
Aakaknsha Khandelwal
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar-125 004, India
S.S. Sindhu
Department of Microbiology, College of Basic Sciences and Humanities, CCS Haryana Agricultural University, Hisar-125 004, India
Research Journal of Microbiology
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 158-170







ABSTRACT
Clusterbean (Cyamopsis tetragonoloba L. Taub.) is a commercially important crop but its nodulation status is poor under field conditions. Recently, ACC (1-aminocyclopropane-1-carboxylate) deaminase containing rhizobia have been found to improve nodulation and plant growth in various legumes by lowering the level of ethylene, which acts as inhibitor of nodulation. In this study, 95 isolates of Bradyrhizobium/Rhizobium were obtained from the nodules of clusterbean. These rhizobia were screened for utilization of ACC on Dworkin and Foster’s minimal medium and 46.3% Bradyrhizobium/Rhizobium isolates showed the ability to grow on ACC supplemented plates. Seed inoculation of clusterbean with different Bradyrhizobium/Rhizobium isolates caused significant root and shoot elongation at 5 and 10 days of growth on water agar plates. Maximum root and shoot elongation was observed by ACC utilizing isolate GSA11 followed by ACC non-utilizing isolates GSA114 and GSA115 at 5 and 10 days as compared to control uninoculated seedlings. ACC deaminase containing rhizobial isolates GSA6, GSA11 and GSA110 formed 42-48 nodules per plant and caused 80.7-205.3% gain in plant dry weight in comparison to uninoculated plants in sterilized chillum jar assembly conditions at 60 days of plant growth. Whereas, ACC deaminase lacking bacterial strains GSA114 and GSA115 caused only 35.7 and 3.6% increase in plant dry weight, respectively. These ACC utilizing rhizobial cultures could be further exploited for inoculation of clusterbean under field conditions.
PDF Abstract XML References Citation
Received: January 16, 2012;
Accepted: February 17, 2012;
Published: June 21, 2012
How to cite this article
Aakaknsha Khandelwal and S.S. Sindhu, 2012. Expression of 1-aminocyclopropane-1-carboxylate Deaminase in Rhizobia Promotes Nodulation and Plant Growth of Clusterbean (Cyamopsis tetragonoloba L.). Research Journal of Microbiology, 7: 158-170.
URL: https://scialert.net/abstract/?doi=jm.2012.158.170
URL: https://scialert.net/abstract/?doi=jm.2012.158.170
INTRODUCTION
Ethylene affects many vital processes of plant development such as seed germination, morphogenesis, flowering induction, fruit ripening, root elongation, lateral root growth and root hair formation (Binder, 2008; Shaharoona et al., 2011). This gaseous plant hormone, ethylene has been found to function as an autoregulatory signal to control lateral root development and nodule formation in legumes (Ligero et al., 1991; Desbrosses and Stougaard, 2011). Ethylene has been reported to inhibit nodulation in Medicago sativa and Pisum sativum (Lee and LaRue, 1992; Caba et al., 1998; Ma et al., 2003b). Exogenous application of ethylene or ethylene precursors (ethephon and ACC) also inhibited nodulation of P. sativum, mungbean and Discaria trinervis (Goodlass and Smith, 1979; Duodu et al., 1999; Valverde and Wall, 2005). Different chemicals such as L-a-(aminoethoxyvinyl)-glycine (AVG), silver thiosulfate, aminooxyacetic acid (AOA) and rhizobitoxine were found to lower either ethylene levels or its perception in plants and increased the number of nodules in pea, Lotus japonicus, Macroptilium atropurpureum and Phaseolus vulgaris (Guinel and LaRue, 1992; Nukui et al., 2000; Yuhashi et al., 2000; Tamimi and Timko, 2003). But, these chemicals are either expensive or potentially harmful to the environment.
Plant enzyme ACC synthase converts S-adenosylmethionine to ACC (Kende, 1993) and ACC in the root exudates is converted to ethylene by the enzyme ACC oxidase. However, the production of ethylene is highly regulated by plants (Shaharoona et al., 2007). Moreover, some beneficial rhizosphere bacteria naturally offset the inhibitory effect of ethylene by expressing the enzyme ACC deaminase (Glick, 2004; Shaharoona et al., 2011). This ACC deaminase enzyme cleaves plant-produced ACC into ammonia and a-ketobutyrate, both of which are readily metabolized by most soil bacteria and thereby, it lowers the ethylene level. The enzyme ACC deaminase has been reported in large number of soil microorganisms (Jacobson et al., 1994; Wang et al., 2000; Belimov et al., 2001) and these plant growth-promoting rhizosphere bacteria (PGPR) were found to facilitate plant growth by lowering of plant’s ethylene concentration (Glick et al., 1999; Glick, 2004). As a consequence of inhibition of ethylene biosynthesis and action, nodulation by rhizobia on legumes is enhanced in alfalfa and Lotus japonicus (Nukui et al., 2000), peas (Ma et al., 2003a, 2004); chickpea (Mann et al., 2002) and Medicago truncatula (Prayitno and Mathesius, 2010).
Clusterbean (C. tetragonoloba) occupies a very important place among commercially utilizable crops in India and is an important drought tolerant legume of arid zone. Clusterbean (guar) is a rich source of high quality galactomannan gum which is in great demand in the world market because of it multi-purpose use in textiles, foods, cosmetics, mining, explosives and oil industries. Protein rich guar meal is also used as animal feed and clusterbean is also emerging as a potential source of vegetable protein for human beings. Like Macroptilium atropurpureum, clusterbean is compatible for nodulation with both fast-growing Rhizobium and slow-growing Bradyrhizobium species (Jordan, 1984) but its nodulation status is poor with only 5 to 10 nodules per plant (Stafford and Lewis, 1980; Yadav, 1980). Though legume-Rhizobium symbiosis has been extensively studied in many crops but no systematic work has been done to exploit the nodulation and nitrogen-fixing ability of clusterbean crop for increasing its production.
MATERIALS AND METHODS
This study, was carried out in the Department of Microbiology, CCS Haryana Agricultural University, Hisar during the month of July 2008 to June 2010.
Isolation of Bradyrhizobium/Rhizobium strains: Plant roots of clusterbean bearing nodules were collected from different locations. Healthy pink nodules were surface sterilized by immersing in 0.1% HgCl2 solution for 5 min. Then nodules were washed with sterilized water 5 times to get rid of the sterilizing agent. The surface sterilized nodules were crushed in sterilized Petri plates and the suspension was streaked on yeast extract mannitol agar (YEMA) medium plates (Sindhu et al., 1999). The plates were incubated for 3 to 8 days at 28±2°C and 95 isolates of Bradyrhizobium/Rhizobium were obtained from these nodules. Isolated colonies of rhizobia were transferred on YEMA medium slopes and stored in the refrigerator for further studies. For inoculation purposes, seeds of clusterbean, commonly known as guar, were obtained from Department of Seed Science and Technology, CCS Haryana Agricultural University, Hisar.
Screening of Bradyrhizobium/Rhizobium for utilization of ACC: The medium plates were prepared with minimal medium (Dworkin and Foster, 1958) supplemented with ammonium sulphate (2 g L-1) or 3 mM ACC (Penrose and Glick, 2003). A loopful of 3 days old growth of the Bradyrhizobium/Rhizobium cultures was spotted on the medium plates. The growth of bacterial isolates on ACC supplemented medium plates was recorded after 2-5 days of incubation at 28±2°C. The bacterial cultures showing good growth on ACC supplemented medium plates and capable of utilizing ACC as nitrogen source, were scored as ACC+.
Root and shoot elongation of clusterbean by ACC utilizing Bradyrhizobium/Rhizobium isolates: Seeds of clusterbean were surface sterilized with acidic alcohol (concentrated sulphuric acid: ethanol, 7:3) for 3 min and washed thoroughly with several changes of sterilized water. The surface sterilized seeds were inoculated with selected ACC utilizing and ACC non-utilizing Bradyrhizobium/Rhizobium cultures individually (Jangu and Sindhu, 2011). Growth of each bacterial culture was harvested in 3 mL sterilized water and used to inoculate 15 seeds of clusterbean. The inoculated seeds were grown on three plain water agar (0.8%) plates containing five seeds on each plate. The plates were incubated at 28±2°C. The observations for elongation/retardation of root and shoot growth of clusterbean seedlings were recorded at 5 and 10 days of growth.
Inoculation with Bradyrhizobium/Rhizobium for the nodulation and plant growth of clusterbean: Selected ACC utilizing and ACC non-utilizing Bradyrhizobium/Rhizobium isolates were checked for nodulation and plant growth on clusterbean variety HG563 in sterilized chillum jar assemblies containing washed river sand in the upper jar and Sloger’s nitrogen-free mineral salt solution (Sloger, 1969) in the lower assembly. Surface sterilized seeds of clusterbean were inoculated with 5 mL of culture (107-108 cells mL-1 of growth suspension) and inoculated seeds were sown in autoclaved chillum jar assemblies (Sindhu et al., 1999). Uninoculated seeds were sown as control. After germination, three healthy seedlings were kept in each chillum jar. The crop was sown during the month of April 2009 to June 2009. The jars were kept in a net house under normal sunlight conditions and quarter-strength Sloger’s nitrogen-free mineral salt solution was used for watering as and when required. The observations for nodulation and plant growth were recorded at 30, 45 and 60 days of plant growth.
RESULTS
Isolation of bacterial strains: During this study, plant roots of clusterbean having nodules were collected from village Kharar-Alipur, Lalpura, Raipur (belonging to Hisar district) and village Kurangawali of Sirsa district in Haryana. Ninety five isolates of Bradyrhizobium/Rhizobium were obtained from the nodules of clusterbean by streaking crushed nodule suspension on YEMA medium plates.
Screening of Bradyrhizobium/Rhizobium isolates for ACC utilization: All the Bradyrhizobium/Rhizobium isolates were spotted on ammonium sulphate (2 g L-1) containing medium (control) and ACC (3 mM) supplemented medium plate. Observations for bacterial growth were taken after 3 days of incubation. On the basis of bacterial growth on two medium plates, Bradyrhizobium/Rhizobium isolates were divided in 6 categories (Table 1, Fig. 1). Seven cultures i.e., GSA5, GSA13, GSA34, GSA54, GSA61, GSA71 and GSA102 did not grow on both type of medium plates and they may require some amino acid for growth on Dworkin and Foster medium. The frequency of ACC utilizing isolates was found 46.3%. In 2nd category, cultures GSA7, GSA25, GSA39 and GSA110 showed slight growth on both the plates. The cultures GSA9, GSA10, GSA14, GSA20, GSA23, GSA30, GSA31, GSA32, GSA38, GSA43, GSA48 and GSA55 showed similar growth on both the medium plates.
 | |
| Fig. 1: | Growth of Bradyrhizobium/Rhizobium isolates on Dworkin and Foster minimal medium containing either ammonium sulphate or ACC |
| Table 1: | Screening of Rhizobium/Bradyrhizobium isolates for utilization of ACC |
 | |
| Growth of bacterial isolates was tested on minimal medium (Dworkin and Foster, 1958) supplemented with ammonium sulphate (2 g L-1) or 3 mM ACC. On the basis of colony size after 2-5 days of incubation at 28±2°C, the growth of the isolates was scored as, - : No growth,+: Little growth, ++ : Moderate growth, +++: Significant growth, ++++: Maximum growth | |
Three isolates GSA15, GSA24 and GSA106 showed significant growth on ammonium sulphate plate and less growth on ACC supplemented plate. In 5th category, twenty four isolates showed more growth on ammonium sulphate plates and slightly less growth on ACC supplemented plate. Isolates GSA50, GSA53, GSA101, GSA114 and GSA115 along with 40 other isolates showed good growth on ammonium sulphate plate but did not grow on ACC supplemented plate. On the basis of ACC utilization and non utilization pattern, nine cultures including GSA6 and GSA11 (as ACC utilizing) and GSA114 and GSA115 (as ACC non-utilizing) were selected for further studies.
Effect of ACC utilizing Bradyrhizobium/Rhizobium isolates on root and shoot elongation of clusterbean: Selected ACC utilizing and non-utilizing Bradyrhizobium/Rhizobium isolates were screened for root and shoot elongation of clusterbean on water agar plates.
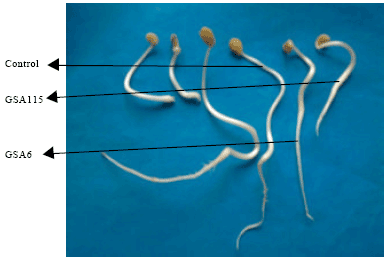 | |
| Fig. 2: | Effect of ACC utilizing and non-utilizing Rhizobium isolates on root and shoot elongation of clusterbean at 10 days of growth |
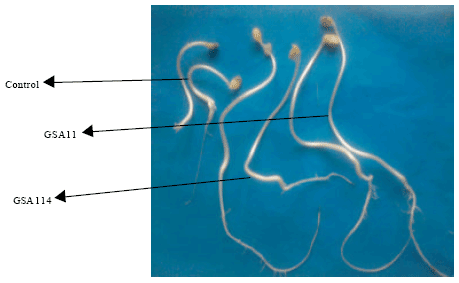 | |
| Fig. 3: | Effect of ACC utilizing and non-utilizing Bradyrhizobium isolates on root and shoot elongation of clusterbean at 10 days of growth |
In control uninoculated plates, root and shoot growth was not observed at 5 days but some growth was observed at 10 days (Table 2, Fig. 2, 3). All the isolates showed stimulation of root growth at 5 days. However, no effect of bacterial inoculation was observed on shoot growth at five days and significant stimulation of shoot growth was observed at 10 days of growth. Maximum root and shoot elongation in clusterbean was observed by ACC utilizing isolate GSA11 followed by ACC non-utilizing isolates GSA114 and GSA115 at 5 and 10 days as compared to control uninoculated seedlings. Isolate GSA26 showed stimulating effect on root length but stunting effect was observed on shoot length 10 days of growth as compared to control. Isolate GSA106 showed stunting effect on root length at both the stages of observation but stimulating effect was observed on shoot length at 10 days of growth as compared to control (Table 2).
| Table 2: | Effect of ACC utilizing Bradyrhizobium/Rhizobium isolates for root and shoot elongation of clusterbean on water agar plates |
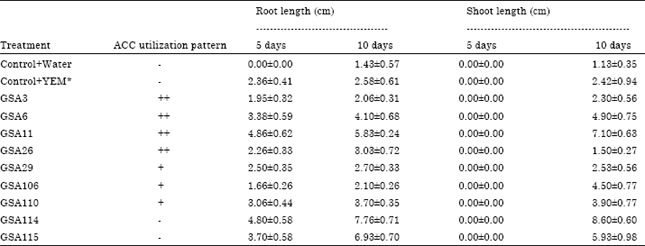 | |
| YEM* stands for yeast extract mannitol broth, Bradyrhizobium/Rhizobium isolates which showed variation in their ACC utilization ability on minimal medium (Dworkin and Foster, 1958) supplemented with 3 mM ACC were used. ACC utilization pattern of the isolates is indicated as, - : No growth, + : Little growth, ++ : Moderate growth | |
Screening of Bradyrhizobium/Rhizobium isolates for nodulation of clusterbean: A total of nine ACC utilizing and non-utilizing Bradyrhizobium/Rhizobium isolates were checked for nodulation and plant growth on clusterbean variety HG563 in sterilized chillum jar assemblies under pot house conditions. Observations for nodulation and plant growth were taken after 30, 45 and 60 days of sowing. Isolate GSA110 followed by GSA6 and GSA11 formed maximum nodules ranging from 10-14 at 30 days of growth and also significantly increased the plant dry weight as compared to control uninoculated treatment (Table 3). Inoculation with isolates GSA110, GSA6 and GSA11 increased the plant dry weight 100.6, 36.7 and 25.9%, respectively as compared to control uninoculated plants. Other isolates GSA3, GSA61, GSA74, GSA114 and GSA115 produced only 3-7 nodules per plant at 30 days of growth. At 45 days of plant growth, isolates GSA6, GSA11 and GSA110 produced maximum number of nodules ranging from 35 to 42 nodules per plant and also significantly increased plant dry weight (Table 3). The increase in the plant dry weight was 232.7% by isolate GSA11, 92.9% by GSA6 and 146.7% by isolate GSA110 as compared to control uninoculated plants. Five isolates GSA3, GSA61, GSA74, GSA114 and GSA115 formed only 7-20 nodules per plant. Three isolates GSA3, GSA61 and GSA115 showed very little increase in plant dry weights at both the stages of plant growth.
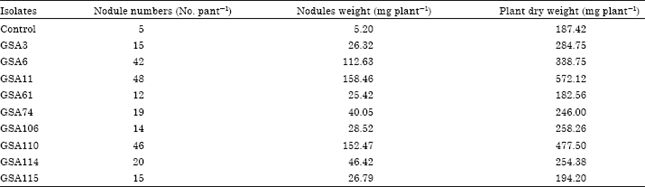
At 60 days of plant growth, ACC containing rhizobial isolates GSA6, GSA11 and GSA110 produced 42, 48 and 46 nodules per plant, respectively (Table 4). Inoculation of ACC containing rhizobial isolates GSA6 and GSA110 increased the plant dry weight by 80.7 and 154.8% and inoculation with isolate GSA11 increased the plant dry weight by 205.3% in comparison to uninoculated plants. Whereas ACC lacking bacterial strains GSA114 and GSA115 caused only 35.7 and 3.6% increase in plant dry weight, respectively as compared to control uninoculated plants. Nodule fresh weight was found maximum by inoculation of Bradyrhizobium isolate GSA11 followed by Rhizobium isolates GSA110 and GSA6. Thus, ACC utilizing Rhizobium/Bradyrhizobium isolates enhanced nodulation in clusterbean and resulted in more gain in plant growth. More nodules were formed by Bradyrhizobium isolates than the Rhizobium isolates.
| Table 3: | Symbiotic effectiveness of selected Bradyrhizobium/Rhizobium isolates in clusterbean at 30 and 45 days of plant growth |
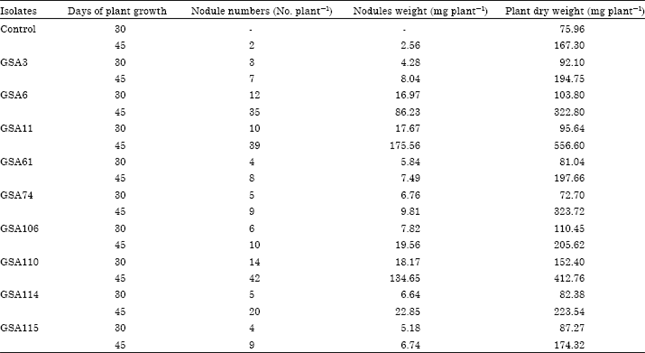 | |
| - : No nodule formed on uninoculated plants | |
| Table 4: | Symbiotic effectiveness of selected Bradyrhizobium/Rhizobium isolates in clusterbean at 60 days of plant growth |
 | |
DISCUSSION
Biological nitrogen fixation plays a dominant role in sustainable agricultural production (Bohlool et al., 1992) and a variety of N2-fixing bacteria have been used to improve the supply of fixed N as nutrient to leguminous and nonleguminous crop plants (Franche et al., 2009; Sindhu et al., 2010; Verma et al., 2010). Invasion of roots by Rhizobium during nodule formation exerts biotic stress and increases the ACC content of the infected roots resulting in enhanced ethylene production in alfalfa (Caba et al., 1998), soybean (Hunter, 1993) and Vicia sativa (Van Workum et al., 1995). This increased ethylene levels probably could be one of the factors for poor nodulation of clusterbean under field conditions. Therefore, it is important that efficient Rhizobium/Bradyrhizobium cultures possessing ACC deaminase activity should be isolated and introduced in clusterbean growing areas. With this objective, 95 isolates of Bradyrhizobium and Rhizobium were screened for their ability to utilize ACC and selected isolates were tested in chillum jar assemblies for nodulation of clusterbean variety HG563 under pot house conditions.
All the isolated Bradyrhizobium/Rhizobium cultures were checked for utilization of ACC. The cultures GSA9, GSA10, GSA14, GSA20, GSA23, GSA30, GSA31, GSA32, GSA38, GSA43, GSA48 and GSA55 showed similar growth on Dworkin and Foster medium supplemented with either ammonium sulphate or ACC (Table 1). Twenty four Bradyrhizobium/Rhizobium isolates showed more growth on ammonium sulphate containing plates and slightly less growth on ACC supplemented medium plate whereas isolates GSA50, GSA53, GSA101, GSA114 and GSA115 showed good growth on ammonium sulphate plate but did not grow on ACC supplemented plate. Rest forty five isolates showed good growth on ammonium sulphate supplemented medium but did not grow on ACC supplemented medium plate indicating that these cultures lacked ACC utilization activity.
In this study, 46.3% of Bradyrhizobium/Rhizobium isolates showed the ability to utilize ACC. Ma et al. (2003b) also observed that only 38.76% rhizobial strains (five positive out of 13 strains tested) possess ACC deaminase enzyme. Whereas, screening of 563 bacteria isolated from the roots of pea, lentil and chickpea for plant growth promotion ability showed that only 5% isolates showed ACC deaminase activity (Hynes et al., 2008). Similarly, 20 bacterial isolates out of 236 of plant growth promoting rhizobacteria were found to contain ACC deaminase activity based on their ability to grow on ACC containing plates using ACC as a sole nitrogen source (Govindasamy et al., 2009). Duan et al. (2009) screened 233 rhizobial strains collected from different locations across Saskatchewan, Canada for ACC deaminase activity and only 27 strains displayed the activity. In contrast, high frequency of ACC utilizing isolates was reported by Husen et al. (2009) in which 11 Pseudomonas isolates (out of total 13 isolates) obtained from the soybean rhizosphere were found to possess ACC deaminase activity and increased root development of soybean. Similarly, 40% plant-associated Burkholderia species (out of 45 strains tested) exhibited ACC deaminase activity (Onofre-Lemus et al., 2009).
Selected ACC utilizing Bradyrhizobium/Rhizobium isolates were screened for root and shoot elongation of clusterbean on water agar plates. In control plates, root and shoot growth of clusterbean was not observed at five days (Table 2; Fig. 2, 3). All the Bradyrhizobium/Rhizobium isolates showed stimulation of root growth at five days whereas no effect of bacterial treatment was observed on shoot length. Significant stimulation of shoot growth was observed at 10 days of growth. Maximum root and shoot elongation in clusterbean was observed by isolates GSA11 followed by GSA114, GSA115 and GSA6 at both the stages of observation as compared to control uninoculated plants. Isolate GSA26 showed the stimulating effect on root length but stunting effect was observed on shoot length as compared to control (Table 2). Two isolates lacking ACC utilization ability i.e., GSA114 and GSA115 also showed significant stimulation of both root and shoot at 10 days of growth.
Lifshitz et al. (1987) reported that most of the Pseudomonas isolates showed retardation effect on root length of canola seedlings as compared to control and similarly treatment of wheat seedlings with a cell free culture filtrate of P. fluorescens had a strong inhibitory effect on root elongation (Astrom et al., 1993). In contrast, seed germination of barley (Hordeum vulgare L.) was significantly enhanced by application of lipo-chitooligosaccharides produced by Rhizobium and gibberellins (Miransari and Smith, 2009). Another beneficial rhizobacterial P. putida strain GR12-2 was reported to stimulate root elongation of different plants and this bacterium was found to contain ACC deaminase (Jacobson et al., 1994). Mutants of this strain lacking ACC deaminase activity did not promote root elongation of canola seedlings, implicating that ACC deaminase enzyme is involved in the root growth promotion mechanism of this bacterium (Glick et al., 1994). Penrose and Glick (2003) also reported that ACC utilizing bacterial strains stimulated root elongation in canola (Brassica campestris) seedlings under gnotobiotic conditions.
Selected ACC utilizing and non-utilizing Bradyrhizobium/Rhizobium isolates were checked for nodulation and plant growth on clusterbean variety HG563 in sterilized chillum jar assemblies. Different Bradyrhizobium/Rhizobium isolates formed nodules that varied from 3-14 nodules per plant at 30 days of growth (Table 3). Isolate GSA110 formed maximum nodules and also significantly increased the plant dry weight by 100.6% as compared to uninoculated control plants. At 45 days of plant growth, isolates GSA6, GSA11 and GSA110 produced maximum nodules varying from 35-42 nodules per plant and 232.7% increase in plant dry weight was observed on inoculation with GSA11 isolate. Other isolates formed 7-20 nodules per plant and showed very little increase in plant dry weight. At the 60 days of plant growth, isolates GSA6, GSA11 and GSA110 produced large number of nodules that varied from 42-48 nodules per plant (Table 4). Thus, ACC utilizing Bradyrhizobium/Rhizobium isolates enhanced nodulation in clusterbean and more nodules were formed by Bradyrhizobium isolates than the Rhizobium isolates. Significant increases in plant dry weight i.e., 80.7 and 154.8% were observed by inoculation of ACC utilizing isolates GSA6 and GSA110. Isolates lacking ACC utilization ability, GSA114 and GSA115 showed only 3.6 and 35.7% gain in plant dry weight, respectively in comparison to control uninoculated plants (Table 4).
Inoculation of various Bradyrhizobium/Rhizobium strains alone or with PGPR strains have been found to significantly enhance nodulation, nitrogen fixation and plant growth of legumes (Vikram and Hamzehzarghani, 2008; Verma et al., 2010; Sahu and Sindhu, 2011). In earlier studies, 32 different cultivars of clusterbean (guar) were studied for nodulation status and the number of nodules ranged between 0.93-5.73. Maximum nodules were observed in guar cultivar HFG197 (Khurana et al., 1978). Yadav (1980) conducted a pot experiment to examine the response of guar cultivar FS277 to inoculation with selected Rhizobium strains 5A/70, Tal174, Tal169 and GR4 in field soil. Different guar cultivars responded differently to inoculation with different strains of Rhizobium. Maximum increase in shoot dry weight and nitrogen content was reported in the guar cultivars HFG75, HFG182, HFG405 and Pusa Nav Bahar after inoculation with Rhizobium strain Tal174 whereas, inoculation with Rhizobium strain 5A/70 showed maximum gain in shoot dry weight and nitrogen content in cultivars HFG119, FS277 and Durgapur Safed.
Ethylene is rapidly synthesized in response to abiotic as well as biotic stresses and attempts are being made to improve tolerance ability of crop plants to these stresses (Saleem et al., 2007; Roy et al., 2011). However, different plant species display different requirements and regulatory mechanisms for ethylene. For example, Lee and LaRue (1992) found that exogenous ethylene inhibited nodulation on the primary and lateral roots of pea (Pisum sativum L. cv. Sparkle). Hunter (1993) reported that nodulated soybean roots produced more ethylene and contained more ACC than uninoculated roots. The addition of the ethylene precursor ACC to plant roots was found to block nodulation in Medicago truncatula (Penmesta and Cook, 1997), whereas the addition of the ethylene inhibitor AVG increased the number of nodules (Nukui et al., 2000). The compound rhizobitoxine (an ethylene biosynthesis inhibitor) which is synthesized by the legume symbiont Bradyrhizobium elkanii and the plant pathogen Burkholderia andropogonis (Yasuta et al., 2001) has been found to enhance nodulation and competitiveness of the legumes Macroptilium atropurpureum (siratro) and Vigna radiata (mungbean) by inhibiting endogenous ethylene synthesis in the host plant (Duodu et al., 1999; Yasuta et al., 1999).
Ma et al. (2003a) found that ACC deaminase minus mutant of Rhizobium leguminosarum bv. viciae strain 128C53K formed approximately 25% fewer nodules than the wild-type strain on Pisum sativum L. cv. Sparkle and also produced approximately 30% less plant biomass. Similarly, a strain of S. meliloti was transformed with ACC deaminase gene (acdS) and its upstream regulatory gene (lrpL) from R. leguminosarum bv. viciae 128C53K. The transformed S. meliloti bacterium showed 35 to 40% greater efficiency in nodulating Medicago sativa (alfalfa), by reducing ethylene production in the host plants and produced nearly 40% more biomass than the non-transformed strain in treated alfalfa seedlings (Ma et al., 2004). Similarly, a significant correlation was found between in vitro ACC deaminase activity of the rhizobacteria and growth promoting activity of these bacteria on maize under axenic conditions and on nodulation of mungbean (Vigna radiata L.) under natural pot and field trials (Shaharoona et al., 2006).
CONCLUSION
The increased use of chemical fertilizers and pesticides in agriculture has helped in achieving self-sufficiency in food grains production but it has polluted the environment and is causing slow deterioration of soil health. Therefore, inoculation with nitrogen-fixing, phosphate solubilizing bacteria and PGPR strains has emerged as a supplemental technology to minimize the use of chemical fertilizers (Shaukat et al., 2006; Linu et al., 2009; Keyeo et al., 2011). ACC released by legume roots acts as precursor of stress hormone ethylene that was found to inhibit nodulation in different legumes. Bradyrhizobium/Rhizobium isolates were obtained from the nodules of clusterbean and 46.3% of these isolates showed ACC utilization ability. ACC utilizing isolate GSA11 followed by ACC non-utilizing isolates GSA114 and GSA115 showed maximum root and shoot elongation at 5 and 10 days as compared to control uninoculated seedlings. ACC containing rhizobial isolates i.e., GSA6, GSA11 and GSA110 formed maximum number of nodules (42-48 nodules per plant) and 80.7-205.3% gain in plant dry weight was observed in comparison to uninoculated plants in sterilized chillum jar assembly conditions at 60 days of plant growth. These results suggested the importance of ACC utilization by rhizobia in promoting nodulation and plant biomass in clusterbean. Therefore, rhizobial strains should first be selected/tested for the presence of a functional ACC deaminase before using these bacteria as inoculant under field conditions.
REFERENCES
- Astrom, B., A. Gustafsson and B. Gerhardson, 1993. Characteristics of a plant deleterious rhizosphere pseudomonad and its inhibitory metabolite(s). J. Applied Bacteriol., 74: 20-28.
CrossRefDirect Link - Belimov, A.A., V.I. Safronova, T.A. Sergeyeva, T.N. Egorova and V.A. Matveyeva et al., 2001. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol., 47: 642-652.
CrossRefDirect Link - Binder, B.M., 2008. The ethylene receptors: Complex perception for a simple gas. Plant Sci., 175: 8-17.
CrossRefDirect Link - Bohlool, B.B., J.K. Ladha, D.P. Garrity and T. George, 1992. Biological nitrogen fixation for sustainable agriculture: A perspective. Plant Soil, 141: 1-11.
CrossRefDirect Link - Caba, J.M., L. Recalde and F. Ligero 1998. Nitrate induced ethylene biosynthesis and the control of nodulation in alfalfa. Plant Cell Environ., 21: 87-93.
CrossRefDirect Link - Duan, J., K.M. Muller, T.C. Charles, S. Vesely and B.R. Glick, 2009. 1-Aminocyclopropane-1-Carboxylate (ACC) deaminase genes in rhizobia from Southern Saskatchewan. Microb. Ecol., 57: 423-436.
CrossRefPubMedDirect Link - Duodu, S., T.V. Bhuvaneswari, T.J.W. Stokkermans and N.K. Peters, 1999. A positive role for rhizobitoxine in Rhizobium-legume symbiosis. Mol. Plant-Microbe Interact., 12: 1082-1089.
CrossRefDirect Link - Dworkin, M. and J.W. Foster, 1958. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol., 75: 592-601.
Direct Link - Franche, C., K. Lindstrom and C. Elmerich, 2009. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil, 321: 35-59.
Direct Link - Glick, B.R., 2004. Bacterial ACC deaminase and the alleviation of plant stress. Adv. Applied Microbiol., 56: 291-312.
CrossRefDirect Link - Glick, B.R., C.B. Jacobson, M.M.K. Schwarze and J.J. Pasternak, 1994. 1-Aminocyclopropane-1-carboxylic acid deaminase mutants of the plant growth-promoting rhizobacteria Pseudomonas putida GR12-2 do not stimulate canola root elongation. Can. J. Microbiol., 40: 911-915.
Direct Link - Goodlass, G. and K.A. Smith, 1979. Effects of ethylene on root extension and nodulation of pea (Pisum sativum L.) and white clover (Trifolium repens L.). Plant Soil, 51: 387-395.
CrossRefDirect Link - Guinel, F.C. and T.A. LaRue, 1992. Ethylene inhibitors partly restore nodulation to pea mutant E107 (brz). Plant Physiol., 99: 515-518.
Direct Link - Hunter, W.J., 1993. Ethylene production by root nodules and effect of ethylene on nodulation in Glycine max. Applied Environ. Microbiol., 59: 1947-1950.
Direct Link - Husen, E., A.T. Wahyudi, A. Suwanto and R. Saraswati, 2009. Soybean seedling root growth promotion by 1-aminocyclopropane-1-carboxylate deaminase-producing pseudomonads. Indonesian J. Agric. Sci., 10: 19-25.
Direct Link - Hynes, R.K., G.C. Leung, D.L. Hirkala and L.M. Nelson, 2008. Isolation, selection and characterization of beneficial rhizobacteria from pea, lentil and chickpea grown in Western Canada. Can. J. Microbiol., 54: 248-258.
CrossRefPubMedDirect Link - Jacobson, C.B., J.J. Pasternak and B.R. Glick, 1994. Partial purification and characterization of 1-aminocyclopropane-1- carboxylate deaminase from the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol., 40: 1019-1025.
CrossRef - Jangu, O.P. and S.S. Sindhu, 2011. Differential response of inoculation with indole acetic acid producing Pseudomonas sp. in green gram (Vigna radiata L.) and black gram (Vigna mungo L.). Microbiol. J., 1: 159-173.
CrossRefDirect Link - Kende, H., 1993. Ethylene biosynthesis. Ann. Rev. Plant Physiol. Plant Mol. Biol., 44: 283-307.
CrossRefDirect Link - Keyeo, F., O.N. Ai'shah and H.G. Amir, 2011. The effects of nitrogen fixation activity and phytohormone production of diazotroph in promoting growth of rice seedlings. Biotechnol., 10: 267-273.
CrossRefDirect Link - Khurana, A.L., M.L. Saini, B.S. Jhorar and S.S. Dudeja, 1978. Nodulation studies in clusterbean. Forage Res., 4: 195-198.
Direct Link - Lee, K.H. and T.A. LaRue, 1992. Exogenous ethylene inhibits nodulation of Pisum sativum L. cv. Sparkle. Plant Physiol., 100: 1759-1763.
CrossRefPubMedDirect Link - Lifshitz, R., J.W. Kloepper, M. Kozlowshi, C. Simonson, J. Carlson, M. Tipping and I. Zalesha, 1987. Growth promotion of Canola (rapeseed) seedlings by a strain of Pseudomonas putida under gnotobiotic conditions. Can. J. Mic., 33: 390-395.
Direct Link - Ligero, F., J.M. Caba, C. Lluch and J. Olivares, 1991. Nitrate inhibition of nodulation can be overcome by the ethylene inhibitor aminoethoxyvinylglycine. Plant Physiol., 97: 1221-1225.
PubMedDirect Link - Linu, M.S., J. Stephen and M.S. Jisha, 2009. Phosphate solubilizing Gluconacetobacter sp., Burkholderia sp. and their potential interaction with cowpea (Vigna unguiculata (L.) Walp.). Int. J. Agric. Res., 4: 79-87.
CrossRefDirect Link - Ma, W., F.C. Guinel and B.R. Glick, 2003. Rhizobium leguminosarum Biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Applied Environ. Microbiol., 69: 4396-4402.
CrossRef - Ma, W., S.B. Sebestianova, J. Sebestian, G.I. Burd, F.C. Guinel and B.R. Glick, 2003. Prevalence of 1-aminocyclopropane-1-carboxylate deaminase in Rhizobium spp. Antonie Van Leeuwenhoek, 83: 285-291.
CrossRefPubMedDirect Link - Ma, W., T.C. Charles and B.R. Glick, 2004. Expression of an exogenous 1-aminocyclopropane-1-carboxylate deaminase gene in Sinorhizobium meliloti increases its ability to nodulate alfalfa. Applied Environ. Microbiol., 70: 5891-5897.
CrossRefPubMedDirect Link - Mann, A., A.S. Nandwal, I.S. Sheoran, B.S. Kundu and S. Sheokand et al., 2002. Ethylene evolution, H2O2 scavenging enzymes and membrane integrity of Cicer arietinum L. nodules as affected by nitrate and aminoethoxyvinylglycine. J. Plant Physiol., 159: 347-353.
CrossRefDirect Link - Miransari, M. and D. Smith, 2009. Rhizobial lipo-chitooligosaccharides and gibberellins enhance barley (Hordeum vulgare L.) seed germination. Biotechnology, 8: 270-275.
CrossRefDirect Link - Nukui, N., H. Ezura, K.I. Yuhashi, T. Yasuta and K. Minamisawa, 2000. Effects of ethylene precursor and inhibitors for ethylene biosynthesis and perception on nodulation in Lotus japonicus and Macroptilium atropurpureum. Plant Cell Physiol., 41: 893-897.
CrossRefDirect Link - Onofre-Lemus, J., I. Hernandez-Lucas, L. Girard and J. Caballero-Mellado, 2009. ACC (1-aminocyclopropane-1-carboxylate) deaminase activity, a widespread trait in Burkholderia species, and its growth-promoting effect on tomato plants. Applied Environ. Microbiol., 75: 6581-6590.
CrossRefPubMedDirect Link - Penmesta, R.V. and D.R. Cook, 1997. A legume ethylene-insensitive mutant hyperinfected by its rhizobial symbiont. Science, 275: 527-530.
CrossRefDirect Link - Penrose, D.M. and B.R. Glick, 2003. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiologia Plantarum, 118: 10-15.
CrossRefPubMedDirect Link - Prayitno, J. and U. Mathesius, 2010. Differential regulation of the nodulation zone by silver ions, L-α-(2-amino-ethoxyvinyl)-glycine and the skl mutation in Medicago truncatula. Hayati J. Biosci., 17: 15-20.
Direct Link - Roy, B., S.K. Noren, A.B. Mandal and A.K. Basu, 2011. Genetic engineering for Abiotic stress tolerance in agricultural crops. Biotechnology, 10: 1-22.
CrossRefDirect Link - Sahu, G.K. and S.S. Sindhu, 2011. Disease control and plant growth promotion of green gram by siderophore producing Pseudomonas sp. Res. J. Microbiol., 6: 735-749.
CrossRefDirect Link - Saleem, M., M. Arshad, S. Hussain and A.S. Bhatti, 2007. Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J. Ind. Microbiol. Biotechnol., 34: 635-648.
PubMed - Shaharoona, B., M. Arshad and A. Khalid, 2007. Differential response of etiolated pea seedlings to inoculation with rhizobacteria capable of utilizing 1-aminocyclopropane-1-carboxylate or L-methionine. J. Microbiol., 45: 15-20.
PubMedDirect Link - Shaharoona, B., M. Arshad, Z.A. Zahir and A. Khalid, 2006. Performance of Pseudomonas spp. containing ACC-deaminase for improving growth and yield of maize (Zea mays L.) in the presence of nitrogenous fertilizer. Soil Biol. Biochem., 38: 2971-2975.
CrossRefDirect Link - Shaharoona, B., M. Imran, M. Arshad and A. Khalid, 2011. Manipulation of ethylene synthesis in roots through bacterial ACC deaminase for improving nodulation in legumes. Crit. Rev. Plant Sci., 30: 279-291.
CrossRef - Shaukat, K., S. Affrasayab and S. Hasnain, 2006. Growth responses of Triticum aestivum to plant growth promoting rhizobacteria used as a biofertilizer. Res. J. Microbiol., 1: 330-338.
CrossRefDirect Link - Sindhu, S.S., S.K. Gupta and K.R. Dadarwal, 1999. Antagonistic effect of Pseudomonas spp. on pathogenic fungi and enhancement of growth of green gram (Vigna radiata). Biol. Fertil. Soils, 29: 62-68.
CrossRefDirect Link - Sloger, C., 1969. Symbiotic effectiveness and N2 fixation in nodulated soybean. Plant Physiol., 44: 1666-1668.
Direct Link - Tamimi, S.M. and M.P. Timko, 2003. Effects of ethylene and inhibition of ethylene synthesis and action on nodulation in common bean (Phaseoulus vulgaris L.). Plant Soil, 257: 125-131.
Direct Link - Van Workum, W.A.T., A.A.N. van Brussel, T. Tak, C.A. Wijffelman and W.J. Kijne, 1995. Ethylene prevents nodulation of Vicia sativa ssp. nigra by exopolysaccharide-deficient mutantsof Rhizobium leguminosarum bv. Viciae. Mol. Plant Microbe Interact., 8: 278-285.
CrossRefDirect Link - Valverde, C. and L.G. Wall, 2005. Ethylene modulates the susceptibility of the root for nodulation in actinorhizal Discaria trinervis. Physiol. Plant., 124: 121-131.
CrossRefDirect Link - Verma, J.P., J. Yadav and K.N. Tiwari, 2010. Application of Rhizobium sp. BHURC01 and plant growth promoting rhizobactria on nodulation, plant biomass and yields of Chickpea (Cicer arietinum L.). Int. J. Agric. Res., 5: 148-156.
CrossRefDirect Link - Vikram, A. and H. Hamzehzarghani, 2008. Effect of phosphate solubilizing bacteria on nodulation and growth parameters of greengram (Vigna radiata L. Wilczek). Res. J. Microbiol., 3: 62-72.
CrossRefDirect Link - Wang, C., E. Knill, B.R. Glick and G. Defago, 2000. Effect of transferring 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase genes into Pseudomonas fluorescens strain CHA0 and its gacA derivative CHA96 on their growth-promoting and disease-suppressive capacities. Can. J. Microbiol., 46: 898-907.
PubMedDirect Link - Yasuta, T., S. Okazaki, H. Mitsui, K.I. Yuhashi, H. Ezura and K. Minamisawa, 2001. DNA sequence and mutational analysis of rhizobitoxine biosynthesis genes in Bradyrhizobium elkanii. Applied Environ. Microbiol., 67: 4999-5009.
CrossRefDirect Link - Yasuta, T., S. Satoh and K. Minamisawa, 1999. New assay for rhizobitoxine based on inhibition of 1-aminocyclopropane-1-carboxylate synthase. Applied Environ. Microbiol., 65: 849-852.
PubMedDirect Link - Yuhashi, K.I., N. Ichikawa, H. Ezura, S. Akao and Y. Minakawa et al., 2000. Rhizobitoxine production by Bradyrhizobium elkanii enhances nodulation and competitiveness on Macroptilium atropurpureum. Applied Environ. Microbiol., 66: 2658-2663.
CrossRefPubMedDirect Link