the screening result. So far, this optimum medium consist of organic nitrogen source (urea and proteose peptone) for the production of mannanase enzymes by A. terreus SUK-1 has not been reported yet.
This finding demonstrates that Aspergillus mannanase induced by PKC was significantly enhanced using organic nitrogen medium compared to that induced using inorganic nitrogen medium sources. The present finding is similar to those reported for other fungal mannanase (Lin and Chen, 2004; Rajoka et al., 2006).
| Table 8: | Comparison between origin medium and medium optimized on mannanase production by A. terreus SUK-1 |
 | |
According to Mohamad et al. (2011), organic nitrogen sources not only have the amino acid content but also have a good source of vitamins and carbohydrates that can stimulate the production of enzymes.
Moreover, the addition of Urea (1% g g-1) using PKC as a sole carbon sources enhanced mannanase enzyme by A. niger (Noraini et al., 2004; Ong et al., 2004). Combination of Yeast Extract and NH4NO3 media have been identified as optimum medium for the production of mannanase by A. niger and A. flavus using copra meal as a sole carbon (Kote et al., 2009). Peptone and NH4NO3 have been reported as the best nitrogen source for the production of mannanase enzymes by Sclerotium rolfsii (Gubitz et al., 1996; Grobwindhager et al., 1999; Sachlehner and Haltrich, 1999). In contrast, a few reports have shown that the supplementation of inorganic nitrogen sources into cultivation medium leads to high level of mannanase production (Rashid et al., 2011a). Youssef et al. (2006) concluded that the addition of 2.5 g L-1 (NH4)Cl is the best medium to stimulate the production of enzymes mannanase by A. niger. For this study, proteose peptone and urea are used as the optimum medium for mannanase production by A. terreus SUK-1 using PKC as a substrate for the next experiment.
CONCLUSION
Using Plackett-Burman design, soya bean, proteose peptone, Urea and NH4NO3 were identified as critical factor on mannanase production. Using RSM methods, the high yield of mannanase production can be predicted when the concentration of Proteose Peptone and Urea were set at 0.5 and 1.0% (g g-1), respectively. The optimal mannanase activities were close to the predicted value, 30.24±1.32-32.60 IU g-1 when the optimum medium composition was employed. Under the optimized medium (0.5% (g g-1) of Proteose Peptone and 1% (g g-1) of Urea), Mannanase production was 3-fold higher than the activity obtained under original medium (9.97±1.34 IU g-1).
ACKNOWLEDGMENTS
The authors wish to thanks the Ministry of Science Technology and Innovation (MOSTI), Malaysia for providing financial supports (01-03-03-003 BTK/ER/008). The authors are also grateful to Strategic Livestock Research Centre, MARDI in providing the equipment to carry out this work.
REFERENCES
- Abd-Aziz, S., G.S. Hung, M.A. Hassan, M.I.A. Karim and N. Samat, 2008. Indirect method for quantification of cell biomass during solid-state fermentation of palm kernel cake based on protein content. Asian J. Sci. Res., 1: 385-393.
CrossRefDirect Link - Ademark, P., A. Varga, J. Medve, V. Harjunpaa, T. Drakenberg, F. Tjerneld and H. Stalbrand, 1998. Softwood hemicellulose-degrading enzymes from Aspergillus niger: Purification and properties of β-mannanase. J. Biotechnol., 63: 199-210.
PubMed - Chandrakant, P. and V.S. Bisaria, 1998. Simultaneous bioconversion of cellulose and hemicellulose to ethanol. Crit. Rev. Biotechnol., 18: 295-331.
PubMed - Youssef, G.A. and M.M. Berekaa, 2009. Improved production of endoglucanase enzyme by Aspergillus terreus: Application of plackett burman design for optimization of process parameters. Biotechnology, 8: 212-219.
CrossRefDirect Link - Gao, J., H. Weng, D. Zhu, M. Yuan, F. Guan and Y. Xi, 2008. Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresour. Technol., 99: 7623-7629.
CrossRefDirect Link - Gubitz, G.M., M. Hayn, M. Sommerauer and W. Steiner, 1996. Mannan-degrading enzymes from Sclerotium rolfsii: Characterisation and synergism of two endo β-mannanases and a β-mannosidase. Bioresource Technol., 58: 127-135.
CrossRef - Grobwindhager, C., A. Sachslehner, B. Nidetzky and D. Haltrich, 1999. Endo-β-1,4-d-mannanase is efficiently produced by Sclerotium (Athelia) rolfsii under derepressed conditions. J. Biotechnol., 67: 189-203.
CrossRef - Jaworska, M. and J. Dluzniewska, 2007. The effect of manganese ions on development and antagonism of Trichoderma isolates. Polish J. Environ. Stud., 16: 549-553.
Direct Link - Rashid, J.I.A., N. Samat and W.M.W. Yusoff, 2011. Optimization of temperature, moisture content and inoculum size in solid state fermentation to enhance mannanase production by Aspergillus terreus SUK-1 using RSM. Pak. J. Biol. Sci., 14: 533-539.
CrossRefDirect Link - Jamal, P., M.F. Tompang and M.Z. Alam, 2009. Optimization of media composition for the production of bioprotein from pineapple skins by liquid-state bioconversion. J. Applied Sci., 9: 3104-3109.
CrossRefDirect Link - Kawaguti, H.Y., E. Manrich and H.H. Sato, 2006. Application of response surface methodology for glucosyltransferase production and conversion of sucrose into isomaltulose using free Erwinia sp. cells. Electron. J. Biotechnol., 9: 5-9.
Direct Link - Kote, N.V., A.G. Patil and V.H. Mulimani, 2009. Optimization of the production of thermostable endo-β-1,4 mannanases from a newly isolated Aspergillus niger gr and Aspergillus flavus gr. Applied Biochem. Biotechnol., 152: 213-223.
PubMed - McCleary, B.V., 1978. A simple assays procedure for β- mannanase. Carbohydrate Res., 67: 213-221.
PubMed - Mohamad, S.N., R.N. Ramanan, R. Mohamad and A.B. Ariff, 2011. Improved mannan-degrading enzymes production by Aspergillus niger through medium optimization. N. Biotechnol., 28: 146-152.
PubMed - Moreira, L.R. and E.X. Filho, 2008. An overview of mannan structure and mannan-degrading enzyme systems. Applied Microbiol. Biotechnol., 79: 165-178.
CrossRefPubMedDirect Link - Tigue, M.A., C.T. Kelly, W.M. Fogarty and E.M. Doyle, 1994. Production studies on the alkaline amylases of three alkalophilic Bacillus spp. Biotechnol. Lett., 16: 569-574.
CrossRef - Jaiswal, N., O. Prakash, M. Talat, S.H. Hasan and R.K. Pandey, 2011. Application of response surface methodology for the determination of optimum reaction conditions (Temperature and pH) for starch hydrolysis by α-amylase. Asian J. Biochem., 6: 357-365.
CrossRef - Noraini, S., S. Vikineswary, M.J. Daud, A. Ibrahin, A. Aman and P. Sevagam, 2004. Biosynthesis of β-mannanase from Aspergillus niger by solid substrate fermentation (SSF) using palm kernel cake as the basis of solid substrate. Proceedings of the International Seminar on Solid State Fermentation, October 4-6, 2004, Penang, Malaysia, pp: 175-176.
- Norita, S.M., M. Rosfarizan and A.B. Ariff, 2010. Evaluation of the activities of concentrated crude mannan-degrading enzymes produced by Aspergillus niger. Malaysian J. Microbiol., 6: 171-180.
Direct Link - Ong, L.G.A., S. Abd-Aziz, S. Noraini, M.I.A. Karim and M.A. Hassan, 2004. Enzyme production and profile by Aspergillus niger during solid substrate fermentation using palm kernel cake as substrate. Applied Biochem. Biotechnol., 118: 73-79.
Direct Link - Pandey, A., C.R. Soccol, J.A. Rodriguez-Leon and P. Nigam, 2001. General Considerations about Solid-State Fermentation Processes. In: Solid State Fermentation in Biotechnology: Fundamentals and Applications, Pandey, A., C.R. Soccol, J.A. Rodriguez-Leon and P. Nigam (Eds.). Asiatech Publishers, New Delhi, pp: 11-18.
- Pedersen, H. and J. Nielsen, 2000. The influence of nitrogen sources on the α-amylase productivity of Aspergillus oryzae in continuous cultures. Applied Microbiol. Biotechnol., 53: 278-281.
PubMed - Van Zyl, P.J., V. Moodley, S.H. Rose, R.L. Roth and W.H. van Zyl, 2009. Production of the Aspergillus aculeatus endo-1,4-β-mannanase in A. niger. J. Ind. Microbiol. Biotechnol., 36: 611-617.
CrossRef - Psomas, S.K., M. Liakopoulou-Kyriakides and D.A. Kyriakidis, 2007. Optimization study of xanthan gum production using response surface methodology. Biochem. Eng. J., 35: 273-280.
CrossRefDirect Link - Rajoka, M.I., M.W. Akhtar, A. Hanif and A.M. Khalid, 2006. Production and characterization of a highly active cellobiase from Aspergillus niger grown in solid state fermentation. World J. Microbiol. Biotechnol., 22: 991-998.
CrossRef - Rashid, S.A., I. Darah and I.C. Omar, 2011. Utilization of palm kernel cake for the production of mannanase by an indigenous filamentous fungus, Aspergillus niger USM F4 under solid substrate fermentation. Internet J. Microbiol., Vol. 9, No. 1.
Direct Link - Regulapati, R., N.P. Malav and N.S. Gummadi, 2007. Production of Thermostable α-amylases by solid state fermentation: A review. Am. J. Food Technol., 2: 1-11.
CrossRefDirect Link - Sabra, W. and M. Hassan, 2008. Exopolysaccharide yield as a kinetic parameter for the statistical optimization of EPS production by Klebsiella pneumoniae. Biotechnology, 7: 27-34.
CrossRefDirect Link - Sabu, A., C. Augur, C. Swati and A. Pandey, 2006. Tannase production by Lactobacillus sp. ASR-S1 under solid-state fermentation. Process Biochem., 41: 575-580.
CrossRef - Sachlehner, A. and D. Haltrich, 1999. Purification and some properties of a thermostable acidic endo-β-1,4-D-mannanase from Sclerotium (Athelia) rolfsii. FEMS Microbiol. Lett., 177: 47-55.
CrossRefDirect Link - Sternberg, D., 1976. Production of cellulase by Trichoderma. Biotechnol. Bioeng. Symp., 6: 35-53.
PubMed - Sivaramakrishnan, S., D. Gangadharan, K.M. Nampoothiri, C.R. Soccol and A. Pandey, 2007. Alpha amylase production by Aspergillus oryzae employing solid-state fermentation. J. Sci. Ind. Res., 66: 621-626.
Direct Link - Singh, S., A.M. Madlala and B.A. Prior, 2003. Thermomyces lanuginosus: Properties of strains and their hemicellulases. FEMS Microbiol. Rev., 27: 3-16.
CrossRefDirect Link - Srinivas, M.R.S., N. Chand and B.K. Lonsane, 1994. Use of plackett-burman design for rapid screening of several nitrogen sources, growth/product promoters, minerals and enzyme inducers for the production of α-galactosidase by Aspergillus niger MRSS 234 in solid state fermentation system. Bioprocess Biosyst. Eng., 10: 139-144.
CrossRef - Wang, Y.X. and Z.X. Lu, 2005. Optimization of processing parameters for the mycelial growth and extracellular polysaccharide production by Boletus spp. ACCC 50328. Process Biochem., 40: 1043-1051.
Direct Link - Youssef, A.S., M.Y. El-Naggar, S.A. El-Assar and E.A. Beltagy, 2006. Optimization of cultural conditions for β-mannanase production by a local Aspergillus niger isolate. Int. J. Agric. Biol., 8: 539-545.
Direct Link - Raimbault, M., 1998. General and microbiology aspect of solid substrate fermentation. Elect. J. Biotechnol, 1: 26-27.
Direct Link