
H.R.M. Galal
Department of Botany, Faculty of Science at Qena, South Valley University, 83523, Egypt
W.M. Salem
Department of Botany, Faculty of Science at Qena, South Valley University, 83523, Egypt
F. Nasr El-Deen
Department of Botany, Faculty of Science at Qena, South Valley University, 83523, Egypt
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 8 | Page No.: 645-657







ABSTRACT
The present research has been conducted to explore the antifungal potency of the methanol and ethyl acetate crude extracts of seaweeds collected from the red sea, Hurghada, Egypt. The concentration of (50 mg mL-1) was tested against Alternaria alternata, Fusarium oxysporium, Alternaria brassicicola, Ulocladium botrytis and Botryotrichum Piluliferum. The antifungal activities were expressed as inhibition in dry mass (mg), protein content (μg mL-1) and enzymatic activity of pectinase and cellulase (units mL-1). For comparative study, the biological activity of standard antibiotic Nystatin was also measured. Crude ethyl acetate extract of Padina gymnospora and methanolic extract of codium fragile exhibited strong activity against most of the tested fungi. All the tested fungi were sensitive to Nestatin, except F. oxysporium (the most resistant fungi), where the dry weight recorded from 68 to 69%, protein recorded 69 to 52%, pectinase and cellulase activity 62 to 58% of control. The most sensitive fungi were U. botrytis where the dry weight, protein content, pectinase and cellulase activities all were completely inhibited in cellulose and pectin media. While, the most active algae is the ethyl acetate extract of P. gymnospora and methanolic extract of C. fragile. inhibited pectinase and cellulase enzymes activities for all the tested fungi except A. brassicicola and F. oxysporium. This report confirms the broad antifungal effect of C. fragile using the methanolic extracts rather than ethyl acetate (P. gymnospora). Additionally, These Egyptian seaweeds, therefore, considered as a potential source for treating infections caused by the tested plant fungi.
PDF Abstract XML References Citation
Received: March 02, 2011;
Accepted: July 18, 2011;
Published: September 05, 2011
How to cite this article
H.R.M. Galal, W.M. Salem and F. Nasr El-Deen, 2011. Biological Control of Some Pathogenic Fungi using Marine Algae Extracts. Research Journal of Microbiology, 6: 645-657.
URL: https://scialert.net/abstract/?doi=jm.2011.645.657
URL: https://scialert.net/abstract/?doi=jm.2011.645.657
INTRODUCTION
The orientations for treatment of plant diseases are currently interesting in extraction of natural products as alternatives to synthetic fungicides for their safety and negligible environmental impacts (Brimmer and Boland, 2003). For example, Khalil et al. (2005) and Zafar et al. (2002) reported some medicinal plants as antifungal activities against plant pathogenic fungi. Osman et al. (2011) examined the antifungal activity of some agriculture wastes (rice straw, maize and cotton wastes) against Rhizoctonia solani with successful results. Also, seaweeds considered a possible and easy source of antimicrobial compounds due to their variety of secondary metabolites with antifungal activities (Cordeiro et al., 2006). Pathogenic fungi are the most organisms responsible for a considerable plant yield losses than other microorganisms (Sexton and Howlett, 2006). Extracted substances from seaweeds confirmed earlier for their antifungal (Khanzada et al., 2007) and antibacterial activities (Salem et al., 2011). The crude extracts obtained from herbs essential oils, Cassia fistula and Mesua ferrea are subjected to broad biological screening for antifungal activity (Sohel and Yeasmin, 2004; Mousavi et al., 2009). Nowadays, extraction of new compounds from macro algae confirmed its biocidal activity as antifungal agents (Bhosale et al., 2002). Such compounds are extracted from different macroalgae families, like green, brown and red algae (Vallinayagam et al., 2009). Tuney et al. (2006) recorded positive results for methanol, acetone, diethyl ether and ethanol extracts of Cystoseira mediterranea and Ulva rigida, for their antifungal activities. According to prior reports, it has been confirmed that most of biologically active compounds can be used as therapeutic agent that are found in seaweeds (Madhusudan et al., 2011).
However, the reports on the antifungal activities of seaweed extracts from Egypt are very limited. Hence, this investigation aimed to screen and evaluate the efficiency of organic solvents such as methanol and ethyl acetate extracts from the Egyptian seaweeds as antifungal agents against some plant pathogenic fungi.
MATERIALS AND METHODS
Algae collection and extract preparation: Eight marine algae were collected by hand picking from the red sea in Hurghada, Egypt during June 2009. Algal samples were cleaned from epiphytes, extraneous matter and necrotic were removed. Samples were collected in sterilized polyethylene bags and put in an ice box, then transferred to the laboratory immediately. The collected algae comprise three different families as follows: Phaeophyceae (Cystoesira myrica, Cystoesira trinodis, Padina gymnospora, Sargassum dentifolium, Sargassum hystrix); Rhodophyceae (Actinotrichia fragilis) and Bryopsidophyaceae (Codium fragile). Samples were washed thoroughly with sea water then sterile distilled water, air dried, cut into small pieces and then ground in a tissue grinder (IKA A 10, Germany) until reach fine powder shape.
The dried samples (10 g) were extracted in two different solvents: (100 mL of methyl alcohol or ethyl acetate) under stirring condition (50 rpm) for 7 days at room temperature. The solution was filtered through Whatman No. 1 sterile filter paper. The filtrates then were dried using desiccator (Cole- parmer instrument, Chicago). The dried precipitates were dissolved in the above two solvents to give 50 mg mL-1 extracts, then stored in airtight bottles in a refrigerator before testing. These crude extracts were screened against common pathogenic fungi as follows:
Fungal source and culture conditions: Alternaria alternata, Fusarium oxysporium, Alternaria brassicicola, Ulocladium botrytis and Botryotrichum Piluliferum were isolated from infected seeds, identified and subcultured on glucose-Czapek's agar medium (Smith and Dawson, 1944). All the fungal isolates were screened for their abilities to produce extracellular cell wall-degrading enzymes such as pectinase according to Hankin et al. (1971) and cellulase according to Eggins and Pugh (1962).
Effect of algal extracts on pectin lyase production: The fungi were grown in Erlenmeyer flasks (100 mL) containing 50 mL of sterilized liquid media, inoculated with a single 6 mm disc cut-out from the margin of a 5 day colony of each fungus grown on glucose-Czapek,s agar medium. The flasks were supplemented with 1% of the algal extracts and incubated at 28±1°C for 7 days. Methanol and ethyl acetate was used as a negative control while nystatin used for comparative study.
After 7 days of incubation, the mycelial growth, PL activity and extracellular protein content were determined. The mycelial growth was then filtered through pre-weighted 0.45 μ filter paper using a filter unit. The agar inoculums were removed from the mycelium. The dry weight of the mycelium was left to dry in an oven at 85°C for 24 h. Filtrates were centrifuged at 15.000 g for 15 min at 4°C for assaying of enzymes activities and extracellular of protein content.
PL assay procedures: The PL assay procedures were carried out according to Sherwood (1966) because there is a correlation between virulence of the pathogens and their ability to degrade cellulose and pectin.
Protein content determination: Protein of culture filtrates (as indicators for fungi activities) was determined according to Lowry et al. (1951).
Effect of algal extracts on cellulase (exo-1,4-β-glucanase) production: The method described by Nelson (1944) and modified by Naguib (1964) was employed for cellulose determination.
Statistical analysis: The data of all experiments were subjected to analysis by the Least Significant Differences test (L.S.D) using PC-STATE program version 1A, coded by Rao, M.; Blane, K. and Zannenberg, M, University of Georgia.
RESULTS
The effect of marine algal extracts on dry mass, protein and extracellular enzyme functions of different fungi are presented in Table 1 to 6.
The data showed that all the tested fungi were sensitive to nestatin, except F. oxysporium where the dry weight recorded 68% of absolute control in pectin medium (Table 1). The most sensitive fungi were U. botrytis where the dry weight completely inhibited in both cellulose and pectin media. While, the most active algae, P. gymnospora (ethyl acetate extract) where dry weight of all tested fungi was inhibited completely, the exception was A. brassicicola (68.6% dry weight compared to control) (Table 1). Methanolic extracts showed much more bioactivity (inhibition of fungal dry weight) than ethyl acetate extracts. For example, the ethyl acetate extracts of C. fragile, A. fragilis and C. trinodes did not show any antifungal activity (indicated as dry weight) against all of the tested fungi (Table 1). C. fragile (methanolic extract) also showed high antifungal activity with all tested fungi, except F. oxysporium (58% dry weight compared to control) (Table 2).
U. botrytis and A. brassicicola are the most sensitive fungi (showed complete inhibition of protein content) when ethyl acetate of and methanolic extracts were employed, respectively (Table 3). While, the most active algae, P. gymnospora (ethyl acetate extract) where protein content of all tested fungi was inhibited completely, the exception was A. brassicicola (61.6% protein content compared to control) (Table 3). C. fragile (methanolic extract) also showed high antifungal activity with all tested fungi, except F. oxysporium (69.88%) protein content compared to control (Table 4). Both ethyl acetate of (C. fragile, A. fragilis and C. trinodes) and methanolic extracts of (S. dentifolium, C. myrica and C. trinodes) showed non antifungal activity (based on protein content) for all tested fungi (Table 4).
For antifungal assay by pectinase and cellulase enzyme activities (units mL-1), U. botrytis with ethyl acetate extracts and A. brassicicola with methanolic extracts, were the most sensitive fungi (did not show any enzymes function activity) (Table 5).
| Table 1: | Antifungal assay by dry weight (mg) in pectinase medium |
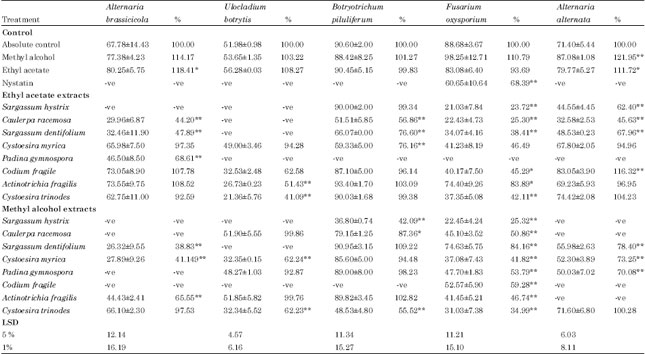 | |
| Mean±SD, n = 3. *Significant differences, **Highly significant differences, -ve: Negative results (complete inhibition). | |
| Table 2: | Antifungal assay by dry weight (mg) in cellulase medium |
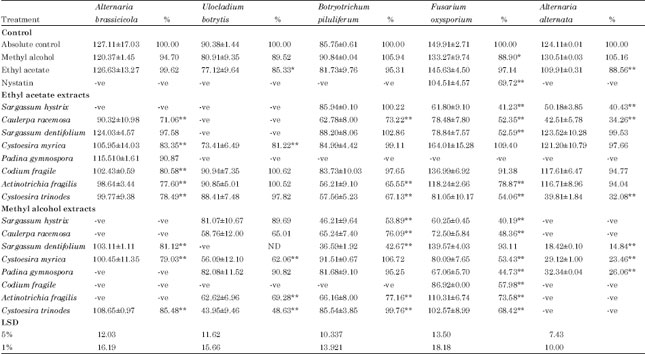 | |
| Mean±SD, n = 3. *Significant differences, ** Highly significant differences, ND: Not determined, -ve: Negative results (complete inhibition) | |
| Table 3: | Antifungal assay by protein content (µg mL-1) in pectinase medium |
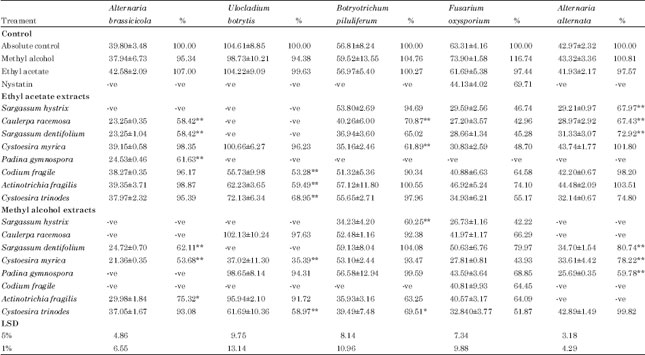 | |
| Mean±SD, n = 3.*Significant differences, **Highly significant differences, -ve: Negative results (complete inhibition) | |
| Table 4: | Antifungal assay by protein content (µg mL-1) in cellulase medium |
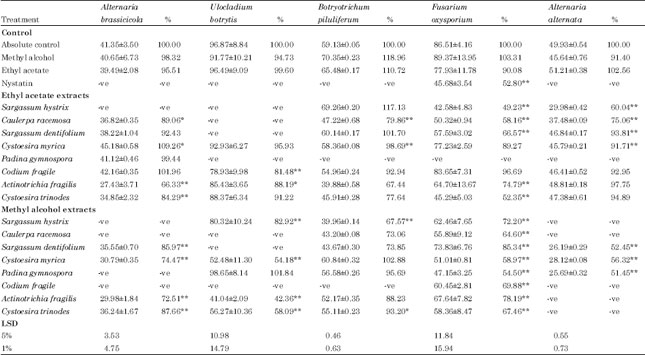 | |
| Mean±SD, n = 3. *Significant differences, **Highly significant differences, -ve: Negative results (complete inhibition) | |
| Table 5: | Antifungal assay by pectinase enzyme activity (units mL-1) in liquid medium |
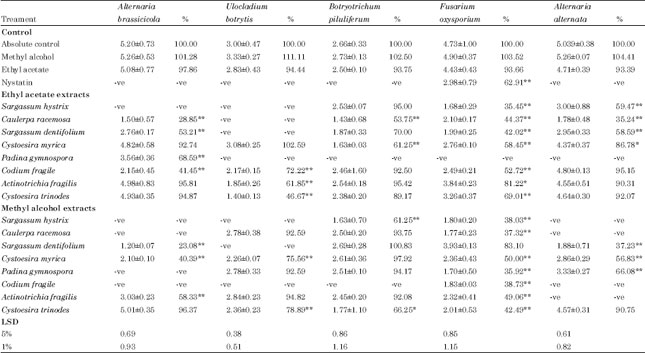 | |
| Mean±SD, n = 3, *Significant differences, **Highly significant differences, -ve: Negative results (complete inhibition) | |
| Table 6: | Antifungal assay by cellulase enzyme activity (units mL-1) in liquid medium |
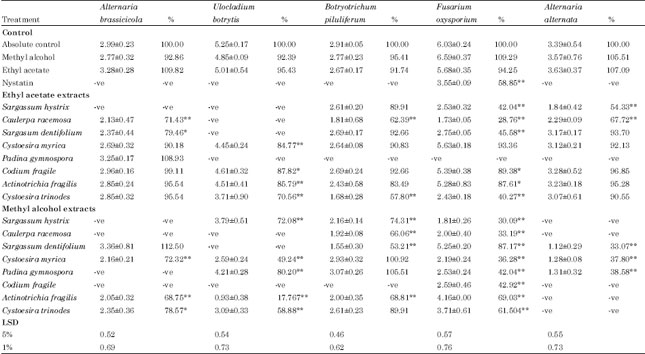 | |
| Mean±SD, n = 3. *Significant differences, **Highly significant differences, -ve: Negative results (complete inhibition) | |
F. oxysporium is the most resistance fungus, whereas the fungicide nystatin did not inhibit its pectinase and cellulase activities (growth percentage was 62.9% as compared to control) (Table 5). On the other hand, the most active algae were, P. gymnospora (ethyl acetate extract) where pectinase and cellulase enzymes activity was inhibited completely all the tested fungi, the exception was A. brassicicola (68.5%) as compared to control (Table 5).
C. fragile (methanolic extract) also showed high antifungal activity with all tested fungi, except F. oxysporium (42.9% compared to control) of cellulase enzyme activities (Table 6). Both ethyl acetate of (C. fragile, A. fragilis and C. trinodes) and methanolic extracts of C. myrica showed non antifungal activity (based on pectinase and cellulase enzymes activities) for all tested fungi (Table 6).
DISCUSSION
Biological natural products isolated earlier from seaweeds are recommended as potential biocidal and pharmaceutical agents (Rangaiah et al., 2010). Broad ranges of data are now available for the in vitro anti-fungal activities of different seaweeds families (Tuney et al., 2006). The results obtained recorded the higher antifungal activity for ethyl acetate extracts of P. gymnospora and methanolic extracts of C. fragile. Then, the methanolic and ethyl acetate extracts of S. hystrix as indicated by inhibition in dry weight, protein content, pectinase and cellulase enzymes activities. The phenolic compound released from dried crudes of seaweed extracts may be answerable for their antimicrobial properties. This was confirmed earlier by Cox et al. (2010) who found that phenolic compounds responsible for the antifungal activities of seaweeds. Other scientists have also reported that phenolic compounds from different plant sources could inhibit various food-borne pathogens (Plaza et al., 2009; Osman et al., 2011). This may be due to the impact of these antifungal compounds on spore germination (El-Mehalawy, 2003). The above results may also be due to the effect of these antifungal on cell wall shifting its permeability (Wen-Bao et al., 2000) or the antifungal suppressed the early stages of mycelial growth (El-Mehalawy, 2003).
In pectin and cellulose medium, there was a linear relationship between protein amount released to the medium and fungal enzymes activities. Obviously, some algal extracts was able to inhibit the activity of pectinase only while cellulase enzyme exhibit some activity for the same fungus. For example, methanolic extract of S. hystrix was able to inhibit pectinase enzyme but had no effect on cellulase enzyme of U. botrytis (Table 5, 6). The explanation of this case was confirmed earlier by Karthikaidevi et al. (2009). This may explain the specific target (receptors) of the antimicrobial compounds of these algal extracts against one enzyme. This result could be related to the presence of bioactive metabolites present in this species of algae which are not soluble in one solvent but they can be soluble in the other.
It is worth mentioning that, the efficiency of extraction of antifungal natural products from marine macroalgae was higher with methanol as confirmed earlier with the obtained results. Manilal et al. (2009) tested several solvents and found that methanol was determined to be the best solvent for isolation of bioactive secondary metabolites from dried red algae followed by ethyl acetate and dichloromethane. These results indicated that extraction method had definite effects on the isolation of bioactive principles. The effectiveness of extraction methods reported by many authors showed that methanol extraction yielded higher antimicrobial activity than n-hexane and ethyl acetate (Paul and Puglisi, 2004). Also, this ensures the importance of selection the appropriate solvent and extraction method to be used. Some extracts seemed to be specific in their activity against some pathogens, which ensures the presence of specific bioactive compounds against certain targets.
Pectinases functions were employed for its important role for fungal virulence and help in the infection process (Jones et al., 1972). Since, they corrupt pectin subunits of the middle lamella and of primary cell wall, support the colonization of plant tissues (Favaron et al., 1994). Consequently, it supplying nutrients for the fungus through the early stages of infection (Deo and Shastri, 2003). The results obtained showed that pectinase activity significantly decreased when P. gymnospora and C. fragile extracts were employed. A wide range of results of in vitro anti-fungal activities of extracts of green algae, diatoms and dinoflagellates have been reported to affect fungal enzymes activities (Tuney et al., 2006). Another important explanation provided by Cordeiro et al. (2006), who approved that chitinasses and β-1,3-glucanase (that can affect pectinase function) are truly recognized as natural antifungal proteins widely found in plants and seaweeds.
The results also reported that crude P. gymnospora and C. fragile extracts (50 mg mL-1) within pectin and cellulose media showed the same effectiveness compared to fungicide nystatin. And among all the seaweeds, P. gymnospora is considered as the ideal antifungal macroalga tested.
CONCLUSION
From cellulase and pectinase tests, we can conclude that, maximum antifungal activities was recorded in Bryopsidophyaceae (C. fragilis) and Phaeophyaceae (P. gymnospora) which was the most effective seaweeds and minimum antifungal activities were recorded for C. myrica a member of the Phaeophyaceae.
REFERENCES
- Bhosale, S.H., V.L. Nagle and T.G. Jagtap, 2002. Antifouling potential of some marine organisms from India species of Bacillus and Pseudomonas. Mar. Biotechnol., 4: 111-118.
PubMed - Brimmer, T. and G.J. Boland, 2003. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ., 100: 3-16.
CrossRefDirect Link - Cordeiro, R.A., V.M. Gomes, A.F.U. Carvalho and V.M.M. Melo, 2006. Effect of proteins from the red seaweed Hypnea musciformis (wulfen) Lamouroux on the growth of human pathogen yeasts. Braz. Arch. Biol. Technol., 49: 915-921.
Direct Link - Cox, S., N. Abu-Ghannam and S. Gupta, 2010. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J., 17: 205-220.
CrossRefDirect Link - Deo, A. and N.V. Shastri, 2003. Purification and characterization of polygalacturonase-inhibitory proteins from Psidium guajava Linn. (guava) fruit. Plant Sci., 164: 147-156.
CrossRef - Eggins, H. and P.J.F. Pugh, 1962. Isolation of cellulose decomposing fungi from soil. Nature, 193: 94-95.
CrossRefDirect Link - Favaron, F., R. D'Ovidio, E. Porceddu and P. Alghisi, 1994. Purification and molecurlar characterization of a soybean PGIP. Planta, 195: 80-87.
PubMed - Hankin, L., M. Zucker and D.C. Sands, 1971. Improved solid medium for detection and enumeration of pectolytic bacteria. Appl. Microbiol., 22: 205-209.
Direct Link - Zafar, I., S. Mussarat, H. Farrakh, B. Sheraz, I. Mohammad, Z. Shahida and A. Bashir, 2002. Antifungal properties of some indigenous plants from Peshawar valley. Asian J. Plant Sci., 1: 708-709.
CrossRefDirect Link - Karthikaidevi, G., K. Manivannan, G. Thirumaran, P. Anantharaman and T. Balasubaramanian, 2009. Antibacterial properties of selected green seaweeds from vedalai coastal waters: Gulf of mannar marine biosphere reserve. Global J. Pharmacol., 3: 107-112.
Direct Link - Khalil, A.B., B.F. Dabaneh and G.H. Anfoka, 2005. Antifungal activity of medicinal plants from Jordan environment. Plant Pathol. J., 4: 130-132.
CrossRefDirect Link - Khanzada, A.K., W. Shaikh, T.G. Kazi, S. Kabir and S. Soofia, 2007. Antifungal activity, elemental analysis and determination of total protein of seaweed, Solieria robusta (Greville) kylin from the coast of Karachi. Pak. J. Bot., 39: 931-937.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Madhusudan, C., S. Manoj, K. Rahul and C.M. Rishi, 2011. Seaweeds: A diet with nutritional, medicinal and industrial value. Res. J. Med. Plant, 5: 153-157.
CrossRefDirect Link - Manilal, A., S. Sujith, J. Selvin, C. Shakir and G.S. Kiran, 2009. Antibacterial activity of Falkenbergia hillebrandii (Born) from the Indian coast against human pathogens. Phyton, 78: 161-166.
Direct Link - Mousavi, S.M., S.S. Mirzargar, H.E.Z. Mousavi, R.O. Baigi, A. Khosravi, A. Bahonar and M.R. Ahmadi, 2009. Evaluation of Antifungal activity of new combined essential oils in comparison with malachite green on hatching rate in rainbow trout (Oncorhynchus mykiss) eggs. J. Fish. Aquat. Sci., 4: 103-110.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Osman, M.E.H., M.M. El-Sheekh, M.A. Metwally, A.A. Ismail and M.M. Ismail, 2011. Efficacy of some agriculture wastes in controlling root rot of Glycine max L. induced by Rhizoctonia solani. Asian J. Plant Pathol., 5: 16-27.
CrossRefDirect Link - Paul, V.J. and M.P. Puglisi, 2004. Chemical mediation of interactions among marine organisms. Nat. Pro. Rep., 21: 189-209.
PubMed - Plaza, M., S. Santoyo, L. Jaime, G.G.B. Reina, M. Herrero, F.J. Senorans and E. Ibanez, 2010. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal., 51: 450-455.
CrossRefPubMedDirect Link - Sexton, A.C. and B.J. Howlett, 2006. Parallels in fungal pathogenesis on plant and animal hosts. Eukaryotic Cell, 5: 1941-1949.
CrossRef - Sohel, F.I. and M.S. Yeasmin, 2004. Antimicrobial screening of Cassia fistula and Mesua ferrea. J. Med. Sci., 4: 24-29.
CrossRefDirect Link - Tuney, I., B.H. Cadirci, D. Unal and A. Sukatar, 2006. Antimicrobial activities of the extracts of marine algae from the coast of Urla (Izmir, Turkey). Turk. J. Biol., 30: 171-175.
Direct Link - Wen-Bao, C., H. Yuh-Felling, C.J. Shung and C.C. Sheno, 2000. Isolation, purification and characterization of killer protein from Schwanniomy cesoccidentalis. Appl. Environ. Microbiol., 66: 5348-5352.
Direct Link