
P.M.K. Njage
Laboratory of Food Biotechnology, Institute of Food Science and Nutrition, Swiss Federal Institute of Technology (ETH), Z�rich, Switzerland
S. Dolci
Laboratory of Food Biotechnology, Institute of Food Science and Nutrition, Swiss Federal Institute of Technology (ETH), Z�rich, Switzerland
C. Jans
Laboratory of Food Biotechnology, Institute of Food Science and Nutrition, Swiss Federal Institute of Technology (ETH), Z�rich, Switzerland
J. Wangoh
Department of Food Technology and Nutrition, College of Agriculture and Veterinary Sciences, University of Nairobi, P.O. Box 29053, Nairobi, Kenya
C. Lacroix
Laboratory of Food Biotechnology, Institute of Food Science and Nutrition, Swiss Federal Institute of Technology (ETH), Z�rich, Switzerland
L. Meile
Laboratory of Food Biotechnology, Institute of Food Science and Nutrition, Swiss Federal Institute of Technology (ETH), Z�rich, Switzerland
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 9 | Page No.: 678-692







ABSTRACT
Little information exists on yeasts involved in camel milk fermentation in the remote arid and semi-arid regions despite spontaneous nature of the fermentations. Yeast species present throughout the distribution process of raw and naturally fermented camel milk (suusac) were therefore studied. Identification was done using API 20C AUX, Restriction Fragment Length Polymorphism (RFLP) of intergenic spacers ITS1 and ITS2 using restriction endonucleases HhaI, HinfI and HaeIII and RAPD with (GTG)5, (GAC)5, (GACA)4 microsatellite primers and an M13 core sequence. DNA sequence analysis of either ITS1 or ITS2 or the 26S rRNA gene was performed. API enabled the identification of 80 (47%) yeasts isolates and resulted in either incorrect identification or inability to identify the others. RFLP, RAPD and sequence analysis enabled complete identification to species and some differentiation at strain level with RAPD allowing more discrimination within species. There were 21 yeast species belonging to the genera Rhodotorula, Cryptococcus, Candida, Saccharomyces, Trichosporon, Geotrichum and Issatchenkia. The most frequently isolated yeasts were Saccharomyces cerevisiae (19%), Candida inconspicua (12%), Trichosporon mucoides (11%), Candida famata (11%), Rhodotorula mucilaginosa (8%), Candida lusitaniae (6%), Cryptococcus laurentii (5%), Cryptococcus albidus (5%), Candida guilliermondii (5%) and Trichosporon cutaneum (5%). Lowest log cfu mL-1 for the identified yeasts were 2.4 for C. tropicalis and highest were 7.6 (range; 2.4-8.5) for C. famata and 8.0 (range; 2.6-8.5) for C. guilliermondii. A combination of phenotypic and molecular methods for proper yeast identification is recommended. Due to the high diversity and numbers of yeasts, their role in the fermented camel milk should be studied to enable exploitation of useful yeasts and prevention of contamination by detrimental yeasts.
PDF Abstract XML References Citation
Received: May 31, 2011;
Accepted: August 02, 2011;
Published: September 06, 2011
How to cite this article
P.M.K. Njage, S. Dolci, C. Jans, J. Wangoh, C. Lacroix and L. Meile, 2011. Characterization of Yeasts Associated with Camel Milk using Phenotypic and Molecular Identification Techniques. Research Journal of Microbiology, 6: 678-692.
URL: https://scialert.net/abstract/?doi=jm.2011.678.692
URL: https://scialert.net/abstract/?doi=jm.2011.678.692
INTRODUCTION
Numerous naturally fermented foods and drinks have played key roles in the diets of African people (Jespersen, 2003). Such natural fermentation is a result of a microbial consortium predominantly composed of Lactic Acid Bacteria (LAB). Additionally, yeasts have been enumerated and identified in a wide range of African fermented food products including milk (Fleet, 2007; Gadaga et al., 2000, 2001; Jespersen, 2003; Kebede et al., 2007; Narvhus and Gadaga, 2003).
The presence of yeasts indicates that they are able to proliferate during milk fermentation (Pereira-Dias et al., 2000; Suzzi et al., 2003; Romano et al., 2001) and positively interact with LAB (Gadaga et al., 2001; Narvhus and Gadaga, 2003). This implies that yeasts play either potentially beneficial or detrimental role to both the quality and safety of these milks (Jespersen, 2003; Gadaga et al., 2001; Lopandic et al., 2006; Pereira-Dias et al., 2000; Suzzi et al., 2003). Yeasts have been important as part of the starter cultures for development of sensory properties in milk products such as kefyr or koumiss (Fleet, 2006; Narvhus and Gadaga, 2003) and many types of cheeses (Lopandic et al., 2006).
Traditionally, the identification of yeasts has relied on phenotypic tests which are time-consuming, laborious and lack reproducibility and accuracy in results due to dependence upon physiological state and growth conditions of the yeasts (Lopandic et al., 2006; Fernandez-Espinar et al., 2006). Developments in molecular biology have however made it possible to overcome these limitations. Such methods include randomly amplified polymorphic DNA (RAPD), Pulsed Field Gel Electrophoresis (PFGE), Restriction Fragment Length Polymorphism (RFLP) and sequence analysis of specific genes. Restriction profiles from amplified 5.8S rRNA-ITS region digestion by endonucleases CfoI, HaeIII, HinfI and ScrfI (Esteve-Zarzoso et al., 1999) have been used to analyse clustered data and compare profiles with existing databases such as www.yeastid.com (Valencia University and CSIC, Spain). Using RAPD which employs single and short primers including both minisatellite and microsatellite DNA (Lieckfeld et al., 1993; Beh et al., 2006) enables accurate differentiation of closely related species. A consensus approach, polyphasic taxonomy, which integrates both phenotypic and genotypic methods for identifying and classifying foodborne yeasts, has been recommended (Lopandic et al., 2006; Arias et al., 2002).
Naturally fermented camel milk contributes largely to the nutrition of pastoralists living in the arid and semi-arid areas of East Africa (Farah et al., 1990). Here, about 10.5 million of the estimated world population of 24.2 million camels are located (FAO., 2007). The fermented milk, called suusac, is made by leaving the milk to ferment spontaneously at ambient temperature for about 24-48 h and yeasts have been reported to be part of the microbiota involved (Lore et al., 2005). A diverse consortium of yeasts might be involved in fermentations in these remote arid and semi-arid regions given the rudimental nature of suusac production. Milk handling and fermentation containers have been found to influence the diversity of yeasts and therefore characteristics of such spontaneously fermented milks (Kebede et al., 2007; Narvhus and Gadaga, 2003).
Classical tests have however mostly been used for identification of yeasts in African fermented milk (Vasdinyei and Deak, 2003). Little information exists on molecular techniques being applied for the identification of yeasts in African fermented milks and none on camel milk. The aim of this study was therefore to identify the yeast species present throughout the distribution process of camel milk and suusac.
MATERIALS AND METHODS
Total of 105 samples consisting of fermented camel milk, fermenting camel milk, raw camel milk and handling container swabs were collected between July and August of year 2007 and in May of year 2008. A laboratory suusac fermentation model was also carried out simulating the traditional process. Fermented or fermenting suusac samples were collected from Kenya in Isiolo, Nanyuki, Mandera and Garissa and Somalia in Burco and Garowe. Raw camel milk was sampled along two marketing chains, Nanyuki and Isiolo, at herd level as individual and pooled milk samples, first collection point and from the final market in Nairobi. Time taken between milking and first collection point varied from 4 to 7 h while it took at least 27 h between milking and final market. The pastoralists hold the milk at ambient temperatures normally between 25 and 35°C during transit due to lack of refrigeration facilities. In both locations, the traditional free ranging and intensive camel dairy herds were chosen for sampling. Samples were stored in dry ice to keep them frozen and transported to the laboratory within 8 h after collection.
Isolation and enumeration of yeasts: Appropriate dilutions of samples were surface plated on YM agar (Beckton Dickson AG) supplemented with chloramphenicol (20 μg mL-1). Viable yeast counts were enumerated after incubation at 25°C for 48 h and representative colonies selected based on their morphology. Three colonies per visual morphology-type were selected and purified by 3 times repetitive streaking. A total of 186 presumptive yeasts were isolated and transported frozen to Zurich. Characterization was done by catalase test (3% H2O2, VWR International), Gram-staining reactions (3% KOH, Sigma-Aldrich) and verification of morphology by microscopic examination. Isolates were then preserved in YM broth (Beckton Dickson AG) and 30% glycerol (Sigma-Aldrich Chemie GmbH, Buchs, Switzerland) at -80°C for use in subsequent experimentation.
Yeast reference strains: For analysis, presumptive yeast profiles were compared with those of reference strains Saccharomyces cerevisiae, Candida lusitaniae, Candida krusei and Rhodotorula mucolaginosa. These reference yeast strains belong to the current collection of microorganisms from the Laboratory of Food Biotechnology, ETH Zurich.
Identification with API 20C AUX: Identification of isolates by sugar assimilation was performed by use of API 20C AUX (bioMérieux, Geneva, Switzerland). API suspension medium ampoules (2 mL) were inoculated with single yeast colonies and the resulting suspension was standardized to turbidity equal to number 2 McFarland standards. Each cupule was inoculated with the standardised suspension and trays were incubated for either 48 or 72 h at 30°C. Cupules showing more turbidity than that of the negative control were considered positive. Identification was made by comparing the generated numerical profile using the API 20 C Analytical Profile Index.
DNA extraction: The method by He et al. (2005) was used to extract yeast DNA with modifications.
Briefly, the cells were collected from a fresh yeast colony and suspended in 50 μL of sterile double distilled water using sterile toothpicks. The mixture was heated at 95°C for 5 min after vortexing, then cooled down to 4°C and centrifuged at 15’000 g for 10 min. The supernatant was stored at -20°C till when needed.
A modified protocol by Yang et al. (2004) was used to for some isolates. A yeast colony was collected and suspended in 200 μL autoclaved water and 500 μL GPT reagent was added. GPT reagent constituents were 6 M guanidine thiocyanate (Fluka) dissolved in 50 mM Tris, pH 8.3, mixed with equal volume of phenol buffered in 10 mM Tris, pH 8.0 (Sigma-Aldrich). The sample was then vortexed and incubated at 95°C for 15 min followed by a 5-sec centrifugation step. Two hundred and fifty μL of 24:1 (v/v) chloroform-isoamyl alcohol (Sigma-Aldrich) was added, mixed and centrifuged at 140’000 rpm for 10 min. From the upper, aqueous phase, 450 μL was transferred to a fresh tube, mixed with an equal volume of isopropanol (Sigma-Aldrich) and kept for at least 1 h at -20°C. The mixture was centrifuged at 14’000 rpm for 15 min and the supernatant was carefully removed. The pellet was suspended in 500 μL of ice cold 70% ethanol prior to centrifugation for 5 min at 14’000 rpm and subsequently dried for 10 min at 700 mbar under vacuum (Univapo 150 ECH, Uniequip, Germany). The pellet was then resuspended in 25 μL of sterile double-distilled water and stored at -20°C.
Molecular typing
Restriction fragment length polymorphism: Intergenic spacers ITS1 and ITS2 were amplified using the PCR protocol described by Esteve-Zarzoso et al. (1999) with modifications as shown in Table 1. The 50-μL PCR mixture consisted of 2 μL DNA template and 48 μL PCR reaction mix composed of 22.5 μL autoclaved double distilled water, 0.5 μM of each of the primers (Table 1) and 25 μL of 2 x master mix (Fermentas) composed of 0.05 u μL-1 Taq DNA Polymerase, reaction buffer, 4 mM MgCl2 and 0.4 mM of each dNTP (dATP, dCTP, dGTP, dTTP). From each of the PCR products, 6 μL, were subjected to gel electrophoresis using 1.5% agarose gel in 1 x TAE (pH 8), stained with ethidium bromide and visualized under UV light (302 nm, AlphalmagerTM).
The remaining PCR products were purified using the GFX™ PCR purification kit (GE Healthcare Europe GmbH) prior to either restriction analysis or sequence analysis.
Restriction digestion was carried out using restriction endonucleases HhaI, HinfI and HaeIII (New England BioLabs, Bioconcept, Allschwil, Switzerland). The 22-μL aliquots contained 2.5 μL of 10x buffer (Bioconcept, Allschwil, Switzerland), 0.2 μL restriction enzyme and 7.5 μL double distilled water. Additionally, 100 μg mL-1 bovine serum albumin (Bioconcept, Allschwil, Switzerland) was included in the HhaI restriction mix. The mixtures were incubated at 37°C for 3 h. Electrophoresis was performed for 20 μL of the digests using a 3% agarose gel at 70 V for 100 min. Resulting fragment sizes were estimated as described in data analysis. The profiles were compared with those in the database at www.yeast-id.com (Valencia University and CSIC, Spain) and also those of reference strains. Where restriction profiles did not match those published in the database, sequence analysis of the PCR products was performed.
Sequence analysis of the 26S rRNA gene: For some yeast isolates, sequence analysis of the 26S rRNA gene was necessary and was performed with primers and amplification conditions shown in Table 1.
The PCR products were purified using the GFX™ PCR purification kit (GE Healthcare Europe GmbH) prior to sequencing at Microsynth (Balgach, Switzerland). Sequences were edited using the software Finch TV version 1.4.0 (Geospiza inc.) and aligned using BioEdit (Hall, 1999). Comparison with available databases was done by marching using the NCBI database accessible at http://blast.ncbi.nlm.nih.gov/Blast.cgi.
RAPD: To further differentiate the strains and for clustering some isolates prior to sequencing, RAPD was performed with (GTG)5, (GAC)5, (GACA)4 microsatellite primers and an M13 core sequence with primers and amplification conditions shown in Table 1. Amplification was done using a 25-μL reaction mixture containing 2 μL template DNA, 4% dimethyl sulfoxide (DMSO), 2 mM MgCl2 (Euroclone), 0.2 mM dNTPs mix (GE Healthcare Europe GmbH, Otelfingen, Switzerland) and 2.5 U Taq DNA polymerase (Euroclone). The PCR products were electrophoresed in aliquots of 15 μL using a 1.5% agarose gel and orange green loading dye.
| Table 1: | Oligonucleotide primers used and PCR conditionsa used for typing yeast isolates |
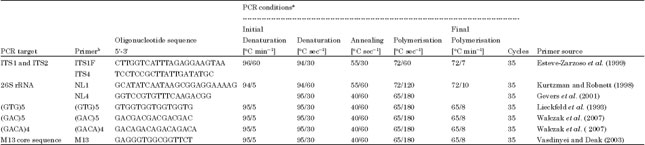 | |
| aPerformed in either Biometra® TGradient Cycler or Biometra® TPersonal Cycler, bSupplied by microsynth AG, Balgach, Switzerland | |
| Table 2: | Yeast species associated with camel milk and suusac from different sources and fermentation stages |
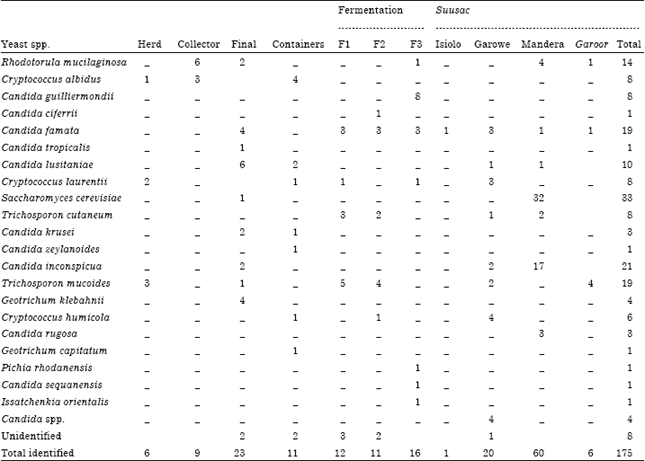 | |
| Herd: Pooled milk after milking; Collector : Milk from 1st collector; Final: Milk from the final market in Nairobi, F1, F2 and F3 means fermented after 23, 47 and 117 h, respectively, -: Indicate species not detected, garoor is a slightly overfermented suusac, numerals represent absolute numbers | |
Data analysis: Fingerprint data was analysed using the GelCompar II version 5.10 (Applied Maths, Sint-Martens-Latem, Belgium) software package. The similarity among digitized profiles was calculated using the Pearson correlation and an average linkage UPGMA (unweighted pair group method with arithmetic averages) based on dice coefficients. The digitized images were normalised to a TriDye 100-bp and 1-kb DNA standard (BioLabs New England, Bioconcept, Allschwil, Switzerland).
RESULTS
Yeasts counts along the market chain: There were very low numbers of yeasts in milk direct from the udder and after pooling from different camels (less than 300 cfu mL-1). At the first collection point after 3-8 h after milking, there were 4.4±1.4 log cfu mL-1 and at the final market more than 24 h after milking, counts went up to 5.0±1.5 log cfu mL-1 (Fig. 1).
Among the suusac samples from various sources, yeast counts varied between 2±1.7 and 5.4±0.5 log cfu mL-1 with suusac from Mandera having the highest number of yeasts (Fig. 2). Garoor, a variation of suusac where the product is allowed to further ferment, had 1.3 log cfu mL-1 higher yeast numbers than suusac both from same source, Garowe, Somalia. The highest yeast counts of 7.5±2.5 log cfu mL-1 were found in laboratory fermented suusac.
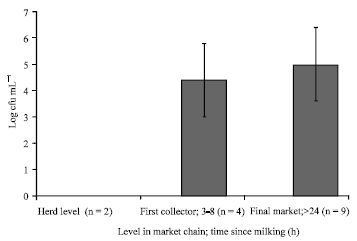 | |
| Fig. 1: | Yeast counts in raw camel milk along the market chain. n = number of samples. Vertical bars represent standard deviations |
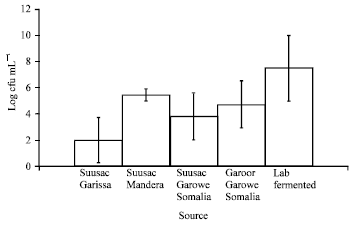 | |
| Fig. 2: | Yeast counts in fermented camel milk from different sources. Garoor is a form of over fermented suusac. Vertical bars represent standard deviations |
Identification of yeasts: Combined phenotypic and molecular approaches enabled the identification of 175 yeast isolates to genus and species levels (Table 2). There were 21 yeast species belonging to the genera Rhodotorula, Cryptococcus, Candida, Saccharomyces, Trichosporon, Geotrichum and Issatchenkia. The most frequently isolated yeasts included 33 Saccharomyces cerevisiae (19%), 21 Candida inconspicua (12%), 19 Trichosporon mucoides (11%), 19 Candida famata (11%), 14 Rhodotorula mucilaginosa (8%), 10 Candida lusitaniae (6%), 8 Cryptococcus laurentii (5%), 8 Cryptococcus albidus (5%), 8 Candida guilliermondii (5%) and 8 Trichosporon cutaneum (5%) (Table 2). Suusac from Mandera had the highest number of identified isolates, 60 (34% of all isolates) and Saccharomyces cerevisiae accounted for 32 (53.3%) of them. Additionally, milk from the final market at Nairobi accounted for 23 (13%), suusac from Garowe 20 (11%) and laboratory fermented camel milk (117 h) 16 (9 %) of all the identified isolates. Final market milk had highest diversity of the yeasts with 9 species isolated. Seven species were isolated from each of the containers, laboratory fermented milk (117 h) and suusac from both Mandera and Garowe (Table 2). Diversity of isolates increased for laboratory fermented camel milk with increase in incubation time from 23, 47 and 117 h after sampling with 4, 5 and 7 species identified, respectively (Table 2).
Lowest viable counts for the identified yeasts were 2.4 log cfu mL-1 for C. tropicalis and highest were 7.6 log cfu mL-1 (range; 2.4-8.5 log cfu mL-1) for C. famata and 8.0 log cfu mL-1 (range; 2.6-8.5 log cfu mL-1) for C. guilliermondii (Table 3).
| Table 3: | Yeast species identified using different methods and their viable counts |
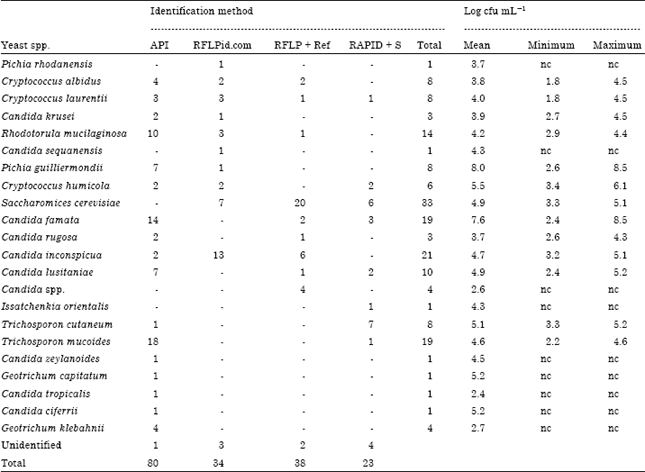 | |
| API: API 20C AUX; RFLPid.com means RFLP and comparison with database at www.yeastid.com; RFLP + Ref means RFLP analysis in comparison with reference strains; RAPD + S means RAPD then sequencing; - :Indicate species not typed conclusively using the method | |
API allowed the identification of 80 (47%) of the isolates to genus and species level at any of the identification classified as excellent, very good, good or acceptable identification (Table 3). Generated profiles of the other isolates were either unacceptable, unreliable or there were no matches for some isolates. However, 7 isolates belonging to the genera Candida, 2 Cryptococcus, 1 Geotrichum and 1 Saccharomyces were misidentified by API 20C AUX (Table 4). All except 2 of the misidentified isolates originated from fermented or fermenting camel milk.
Seventy three of the remaining isolates were identified using RFLP followed by either comparison of the profiles using the database at www.yeast-id.com or with those from reference strains (Table 3). The rest of the identified isolates were typed by sequencing of representative clusters obtained from RAPD profiles.
By clustering of fingerprints from RFLP and RAPD of the predominant isolates, RAPD revealed more variation than RFLP. For instance amongst a group of C. inconspicua, 8 cluster groups were generated using RAPD while 3 were generated using RFLP (Fig. 3a) and for R. mucilaginosa, 7 cluster groups were generated using RAPD while 6 were generated using RFLP (Fig. 3b). However for S. cerevisiae, 6 cluster groups were generated using RAPD while 7 were generated using RFLP (Fig. 3c).
| Table 4: | Yeast isolates not correctly identified by API 20C AUX but differently and conclusively identified using molecular methods |
 | |
| ID: Isolate study code; API = API 20C AUX; RFLPid.com means RFLP, then comparison with database at www.yeastid.com; RFLP + Ref means RFLP, then comparison with reference strains; RAPD+S means RAPD then sequencing | |
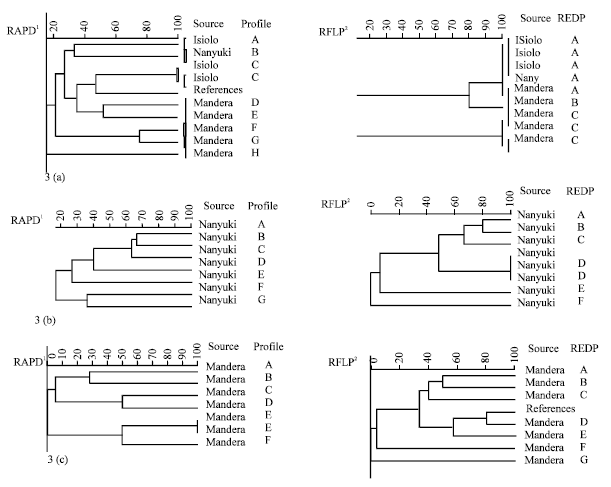 | |
| Fig. 3(a-c): | UPGMA dendrograms showing the relationships among predominant (a) C. inconspicua (b) R. mucilaginosa and (c) S. cerevisiae strains. 1RAPD calculated for four analysis with (GTG)5, (GAC)5, (GACA)4 microsatellite primers and M13 core sequence, 2RFLP by restriction analysis, ID and REDP stands for strain code and restriction endonuclease digestion profile respectively |
DISCUSSION
There exists little information about yeasts in camel milk and its products and no information applying molecular tools. A polyphasic approach was therefore used to identify the yeast species present throughout the distribution process of camel milk and suusac, a naturally fermented camel milk of East Africa.
We identified a diverse yeast consortium predominated by Saccharomyces cerevisiae, Candida inconspicua, Trichosporon mucoides, Candida famata, Rhodotorula mucilaginosa, Candida lusitaniae, Cryptococcus laurentii, Cryptococcus albidus, Candida guilliermondii and Trichosporon cutaneum. Similar yeast species have also been isolated and enumerated not only in milk (Kavas et al., 2006) but also in various other fermented foods and beverages such as cereal based (Omemu et al., 2007), cassava based (Omafuvbe et al., 2007) and rice based (Kofli and Dayaon, 2010). Saccharomyces cerevisiae which was the predominant yeast in our study has also been mentioned as the predominant yeast species in several African indigenous foods (Jespersen, 2003). Yeasts found in amasi a naturally fermented milk from Zimbabwe included S. cerevisiae and C. lusitaniae (Gadaga et al., 2007). In review on yeasts in naturally fermented milks in Zimbabwe and Uganda, predominant yeasts included Candida inconspicua (35%), C. kefir (20%), C. krusei (11%) and S. cerevisiae (10%). In the current study, Candida inconspicua (12%) and C. krusei (2%) were therefore less prevalent and S. cerevisiae (19%) more prevalent than in these Zimbabwean and Ugandan naturally fermented milks.
Viable yeasts increased with time from non-detectible at milking to 5.0±1.5 log cfu mL-1 in final market milk. Similar trends have been reported in Sudanese white cheese (El-Owni and Hamid, 2008; Idris and Alhassan, 2010), cooked raw salt white soft cheese (Abdalla and Mohammed, 2010), West African soft cheese (Adetunji and Chen, 2011), traditionally fermented peal millet (Hama et al., 2009) and corn based weaning foods (Omemu et al., 2007). In our study yeasts belonging to Candida spp. were found in highest numbers up to 7.6 log cfu mL-1 (range; 2.4-8.5 log cfu mL-1) and 8.0 log cfu mL-1 (range; 2.6-8.5 log cfu mL-1) for C. famata and C. guilliermondii respectively. Candida spp. has similarly been previously reported as the predominant yeast species growing up to log 6 cfu mL-1 in a review on African naturally fermented milks (Narvhus and Gadaga, 2003). The predominant yeast, S. cerevisiae, was enumerated at 4.9 log cfu mL-1 (range; 3.3-5.1 log cfu mL-1). Roostita and Fleet (1996) demonstrated growth of S. cerevisiae in UHT-treated cow milk up to 107-108 cfu mL-1 in 2-3 days.
Few studies report the sources of yeasts in milk and also other foods though it has been suggested that among the possible sources include chance contamination from environment and equipment used especially fermentation vessels (Narvhus and Gadaga, 2003). At milking, we detected very low yeast numbers. However, at the final market, counts of up to 6.5 log cfu mL-1 were detected. This indicates either the significant microbial build up or contamination. Several factors including unclean udder, milking with unclean hands, personal hygiene of milkmen, unclean (plastic) milking bucket, unclean milking site, pooling of fresh and old milk, prolonged exposure to high environmental temperatures, amongst other factors have been identified as potential sources of contaminants in camel milk (Younan, 2004). The differences in both the yeast numbers and diversity between the sampling sites also indicate that geographic factors could be as important and in some cases more than specific natural fermentation processes and hygienic practices. In a study of the effect of container type on the growth of yeast and lactic acid bacteria in sethemi, a South African traditional fermented milk there were no significant differences in the number of microorganisms between the different containers (Kebede et al., 2007). Differing results from those by Kebede et al. (2007) were found where there was a significant effect of packaging material on yeast counts in soft white cheese by Idris and Alhassan (2010). However like in our study, Kebede et al. (2007) report significant differences in both the dominant yeast species and the diversity of the yeast species.
Similar yeast species as isolated in this study have been reported to either play beneficial roles together with LAB in the fermentation processes or simply contribute to the spoilage of fermented milk. For instance, S. serevisiae, C. zeylanoids and various Pichia have been found to play beneficial role in several cheeses and fermented milks such as Kefir and Koumiss. S. cerevisiae has been associated with not only the production of alcohols and other aroma compounds but also stimulation of lactic acid bacteria, improvement of nutritional value and inhibition of undesired microorganisms (Jespersen, 2003). Roostita and Fleet (1996) found small amounts of ethanol (5 g L-1), lactic acid (1 mg mL-1 in 10 days) and propionic acid (0.7 mg mL-1) produced in milk by Saccharomyces cerevisiae. Zimbabwean traditionally fermented milk yeasts C. lusitaniae and S. cerevisiae enhanced growth of L. lactis subsp. lactis Lc261 and S. cerevisiae additionally stimulated growth of Lb. paracasei subsp. paracasei Lb11 in UHT milk (Gadaga et al., 2001). LAB from sugary kefir were also stimulated in 10 of 18 LAB and yeast combinations grown in glucose yeast extract (Leroi and Pidoux, 1993). The survival of and production of lactic acid by Lactobacillus hilgardii was enhanced by Saccharomyces florentinus (Leroi and Pidoux, 1993). C. humicola, C. krusei and Issatchenkia orientalis also contribute to fermentation of foods (Fleet, 2006). Yeasts may produce antimicrobial metabolites or bind bacteria on their surface therefore inhibiting or eliminating spoilage and/or potentially pathogenic microorganisms but they also have been reported to either inhibit the starter or support its function (Jakobsen and Narvhus, 1996). Similar yeast species including C. krusei, Cryptococcus neoformans, C. tropicalis and C. zeylanoids as identified in the present study were also isolated in cancer patients by Dorko et al. (2001) although causal connection was not established. We could however not establish such possibilities in our study and the yeasts need to be studied further for their role in camel milk.
In our study, API 20C AUX was only able to reliably identify 47% of the isolates. Likewise, in a study on yeasts in citrus juices, 33 and 42% of the isolates at the species and genus level, respectively were identified using RapID while 36% of the strains could not be identified and 21% were misidentified (Arias et al., 2002). API 20C AUX could identify only 12% of isolates to the species level with 82% of the isolates remaining unidentified (Arias et al., 2002). API 20C AUX in the present study therefore was able to identify higher percentage of isolates a fact that could be explained by difference in food type between the two studies.
Precise delineation of the importance of the yeasts based on accurate species identification is of great importance. When yeasts are involved in fermentations, the specific characteristics and quality of foods and beverages is to a great extent strain-specific (Beh et al., 2006). These strain specific characteristics are related to the fermentative and assimilative activity and production of various aromas by the yeasts (Lieckfeld et al., 1993). For instance several different isolates of S. cerevisiae involved in the fermentations show pheno- and genotypic characteristics that deviate from those normally recognised for S. cerevisiae (Jespersen, 2003). In analysis of yeasts in foods and beverages, the proposed sequence of operations is isolation, enumeration, identification to genus and species and strain differentiation (Beh et al., 2006). Conventional benchtop phenotypic characterization has been used in identification with limitations in precision and accuracy leading to poor resolution. Rapid identification kits API 20 C AUX and RapID Yeast Plus have been simplified the traditional methods. However, these yeast identification kits were initially designed for use in clinical diagnosis and their application is consequently confined to few yeast species (Arias et al., 2002; Kofli and Dayaon, 2010). The fact that the species found in milks are not present in the databases of commercial methods may also account for the low ability to correctly identify yeast isolates. However, new profiles from milk isolated yeasts could be added to the database as has been done previously with yeasts from orange juices (Arias et al., 2002). This would make these simple and rapid methods more robust and suitable for use in the food industry. Differences in identification accuracy between commercial kits have also been noted (Kofli and Dayaon, 2010).
In our study, RFLP and sequencing of the of ITS1-5.8S rRNA-ITS2 region with primers ITS1 and ITS4 and restriction enzymes CfoI, HaeIII, HinfI not only enabled the successful identification of 72 isolates where API 20 C AUX could not but also correct identification of misidentified isolates. Molecular techniques, such as those used for the current work, with high discriminatory ability exploiting various molecular targets have been published. RFLP of the ITS1-5.8S rRNA-ITS2 with several different restriction enzymes for identification of food associated yeasts has been used in several studies. However in many cases choice of the restriction enzymes will affect the size of the PCR products and the restriction patterns obtained. We noted unique profiles from each of the endonucleases CfoI, HaeIII and HinfI for each species. Similar observations have been made by others (Esteve-Zarzoso et al., 1999). In similar studies, a quite heterogeneous population of S. cerevisiae isolated from table olives was shown when restriction of amplified 5.8S rRNA-ITSs analysis was performed with RFLP with CfoI, HaeIII, HinfI and ScrfI (Arroyo-Lopez et al., 2006). It therefore requires a combination of enzymes to enable complete unequivocal identification. For instance due to the diversity and complexity of Candida spp. three restriction enzymes CfoI, HaeIII, HinfI are required for reliable identification. These 3 restriction enzymes were also recommended as effective for the identification of the genera Rhodotorula, Cryptococcus, Pichia and Candida (Esteve-Zarzoso et al., 1999) which we also typed in the present study.
When fingerprints from RFLP and RAPD of the predominant isolates were clustered, RAPD enabled further distinction of strains amongst the predominant species except for S. cerevisiae, where RFLP yielded more differentiation. In agreement with our results, RAPD has been used by others to reveal polymorphisms that differentiate at both the species and strain levels (Beh et al., 2006; Walczak et al., 2007). For instance RAPD enabled identification and differentiation of S. pustoriunus, S. williunus and S. buyanus (competitors of S. cerevisiae during fermentation), which have many times been termed synonyms of S. cerevisiae based on similarity in morphological and physiological properties (Barnett et al., 1990). The level of resolution however varies with the taxa under study and the primers used (Beh et al., 2006). Among the species we identified in this study, Cryptococcus neoformans, Cryptococcus albidus, Cryptococcus laurentii, Rhodotorula rubra and strains of Cryptococcus neoformans have in other studies been differentiated effectively using primers (GTG)5, (GACA)4 and M13 (Lieckfeld et al., 1993; Beh et al., 2006). Lieckfeldt et al. (1993) differentiated 23 biotechnologically important strains of the yeast Saccharomyces cerevisiae at strain level using RAPD probes and primers (GTG)5, (GACA)4 phage MI3 DNA and the M13 core sequence.
CONCLUSIONS
The successful integration of different techniques in the identification of yeasts in our work emphasise the importance of a polyphasic approach as the optimal way for proper yeast identification. The various data facilitate generation of consensus identification for each isolate where the limitations of each technique are overcome and the reliability of the identification improves. New profiles from milk isolated yeasts should also be added to the database of commercial kits to make these simple and rapid methods more robust and suitable for use in the food industry. The diversity and high numbers of yeasts indicate that they have a role either positive or detrimental in the fermentation of camel milk. As a prerequisite to their inclusion as starter cultures, it will therefore be important to investigate further the interaction of such yeasts with LAB and also their metabolic properties.
ACKNOWLEDGMENTS
This project was funded by the North-South Centre ETH. The authors thank Dr. Zakariah Farah (ETH Zurich), Dr. Mario Younan (Kenya Agricultural Research Institute), Dr. Esther Schelling (Swiss Tropical Institute), PD Dr. Jakob Zinstag (Swiss Tropical Institute), Dr. Chris Field (Kenya Camel Association), Dasel Mulwa (University of Nairobi), Monika Weller (ETH Zurich) and Swiss Federal Scholarship Commision (ESKAS) for their invaluable personal and institutional support and assistance.
REFERENCES
- Abdalla, M.O.M. and E.H.S. Mohammed, 2010. Effect of storage period on the microbiological and sensory characteristics of cooked low salt white soft cheese (Gebna Beyda). Pak. J. Nutr., 9: 205-208.
CrossRef - Adetunji, V.O. and J. Chen, 2011. Effect of temperature and modified vacuum packaging on microbial quality of wara a West African soft cheese. Res. J. Microbiol., 6: 402-409.
CrossRefDirect Link - Arias, R.C., J.K. Burns, L.M. Friedrich, M.R. Goodrich and E.M. Paricsh, 2002. Yeast species associated with orange juice: Evaluation of different identification methods. Applied Environ. Microbiol., 68: 1955-1961.
Direct Link - Arroyo-Lopez, F.N., M.C. Duran-Quintana, J.L. Ruiz-Barba, A. Querol and A. Garrido-Fernandez, 2006. Use of molecular methods for the identification of yeast associated with table olives. Food Microbiol., 23: 791-796.
CrossRef - Dorko, E., M. Kmetova, E. Pilipcinec, F. Dorko, I. Bracokova, A. Marossy and I. Skardova, 2001. Non-albicans Candida species in cancer patients. J. Medical Sci., 1: 11-15.
CrossRefDirect Link - El Owni, O.A.O. and O.I.A. Hamid, 2008. Effect of storage period on weight loss, chemical composition, microbiological and sensory characteristics of sudanese white cheese (Gibna bayda). Pak. J. Nutr., 7: 75-80.
CrossRefDirect Link - Esteve-Zarzoso, B., C. Belloch, F. Uruburu and A. Querol, 1999. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers Int. J. Syst. Bacteriol., 49: 329-337.
CrossRef - Farah, Z., T. Streiff and M.R. Bachmann, 1990. Preparation and consumer acceptability tests of fermented camel milk in Kenya. J. Dairy Res., 57: 281-283.
CrossRef - Fleet, G.H., 2007. Yeasts in foods and beverages: Impact on product quality and safety. Curr. Opin. Biotechnol., 18: 170-175.
CrossRef - Gadaga, T.H., A.N. Mutukumira and J.A. Narvhus, 2000. Enumeration and identification of yeasts isolated from Zimbabwean traditional fermented milk. Int. Dairy J., 10: 459-466.
CrossRefDirect Link - Gadaga, T.H., A.N. Mutukumira and J.A. Narvhus, 2001. The growth and interaction of yeasts and lactic acid bacteria isolated from Zimbabwean naturally fermented milk in UHT milk. Int. J. Food Microbiol., 68: 21-32.
CrossRef - Gevers, D., G. Huys and J. Swings, 2001. Applicability of rep-PCR fingerprinting for identification of Lactobacillus spieces. FEMS Microbiol. Lett., 205: 31-36.
CrossRef - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hama, F., A. Savadogo, C.A.T. Ouattara and A.S. Traore, 2009. Biochemical, microbial and processing study of Degue a fermented food from Pearl millet dough) from Burkina Faso. Pak. J. Nutr., 8: 759-764.
CrossRefDirect Link - He, R., Q. Wang, Y. Zhang, Q.Y. Liu and G. Li et al., 2005. Rapid and efficient generation of PCR templates from Escherichia coli, Saccharomyces cerevisiae and Oryza sativa using a microwave and by boiling. J. Rapid Meth. Autom. Microbiol., 13: 19-28.
CrossRef - Idris, Y.M.A. and I.H. Alhassan, 2010. Effect of packaging material on microbiological properties of sudanese white cheese. Int. J. Dairy Sci., 53: 128-134.
CrossRefDirect Link - Jakobsen, M. and J. Narvhus, 1996. Yeasts and their possible beneficial and negative effects on the quality of dairy products. Int. Dairy J., 60: 755-768.
CrossRefDirect Link - Jespersen, L., 2003. Occurrence and taxonomic characteristics of strains of Saccharomyces cerevisiae predominant in African indigenous fermented foods and beverages. FEMS Yeast Res., 3: 191-200.
CrossRefDirect Link - Kavas, G., O. Kinik, H. Uysal, S. Kilic, N. Celikel and N. Akbulut, 2006. Characterisation of yeasts isolated from artisanal Turkish dairy products. Int. J. Dairy. Sci., 1: 44-50.
CrossRefDirect Link - Kebede, A., B.C. Viljoen, T.H. Gadaga, J.A. Narvhus and A. Lourens-Hattingh, 2007. The effect of container type on the growth of yeast and lactic acid bacteria during production of Sethemi, South African spontaneously fermented milk. Food Res. Int., 40: 33-38.
CrossRef - Kofli, N.T. and S.H.M. Dayaon, 2010. Identification of microorganism from ragi for bioethanol production by API kit. J. Applied Sci., 10: 2751-2753.
CrossRefDirect Link - Kurtzman, C.P. and C.J. Robnett, 1998. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek, 73: 331-371.
CrossRefPubMedDirect Link - Leroi, F. and M. Pidoux, 1993. Detection of interactions between yeasts and lactic acid bacteria isolated from sugary kefir grains. J. Appl. Bacteriol., 74: 48-53.
CrossRef - Lieckfeld, E., W. Meyer and T. Borner, 1993. Rapid identification and differentiation of yeasts by DNA and PCR fingerprinting. J. Basic Microbiol., 33: 413-425.
PubMed - Lopandic, K., S. Zelger, L.K. Banszky, F. Eliskases-Lechner and H. Prillinger, 2006. Identification of yeasts associated with milk products using traditional and molecular techniques. Food Microbiol., 23: 341-350.
CrossRef - Lore, T.A., S.K. Mbugua and J. Wangoh, 2005. Enumeration and identification of microflora in suusac, a Kenyan traditional fermented camel milk product. LWT-Food Sci. Technol., 38: 125-130.
CrossRefDirect Link - Omafuvbe, B.O., A.R. Adigun, J.L. Ogunsuyi and A.M. Asunmo, 2007. Microbial diversity in ready-to-eat fufu and lafun-fermented cassava products sold in Ile-Ife, Nigeria. Res. J. Microbiol., 2: 831-837.
CrossRefDirect Link - Omemu, A.M., M.O. Bankole, O.B. Oyewole and A.K. Akintokun, 2007. Yeasts and moulds associated with ogi-a cereal based weaning food during storage. Res. J. Microbiol., 2: 141-148.
CrossRefDirect Link - Pereira-Dias, S., M.E. Potes, A. Marinho, M. Malfeito-Ferreira and V. Loureiro, 2000. Characterisation of yeast flora isolated from an artisanal Portuguese ewes' cheese. Int. J. Food Microbiol., 60: 55-63.
CrossRef - Romano, P., A. Ricciardi, G. Salzano and G. Suzzi, 2001. Yeasts from Water Buffalo Mozzarella, a traditional cheese of the Mediterranean area. Int. J. Food Microbiol., 69: 45-51.
CrossRef - Roostita, R. and G.H. Fleet, 1996. Growth of yeasts in milk and associated changes to milk composition. Int. J. Food Microbiol., 31: 205-219.
CrossRefDirect Link - Suzzi, G., M. Schirone, M. Martuscelli, M. Gatti, M.E. Fornasari and E. Neviani, 2003. Yeasts associated with Manteca. FEMS Yeast Res., 3: 159-166.
CrossRef - Vasdinyei, R. and T. Deak, 2003. Characterization of yeast isolates originating from Hungarian dairy products using traditional and molecular identification techniques. Int. J. Food Microbiol., 86: 123-170.
CrossRef - Yang, Z.Y., W.B. Shim, J.H. Kim, S.J. Park, S.J. Kang, B.S. Nam and D.H. Chung, 2004. Detection of aflatoxin-producing molds in Korean fermented foods and grains by multiplex PCR. J. Food Prot., 67: 2622-2626.
PubMed