
K. Siva Kumar
Department of Biotechnology, College of Science and Technology andhra University, Visakhapatnam-530 003, Andhra Pradesh, India
R. Haritha
Department of Biotechnology, College of Science and Technology andhra University, Visakhapatnam-530 003, Andhra Pradesh, India
A. Swathi
Department of Biotechnology, College of Science and Technology andhra University, Visakhapatnam-530 003, Andhra Pradesh, India
B. Sirisha
Department of Biotechnology, College of Science and Technology andhra University, Visakhapatnam-530 003, Andhra Pradesh, India
T. Ramana
Department of Biotechnology, College of Science and Technology andhra University, Visakhapatnam-530 003, Andhra Pradesh, India
Research Journal of Microbiology
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 171-181







ABSTRACT
A new isolate of actinomycete has been isolated from marine sediment sample collected from Bay of Bengal near Visakhapatnam coast, as a result of screening actinomycete isolates producing antimicrobial compounds. The isolate exhibited broad spectrum of antimicrobial activity against the pathogenic bacteria and fungi used in the study. The isolate exhibited bluish green to light blue aerial mycelium with long spore chains which are non-motile spores with spiny surfaces. Growth of the isolate occurred in 2-7% (w/v) NaCl at pH 5.2-10.0 and at 25-42°C. The predominant menaquinones are MK-9 (H6) (81%) and MK-9 (H8) (19%) and the major cellular fatty acids were anteiso-C15:0, iso-C16:0 and iso-C15:0. The DNA G+C content of the strain was 64.5 mol%. Phylogenetic analysis based on 16S rRNA gene sequences showed that the strain belongs to genus Streptomyces, with highest similarity to Streptomyces coeruleorubidus NBRC 12844T (99.5%). Based on the physiological, biochemical and chemotaxonomic tests, the actinomycete was identified as an isolate of Streptomyces coeruleorubidus sp. from marine sediment.
PDF Abstract XML References Citation
Received: December 22, 2011;
Accepted: January 26, 2012;
Published: June 21, 2012
How to cite this article
K. Siva Kumar, R. Haritha, A. Swathi, B. Sirisha and T. Ramana, 2012. Taxonomy and Antimicrobial Activity of Streptomyces coeruleorubidus sp. Isolated from Marine Sediment. Research Journal of Microbiology, 7: 171-181.
URL: https://scialert.net/abstract/?doi=jm.2012.171.181
URL: https://scialert.net/abstract/?doi=jm.2012.171.181
INTRODUCTION
Actinomycetes are gram positive bacteria contain high G+C content in the genetic material, were ascertained as the potential source of secondary metabolites of pharmaceutical importance (Ellaiah et al., 2004) and recycling of metabolites from the degradation of biopolymers such as keratin, lignocellulose and chitin (McCarthy, 1987). Among actinomycetes Streptomyces hold an important position due to their ability to produce several bioactive compounds such as antibiotics, industrially useful enzymes (Narayana and Vijayalakshmi, 2008a, b; Arunachalam et al., 2010) and enzyme inhibitors (Berdy, 1995; Fiedler et al., 2005; Goodfellow, 1988; Imada, 2005; Miao and Davies, 2010). The genus Streptomyces proposed by Waksman and Henrici (1943) forms a major group of microorganisms that are widely distributed in nature. Several actinobacterial strains capable of producing bioactive compounds from diverse habitats have been reported from the past fifty years (Raja and Prabakarana, 2011). Recently, the rate of discovery of new compounds from terrestrial actinomycetes has been decreased. Thus, the unexplored or underexploited marine habitats were selected as rich sources of new bacterial taxa producing novel bioactive metabolites (Lam, 2006).
Oceans cover 70% of the earth surface and the organisms growing in marine environments and mangrove swamps were metabolically and physiologically different from terrestrial organisms (Kannan and Vincent, 2011). The diversity of actinomycetes in the marine environment usually is investigated with culture dependent and/or culture-independent techniques. The approach to culture dependent diversity surveys is based upon the use of sediment pre-treatments, taxon-selective isolation media, preliminary circumscription and dereplication of isolates (Maldonado et al., 2005). Similarly the approach to culture-independent surveying has relied on actinobacterial primers designed on the basis of 16S rRNA sequence databases (Stach et al., 2003). Some strains have been already reported from marine environments (Takizawa et al., 1993; Kumar et al., 2011). However, the full potential of this domain as the basis for biotechnology, particularly in India, remains largely unexplored. In the course of screening for novel antibiotics, one actinomycete strain with potent antibacterial and antifungal activities was isolated from the sediments of Bay of Bengal near Visakhapatnam, India. The isolate was then identified using a polyphasic approach (Khucharoenphaisan et al., 2012). The results showed that the strain belongs to the genus Streptomyces.
MATERIALS AND METHODS
Strain and culture conditions: An actinomycete was isolated from a marine sediment sample collected from Bay of Bengal near Visakhapatnam coast (Latitude 17°51.264 N and Longitude 83°32.060 E) at a depth of 30 m. This strain was isolated by the standard serial dilution technique (Jensen et al., 1991) using glycerol asparagine agar (Shirling and Gottlieb, 1966) and the plates were incubated at 28°C for 4 weeks. The purified strain was picked and maintained on (International Streptomyces Project) ISP 4 slants and deposited in Microbial Type Culture Collection and Gene Bank (MTCC), Institute of Microbial Technology, Chandigarh as strain MTCC 33992T. Biomass for chemical and molecular characterization studies was obtained by cultivation in shake flasks cultured at 120 rpm using soybean meal 1% (w/v), NaCl 1% (w/v), glucose 1% (w/v), CaCO3 0.1% (w/v) at 28°C for 6 days.
Morphological, physiological and biochemical characteristics: The cell morphology of the strain was studied with the cultures grown on yeast-malt extract agar (ISP-2) medium (Shirling and Gottlieb, 1966) at 28°C for 2 weeks by light microscopy (Magnus microscope MLX). For Scanning Electron Microscopy (SEM) the strain was fixed with 1.5% glutaraldehyde and dehydrated with graded series of ethanol washes followed by drying in desiccators (EMITECH-K850-CPD). Samples were fixed to SEM stubs using carbon tape followed by thin coating with platinum and examined with scanning electron microscope (JEOL; JSM-6610LV). Aerial mycelium, spore mass colour, substrate mycelium pigmentation and coloration of diffusible pigments of the strain were recorded on ISP media. The phenotypic properties of the strain were studied using standard procedures (Shirling and Gottlieb, 1966; Williams et al., 1983). Physiological tests such as growth at different temperatures (15, 25, 37, 42, 50°C), pH (5.2, 8.0, 9.0, 10.0) and NaCl concentrations (2, 5, 7, 10% w/v) were performed according to the method described by Williams et al. (1983). In addition, carbon source utilization and acid production were studied using media and methods described by Gordon et al. (1974). The colours were determined by comparing with colour chips from the ISCC-NBS colour charts standard samples No. 2106 (Kelly, 1964).
Chemotaxonomy: The procedure of Lechevalier and Lechevalier (1980) was used for analysis of whole cell sugars. Cell wall amino acid were analysed by the methods of Jiang et al. (2001). Phospholipid analysis was carried out as described by Lechevalier et al. (1981). Menaquinones were determined using the procedures of Collins (1985). Fatty acid composition was determined by following the procedures of Sasser (1990) and Kampfer and Kroppenstedt (1996) by using the standard MIDI (Microbial identification) system.
Molecular analysis: The 16S rRNA gene was amplified and analysed as described by Li et al. (2007) and the 16S rRNA gene sequence (1463 nt) of the strain was determined. The variable γ region (position 158-277) of the 16S rRNA gene sequence and the almost complete 16S rRNA gene sequence of the strain was compared with closely related sequences of reference organisms. Sequence data were aligned with CLUSTAL_X (Thompson et al., 1997). The evolutionary tree rooted with Streptomyces viridis (AB184361.2) as the outgroup, was inferred by using maximum-likelihood method by Felsenstein (1981) with PHYLIP package. The topology of the resultant tree was evaluated based on 1000 resamplings (Felsenstein, 1985). Dendroscope program was used to display, edit and print phylogenetic trees (Huson et al., 2007).
The chromosomal DNA was extracted as described by Marmur (1961) and the genomic DNA G+C content of the strain was determined by thermal denaturation method of Marmur and Doty (1962).
Nucleotide sequence accession number: The almost complete 16S rRNA gene sequence of strain (1463 nucleotides) has been deposited in GenBank under the accession number HQ711986.
Antimicrobial activity: Spores from a mature slant culture of the actinomycete strain was inoculated aseptically into a 250 mL Erlenmeyer flask containing 50 mL production medium having the composition glucose 1%, soybean meal 1%, NaCl 1% and CaCO3 0.1% and incubated under submerged fermentation conditions at 28°C for 96 h at 180 rpm. After incubation, the fermented broth was centrifuged at 5000 rpm for 15 min. and the culture filtrate was assayed for antimicrobial activity using the agar well diffusion method. The assay plates were seeded with S. aureus (MTCC 3160), B. Subtilis (MTCC 441), B. cereus (MTCC 430), P. aeruginosa (MTCC 424), E. coli (MTCC 443), P. vulgaris (MTCC 426) using Muller Hinton agar and S. cerevisiae (MTCC 170), C. albicans (MTCC 227), A. niger (MTCC 961) and A. flavus (MTCC 3396) using yeast extract-malt extract agar were used for the antimicrobial assay, respectively. Culture filtrate (60 μL) was added to each well and the antimicrobial activity was observed after 24 h of incubation at 37°C for bacteria and 48 h of incubation at 25°C for fungi and the zone of inhibitions were expressed as diameter (mm).
RESULTS AND DISCUSSION
Characteristics of the isolate: Morphological observation of a 14 day culture of an actinomycete strain grown on yeast-malt extract agar (ISP-2 medium) revealed that strain had the typical characteristics of the genus Streptomyces.
 | |
| Fig. 1: | Scanning electron micrograph of the actinomycete strain grown on ISP2 medium for 2 weeks at 28°C |
| Table 1: | Culture characteristics of actinomycete strain on various media |
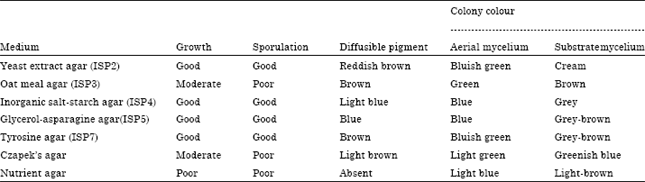 | |
| Note: Colors taken from ISCC-NBS COLOR CHARTS standard samples No 2106 Kelly (1964) | |
Aerial and vegetative hyphae were abundant, well developed without fragmentation. Long spore chains in spiral were borne on the aerial mycelium, the spores were rod shaped (1μm) with a spiny surface and non-motile (Fig. 1).
The Strain grew well on several media including yeast-malt extract agar (ISP2), Inorganic salt-starch agar (ISP4), glycerol-asparagine agar (ISP5) and tyrosine agar (ISP7). It exhibited moderate growth on oat meal agar (ISP3) and Czapek’s agar and showed poor growth on nutrient agar media (Table 1). Diffusible pigments were observed in test media and melanin was produced on peptone-yeast extract-iron agar. The strain could grow well between pH5.2 and 10.0, with an optimum pH value between 7.0-8.0. Growth was observed between 25-42°C, with an optimum temperature at 28°C. The strain grew in presence of 2-7% NaCl (w/v). The morphological, cultural characteristics and physiological characteristics of the strain were summarized in Table 2.
Chemotaxonomy of the isolate: The cell wall of the strain contained LL-Diaminopimelic acid and glycine as the diagnostic diamino acids which indicated that it, has a cell wall of chemotype I (Lechevalier and Lechevalier 1970a, b).
| Table 2: | Morphological, cultural and physiological characteristics of the actinomycete strain |
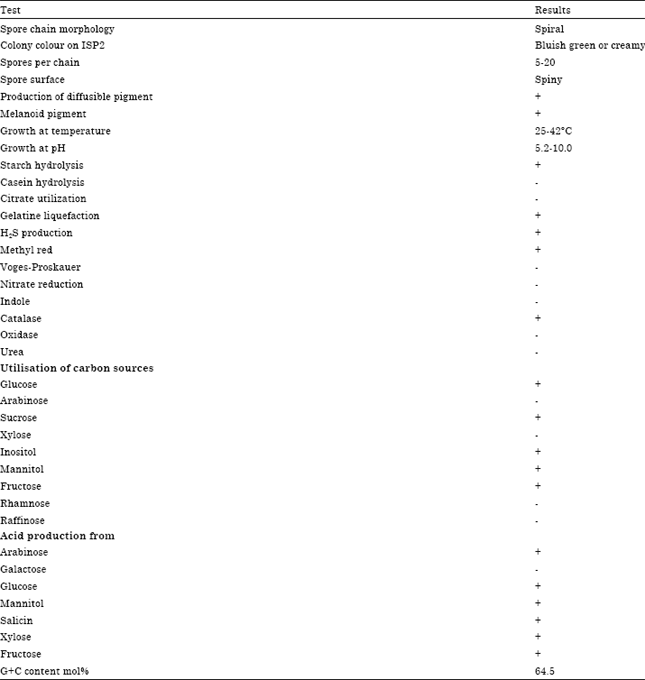 | |
| +: Utilization; -: Non-utilization | |
No diagnostic sugar was detected in whole cell hydrolysates. Phosphatidyl ethanolamine is a diagnostic phospholipid detected phospholipid type II sensu (Lechevalier et al., 1977). The predominant menaquinones were MK-9(H6) (81%) and MK-9(H8) (19%). The major fatty acids found were anteiso-C15:0(24.29%), iso-C16:0 (17.40%), iso-C14:0 (15.83%), iso-C15:0 (14.94%), C16:0 (12.22%), C14:0 (3.58%) and C16:1 ω7c/C15:0 iso 2-OH (2.21%) (Table 3). Thus chemotaxonomic and phenotypic data showed that the strain belongs to the genus Streptomyces.
| Table 3: | Fatty acid percentage profile of the actinomycete strain |
 | |
| ND: Not detected | |
Molecular characterization of the isolate: Analysis of the γ region sequences of the 16S rRNA from Streptomyces species showed that the strain was grouped into a branch with type strain of Streptomyces sp. ZG0650 (Fig. 2). The almost complete 16S rRNA gene sequence of the strain was determined in this study and has been deposited in the GenBank database (Accession No. HQ711986). This sequence was compared with the corresponding partial 16S rRNA sequence of the type strains of representative members of the genus Streptomyces retrieved from the public database by using BLAST (Altschul et al., 1997). Phylogenetic analysis revealed that the strain is a member of the genus Streptomyces. The comparative analysis of 16S rRNA gene sequence and the estimation of phylogenetic relationships showed that the strain formed a distinct lineage in the tree and showed the closest level of sequence similarity of 99.5% (a 6 nucleotide difference out of 1454 nt) with Streptomyces coeruleorubidus NBRC 12844T (GenBank accession No. AB184849). The sequence divergence values between the strain and the other members of Streptomyces ranged from 0.43% (a 6nucleotide difference out of 1425 nt) with Streptomyces coerulescens ISP5146T (GenBank accession No. AY999720) and 0.5% (a 7 nucleotide difference out of 1422 nt) with Streptomyces bellus ISP5185T (GenBank accession No. AJ399476) to 0.56% (a 8 nucleotide difference out of 1447 nt) with Streptomyces violaceochromogenes NBRC 13100T (GenBank accession No. AB184312). The G+C content of the DNA of the strain was 64.5 mol%.
Antimicrobial activity of the isolate: The antimicrobial activity of Streptomyces coeruleorubidus was screened against three Gram-positive and three Gram-negative bacteria and four fungal species. The strain inhibited the growth of all the test organisms, used in the study (Fig. 3, 4). This clearly suggests the broad spectrum nature of the bioactive secondary metabolite from marine Streptomyces.
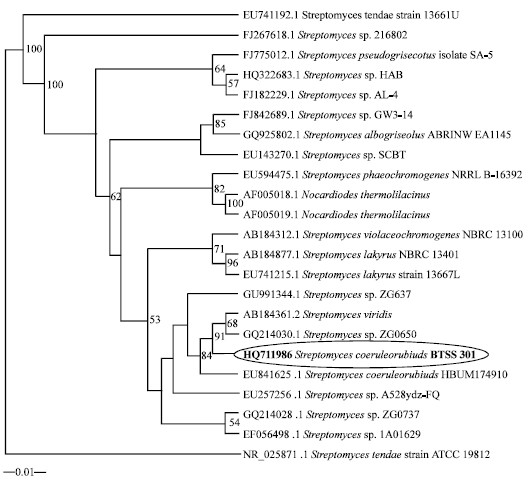 | |
| Fig. 2: | Phylogenetic dendrogram obtained by maximum likelihood method of 16S rRNA sequences, showing the position of strain Streptomyces coeruleorubidus among phylogenetic neighbours. Numbers on branch nodes are bootstrap values (1000 resamplings). The sequence of Streptomyces viridis (AB184361.2) was used as root bar, 1% sequence divergence |
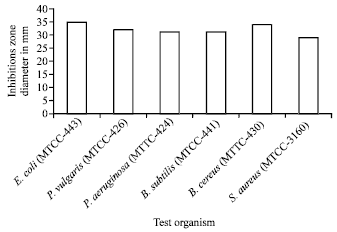 | |
| Fig. 3: | Antibacterial of Streptomyces coeruieorbidus sp. |
In a similar study by Reddy et al. (2011) and Mohd-Fuat et al. (2010) reported the broad spectrum nature of antibiotics from Streptomyces which inhibited the growth of both pathogenic bacteria and fungi while other studies, solely reported the anticandidal activity of Streptomyces (Sanasam and Ningthoujam, 2010).
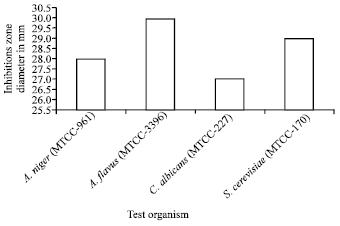 | |
| Fig. 4: | Antifungal activity of Streptomyces coeruieorbidus sp. |
Description of marine Streptomyces coeruleorubidus sp.: Gram positive, non-acid-fast and aerobic. Both vegetative and aerial hyphae are abundant, well developed and not fragmented. Vegetative hypha is extremely branched and does not bear any spores. Aerial mycelium appears usually after 14 days of incubation and varies from bluish green to light blue in the test media. At maturity, it forms spiral type of spore chains. Spore surface is spiny and non-motile. Casein is not hydrolysed. Positive in tests for starch hydrolysis and gelatine liquefaction. H2S is produced. Nitrate is not reduced. Soluble pigments are observed in all test media and melanin is observed in peptone-yeast extract-iron agar. The temperature range for growth is 25-42°C, with an optimum at 28°C. Grows at pH 5.2-10.0 and in the presence of 2- 7% (w/v) NaCl. Good growth is found on almost all carbon sources tested. Acid is formed from glucose, arabinose, mannitol, salicin, xylose and fructose as carbohydrate sources. Cell wall contains LL-DAP and glycine. Whole cell hydrolysates contain galactose and glucose and no diagnostic sugar. The predominant menaquinones were MK-9 (H6) (81%) and MK-9 (H8) (19%) and the diagnostic phospholipid is phosphatidyl ethanolamine. Major fatty acid components are anteiso-C15:0 (24.29%), iso-C16:0 (17.40%), iso-C14:0 (15.83%), iso-C15:0 (14.94%) and C16:0 (12.22%). The DNA G+C content is 64.5 mol%. The strain isolated from a marine sediment sample collected from Bay of Bengal near Visakhapatnam coast andhra Pradesh province, india and has been deposited in the microbial type culture collection and Gene Bank, institute of microbial technology (MTCC; IMTECH), Chandigarh as strain MTCC 33992T.
CONCLUSION
It is the first of its kind to report the Streptomyces coeruleorubidus from marine sediment, producing bioactive compound. After all these years there is an exponential increase in the number of new bioactive secondary metabolites which were produced by the actinomycetes isolated from marine source. They have the ability to produce potent, unique, modified, unprecedented bioactive secondary metabolites. The complex natures of these compounds enable them to be used as potent therapeutic agents to treat the emerging challenging diseases.
ACKNOWLEDGMENTS
This research was supported by the Ministry of Earth Sciences (MoES), Government of India, New Delhi (Sanction No. MoES/11-MRDF/1/26/P/08). We are thankful to the Department of Science and Technology to Promote University Research for Scientific Excellence (DST-PURSE) programme studies for providing the JEOL; JSM-6610LV scanning electron microscope studies.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Arunachalam, R., E.G. Wesely, J. George and G. Annadurai, 2010. Novel approaches for identification of Streptomyces noboritoensis TBG-V20 with cellulase production. Curr. Res. Bacteriol., 3: 15-26.
CrossRefDirect Link - Felsenstein, J., 1981. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol., 17: 368-376.
CrossRefPubMedDirect Link - Felsenstein, J., 1985. Confidence limits on phylogenesis: An approach using the bootstrap. Evolution, 39: 783-791.
Direct Link - Fiedler, H.P., C. Bruntner, A.T. Bull, A.C. Ward and M. Goodfellow et al., 2005. Marine actinomycetes as a source of novel secondary metabolites. Antonie van Leeuwenhoek, 87: 37-42.
CrossRef - Goodfellow, M., 1988. Numerical taxonomy and selective isolation of industrially important actinomycetes. Actinomycetologica, 2: 13-19.
CrossRefDirect Link - Huson, D.H., D.C. Richter, C. Rausch, T. Dezulian, M. Franz and R. Rupp, 2007. Dendroscope: An interactive viewer for large phylogenetic trees. BMC Bioinformatics, 8: 460-460.
PubMed - Imada, C., 2005. Enzyme inhibitors and other bioactive compounds from marine actinomycetes. Antonie Van Leewenhoek, 87: 59-63.
CrossRefDirect Link - Jensen, P.R., R. Dwight and W. Fenical, 1991. Distribution of actinomycetes in near-shore tropical marine sediments. Applied Environ. Microbiol., 57: 1102-1108.
Direct Link - Jiang, L., M. Li, W. Li, X. Cui, L. Xu and C. Jiang, 2001. Study on the application of quantitative analysis of cell-wall amino acids in actinomycetes classification. Acta Microbiol. Sin., 41: 270-277.
PubMed - Kampfer, P. and R.M. Kroppenstedt, 1996. Numerical analysis of fatty acid patterns of Coryneform bacteria and related taxa. Can. J. Microbiol., 42: 989-1005.
CrossRefDirect Link - Khucharoenphaisan, K., N. Sripairoj and K. Sinma, 2012. Isolation and identification of actinomycetes from termite's gut against human pathogen. Asian J. Anim. Vet. Adv., 7: 68-73.
CrossRefDirect Link - Lam, K.S., 2006. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol., 9: 245-251.
CrossRefPubMedDirect Link - Lechevalier, H.A. and M.P. Lechevalier, 1970. A Critical Evaluation of the Genera of Aerobic Actinomycetes. In: The Actinomycetales, Prauser, H. (Eds.). Gustav Fischer Verlag, Jena, Germany, pp: 393-405.
Direct Link - Lechevalier, M.P., C. De Bievre and H.A. Lechevalier, 1977. Chemotaxonomy of aerobic actinomycetes: Phospholipid composition. Biochem. Syst. Ecol., 5: 249-260.
CrossRefDirect Link - Lechevalier, M.P. and H. Lechevalier, 1970. Chemical composition as a criterion in the classification of aerobic actinomycetes. Int. J. Syst. Evol. Microbiol., 20: 435-443.
CrossRefDirect Link - Li, W.J., P. Xu, P. Schumann, Y.Q. Zhang and R. Pukall et al., 2007. Georgenia ruanii sp. nov., a novel actinobacterium isolated from forest soil in Yunnan (China) and emended description of the genus Georgenia. Int. J. Syst. Evol. Microbiol., 57: 1424-1428.
CrossRef - Maldonado, L.A., J.E. Stach, W. Pathom-aree, A.C. Ward, A.T. Bull and M. Goodfellow, 2005. Diversity of cultivable actinobacteria in geographically widespread marine sediments. Antonie van Leeuwenhoek, 87: 11-18.
PubMed - Marmur, J., 1961. A procedure for the isolation of deoxyribonucleic acid from micro-organisms. J. Mol. Biol., 3: 208-218.
CrossRefDirect Link - Marmur, J. and P. Doty, 1962. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J. Mol. Biol., 5: 109-118.
CrossRefPubMedDirect Link - Mohd-Fuat, A.R., A.R.B. Amini and O.M. Hasyima, 2010. Isolation of Streptomyces with broad spectrum antifungal activity from polyherbal products. Int. J. Bot., 6: 259-265.
CrossRefDirect Link - Narayana, K.P.J. and M. Vijayalakshmi, 2008. Production of Extracellular α-amylase by Streptomyces albidoflavus. Asian J. Biochem., 3: 194-197.
Direct Link - Narayana, K.J.P. and M. Vijayalakshmi, 2008. Production of extracellular protease by Streptomyces albidoflavus. Asian J. Biochem., 3: 198-202.
CrossRefDirect Link - Raja, A. and P. Prabakarana, 2011. Actinomycetes and drug-An overview. Am. J. Drug Discovery Dev., 1: 75-84.
CrossRefDirect Link - Kannan, R.R. and S.G.P. Vincent, 2011. Molecular characterization of antagonistic Streptomyces isolated from a mangrove swamp. Asian J. Biotechnol., 3: 237-245.
CrossRefDirect Link - Reddy, N.G., D.P.N. Ramakrishna and S.V.R. Gopal, 2011. A morphological, physiological and biochemical studies of marine Streptomyces rochei (MTCC 10109) showing antagonistic activity against selective human pathogenic microorganisms. Asian J. Biol. Sci., 4: 1-14.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Kumar, K.S., R. Haritha, Y.S.Y.V.J. Mohan and T. Ramana, 2011. Screening of marine actinobacteria for antimicrobial compounds. Res. J. Microbiol., 6: 385-393.
CrossRef - Stach, J.E.M., L.A. Maldonado, A.C. Ward, M. Goodfellow and A.T. Bull, 2003. New primers for the class Actinobacteria: pplication to marine and terrestrial environments. Environ. Microbiol., 5: 828-841.
PubMed - Sanasam, S. and D.S. Ningthoujam, 2010. Screening of local actinomycete isolates in manipur for anticandidal activity. Asian J. Biotechnol., 2: 139-145.
CrossRefDirect Link - Takizawa, M., R.R. Colwell and R.T. Hill, 1993. Isolation and diversity of actinomycetes in the Chesapeake Bay. Applied Environ. Microbiol., 59: 997-1002.
PubMedDirect Link - Thompson, J.D., T.J. Gibson, F. Plewniak, F. Jeanmougin and D.G. Higgins, 1997. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res., 25: 4876-4882.
CrossRefPubMedDirect Link - Waksman, S.A. and A.T. Henrici, 1943. The nomenclature and classification of the actinomycetes. J. Bacteriol., 46: 337-341.
PubMed - Williams, S.T., M. Goodfellow, G. Alderson, E.M.H. Wellington, P.H.A. Sneath and M.J. Sackin, 1983. Numerical classification of Streptomyces and related genera. J. Gen. Microbiol., 129: 1743-1813.
CrossRefPubMedDirect Link - Gordon, R.E., D.A. Barnett, J.E. Handerhan and C.H.N. Pang, 1974. Nocardia coeliaca, Nocardia autotrophica and the Nocardin strain. Int. J. Syst. Evolut. Microbol., 24: 54-63.
CrossRef - McCarthy, A.J., 1987. Lignocellulose degrading actinomycetes. FEMS Microbiol. Lett., 46: 145-163.
CrossRef