
Rachel Fernandez
Physiology, Nutrition and Pathology Division, Central Marine Fisheries Research Institute, Kerala 682014, India
Manpal Sridhar
Physiology, Nutrition and Pathology Division, Central Marine Fisheries Research Institute, Kerala 682014, India
N. Sridhar
Physiology, Nutrition and Pathology Division, Central Marine Fisheries Research Institute, Kerala 682014, India
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 466-479







ABSTRACT
To evaluate the nutritional quality of LAB and their possible effect on growth and survival of juvenile Penaeus indicus four commercially available cultures viz., L. acidophilus, S. cremoris, L. bulgaricus-56 and L. bulgaricus-57 were orally administered via a moist feed base (cell concentration 5x106 cells g-1 of diet) to four treatment groups in three replicates and designated as D1, D2, D3 and D4 at 12% of the body weight for a period of 35 days. A control group devoid of bacteria designated DO served as control. The diets contained 55% protein and 4.25% crude fat. All the treatment groups recorded higher weight as compared to the weight gain of 21.32% recorded in control group of shrimp (p<0.05). Specific growth rates and survival rates were also better in all the treatment groups as compared to control (p<0.05). Feed D2 recorded the lowest and best FCR of 0.64 as compared to 3.28 recorded on the control diet. The same was the case with PER, K1 and K2 showing S. cremoris to be the most efficient nutritionally. The probiont incorporated diets recorded higher digestibilities ranging between 93.98 to 82.15% as compared to 70.46% obtained on the control diet (p<0.05). Total Plate Count of the gut microflora revealed localization of LAB. The present investigation clearly demonstrates the beneficial role of LAB probionts orally monitored via compounded feed on growth in P. indicus juveniles.
PDF Abstract XML References Citation
Received: December 08, 2010;
Accepted: March 08, 2011;
Published: May 21, 2011
How to cite this article
Rachel Fernandez, Manpal Sridhar and N. Sridhar, 2011. Effect of Lactic Acid Bacteria Administered Orally on Growth Performance of Penaeus indicus (H. Milne Edwards) Juveniles. Research Journal of Microbiology, 6: 466-479.
URL: https://scialert.net/abstract/?doi=jm.2011.466.479
URL: https://scialert.net/abstract/?doi=jm.2011.466.479
INTRODUCTION
During the last few decades, aquaculture has become the worlds fastest growing food production sector, with cultured shrimp growing at an annual rate of 16.8%. Meanwhile, according to a World Bank report, global losses resulting from shrimp diseases are around 3 billion US dollars. The potential negative consequences of using antibiotics in aquaculture, such as the development of drug resistant bacteria and the reduced efficiency of antibiotic resistant for human and animal diseases, have led to suggestions of the use of nonpathogenic bacteria as probiotic control agents (Vaseeharan and Ramasamy, 2003).
Single cell proteins offer many advantages over conventional animal and plant proteins. They have a high protein content (40-80% protein on dry weight basis) along with a very short generation time. They can be easily cultivated in limited land space over available carbon sources. It is possible to control their nutritional composition as they can be genetically manipulated (Tacon, 1990). The use of microbial probiotics in aquaculture is now widely accepted (Mohanty et al., 1993; Mohanty et al., 1996; Gatesoupe, 1999; Sharma and Bhukhar, 2000; Gomez-Gil et al., 2000; Verschuere et al., 2000; Irianto and Austin, 2002; Wang and Zirong, 2006; Vine et al., 2006; Wang, 2007). Probionts have been used to feed live feed and larvae of finfish and shellfish directly as well as indirectly and their use has produced some positive results. The use of Probionts as feed additives has been preferred over that of antibiotics. They do not exhibit any undesirable effects associated in the use of antibiotics. Most studies carried out in larvae failed to record toxicity or allergy. Bacterial drug resistance and indiscriminate suppression of intestinal micro flora are not observed with the use of probiotics. Also residues which remain in food like in the case of antibiotics are not obtained.
The nutritional merits of Lactic acid bacteria as probiotic have been proven in humans and a number of other terrestrial organisms. Many studies using animal models have clearly demonstrated a protective effect of dietary supplements of lactic acid bacteria against colon tumor development (Kabir et al., 2004). Gursoy and Kinik (2006) showed probiotics supplementation promoted significant influence on live weight gain, high carcass yield, prominent cut up meat parts and immune response in day old broiler chicks.
Silver-Ps a probiotic applied in culture ponds of Peneaus monodon yielded higher body weights, total production and survival rates compared with control ponds (without probiotics). The shrimps in control ponds were seriously affected by bacterial infection; gill soaking and tail rot diseases while these problems were not encountered in probiotics applied ponds showing that probiotics play a vital role in growth, survival and production of the shrimps by maintaining good water quality parameters through out the culture period (Soundarapandian and Sankar, 2008). The experimental introduction of Lactic Acid Bacteria (LAB) into the intestine of fish has been reviewed (Ringa and Gatesoupe, 1998). Several studies have shown that it is possible to maintain artificially the LAB population at high levels by regular intake through feed especially in cod, salmon and turbot. Adhesion is acknowledged as the first step of a microorganism in the process of colonization and the intestinal mucous plays a vital role in this process. There are reports in endothermic animals that there is a certain degree of host-specificity in the adhesion process. Adhesion to intestinal mucous has also been assayed in vitro in fish (Joborn et al., 1997). Intestinal bacteria of turbot adhere specifically to intestinal mucous than to any control surface. Specific adhesions have been demonstrated in the adhesion of yeasts to intestinal cell walls of rainbow trout (Vazquez-Juarez et al., 1997). Similar studies on crustaceans in general and commercially important shrimp in particular are wanting. The present study was thus taken up with the main objective of evaluating the effect of four commercially available LAB fed orally via a compounded feed on growth and survival of juvenile Penaeus indicus.
MATERIALS AND METHODS
Microorganisms: For the present study four commonly utilized LAB strains viz., Lactobacillus acidophilus NCIM 2285, Streptococcus cremoris NCIM 2285, Lactobacillus bulgaricus NCIM 2285 (2056) and Lactobacillus bulgaricus NCIM 2285 (2057 ). Hansens strain obtained from the National Collection of Industrial Micro-organisms ( NCIM ) National Chemical Laboratory, Pune, India were used. They were revived on NCIM enrichment media containing glucose, lactose, liver extract yeast extract and salts (pH 7.6) and maintained as slant cultures on MRS agar and sub cultured every week. The authenticity of the cultures was verified by sub culturing and identification based on their morphological and biochemical characteristics (Holt, 1989). Pure cultures were stored at 4°C in agar slants with sub culturing every 3-4 weeks. Pure culture of Vibrio alginolyticus obtained from the collection maintained at the Microbiology Laboratory, Center for Fish disease diagnosis and management, School of Environmental studies, Cochin University of Science and Technology was used as the test organism. The microbes were sub cultured every week and stored at 4°C as TSA slants containing 1.0% sodium chloride.
Production of bacterial biomass: L. acidophilus, S. cremoris, L. bulgaricus-56 and L. bulgaricus-57 were grown aseptically in 10 mL of nutrient broth for 24 h at room temperature (28±2°C ). Five millilitre of log phase culture was then transferred under aseptic conditions into 250 mL of MRS broth and placed on a rotary shaker at 150 rpm for 24 h at 28±2°C. The cells of each strain were harvested separately by centrifuging at 10.000 rpm under aseptic conditions for 15 min at 4°C. The supernatants were saved and used for testing in vitro the antagonistic property of Vibro while the accumulated cell biomass was dispensed in 5 mL of PBS in individual sterile glass vials and used for incorporating in feed.
Feed preparation: A moist compounded feed base was prepared fresh every week as per the specification of New (1989). A pre-weighed quantity of freshly extracted fish oil (sardine) procured from the local extraction mill was added to each individual LAB biomass. The whole was thoroughly remixed in a vortex mixer . Each was separately incorporated into feed to give a final concentration of 5x10°cells/g of diet. The control feed contained only PBS and oil. The feeds were stored at 4°C and viability of the bacterial cells determined at regular intervals of 24 h by standard plate count using MRS agar.
Feeding experiments: Juveniles of Penaeus indicus averaging 0.905±0.23 g in weight and 6.27±0.12 cm in length were procured from the ponds of the Central Institute of Brackishwater Aquaculture, Narakkal. They were transported to the laboratory in oxygen bags and acclimatized for a week in 500 L capacity fiber glass tanks supplied with aeration at 28±2°C prior to commencement of the experiment. The shrimp were randomly segregated into fifteen groups so that three replicates were available for each of the four treatments and the control groups. The animals were maintained in plastic tubs containing 40 L of filtered seawater irradiated with ultra violet light. They were maintained on a pelleted diet fed at 12% of body weight per day in two divided doses at 10 and 16 h, respectively. The physico-chemical parameters of the rearing water were monitored and averaged at 32±2‰ for salinity; 4.5±0.5 mg L-1 for dissolved oxygen; 8±0.5 for pH and 28±1°C for temperature during the entire experimental duration. The feeding experiment was carried out for 35 days from September to October at the wet laboratory of the Institute after which the animals were sacrificed and gut dissected out under aseptic conditions. Serial dilutions were carried out and plated onto Tryptone Soya Agar (TSA) plates containing 1.0% sodium chloride and incubated at 28±2°C and developed colonies identified.
Analytical methods: Proximate composition of the feed ingredients and experimental feeds was evaluated as per AOAC (1990). Total Plate Count (TPC) of gut micro flora, feed and water was carried out as per standard methods (APHA, 1980). Analysis of Variance (ANOVA) was carried out for all data. All analysis was carried out in three replicates.
RESULTS
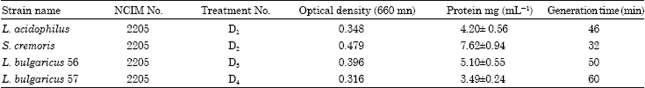
The four LAB cultures were checked for their growth and protein yield prior to incorporation into feeds. S. cremoris had the shortest doubling time of 32 min while L. bulgaricus 57 had the longest generation time of 60 min (Table 1).
| Table 1: | Growth and yield of the four LAB culture used in the feeding experiments |
 | |
| Table 2: | The biochemical characteristics of the lactic acid bacteria (LAB) used for feeding experiments with P. indicus juveniles |
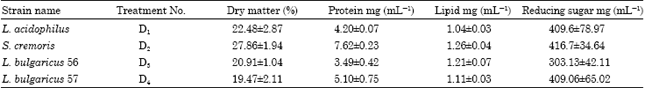 | |
The biochemical characteristics of these four LAB strains, used for biomass accumulation for the feeding trials with P. indicus juveniles are given in Table 2. S. cremoris recorded the highest protein content of 7.62 mg mL-1 followed by L. bulgaricus-57 with 5.10 mg mL-1 where as L. bulqaricus-56 showed the lowest protein content of 3.49 mg mL-1 amongst the four LAB strains. Dry matter content of the four strains ranged from 27.86% (S. cremoris) to 19.47% (L. bulgaricus-57). Lipid content did not show much variation between strains and ranged from 1.26 in S. cremoris to 1.04 mg mL-1 in L. acidophilus. A reducing sugar content of 409.6, 416.7 and 409.06 mg mL-1, respectively was obtained for L. acidophilus, S. cremoris and L. bulgaricus-57 while L. bulgaricus-56 recorded comparatively lower reducing sugar content of 303.13 mg mL-1.
Proximate composition of feed ingredients and feed: All the selected ingredients were subjected to proximate composition analysis before formulation of the feeds (Table 3). The control feed DO was devoid of bacterial biomass had a crude protein content of 55.0%. Feeds D1, D2, D3 and D4 had the four LAB cultures incorporated individually and their protein content varied from 55.04 to 55.08%. All the feeds had a lipid content averaging 4%. The other nutrients of these feeds were well within the limits fulfilling the nutritional requirements of shrimp (Table 4).
Growth performance of P. indicus juveniles: The treatment groups showed a statistically significant increase in length and weight (Table 5) in comparison to control (p<0.05). Maximum increase in length (0.58 cm) was recorded in D2 the group fed with the biomass of S. cremoris and the least of 0.27 cm in D1 the group fed with L. acidophilus biomass. The control group DO recorded an increase of only 0.18 cm which was the lowest as compared to the treatment groups. Correspondingly the highest increase in length of 9.35% was obtained in D2 and lowest of 4.37% in treatment D. Increase in weight was maximum (1.741 g) in group D2 the animal fed biomass of S. cremoris while lowest weight gain (1.250 g) was obtained in group D3 the L. bulgaricus-56 fed animals. (Table 5). The control group of shrimp showed an increase in weight of 1.195 g-1 corresponding to an increase of 21.32%.
As also evident from Table 5 the highest percentage weight gain of 86.80% was obtained in D2 followed by 48.37% in group D4 (p<0.05). Significant differences were also observed between the specific growth rate and survival of the treatment groups and the control. Maximum SGR of 0.023 and 0.014 was observed in groups D2 and D4, respectively. Highest survival of 98 was obtained in group D2 followed by 95% in groups D1 and D4 while in the control group it was 70% (p<0.05).
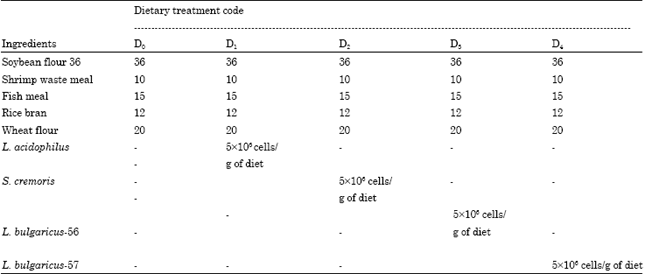
| Table 3: | Details of the feeding treatments and ingredients inclusion levels adopted in the feeding experiment with P. indicus juveniles |
 | |
| Table 4: | Proximate chemical composition of the experimental (moist) and control feeds used for the feeding experiments with P. indicus juveniles |
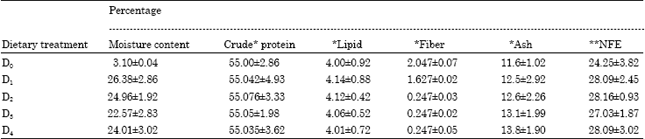 | |
| * % Dry matter basis, *NFE-nitrogen free extractives calculated as (100- % crude protein + crude lipid + ash + crude fiber + moisture) | |
| Table 5: | Growth performance of P. indicus juveniles reared on the experimental and control feeds for four weeks |
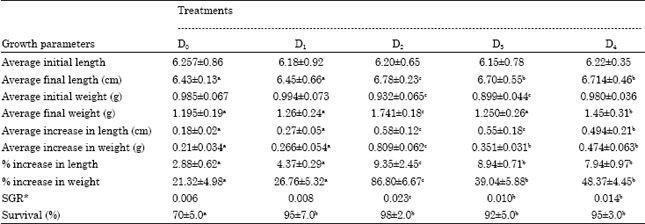 | |
| *SGR-Specific growth rate. Row means with same superscripts are not significantly different (p>0.05) | |
Amongst the experimental feeds feed D2 incorporated with S. cremoris gave the lowest and best FCR of 0.64 followed by 1.66 for feed D4 (L. bulgaricus-57), 1.99 for feed D3 (L. bulgaricus-56) and 2.64 in case of feed D1 (L. acidophilus), respectively.
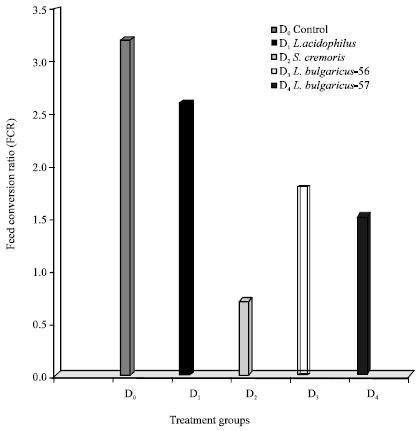 | |
| Fig. 1: | Effect of LAB probionts on feed conversion ratios |
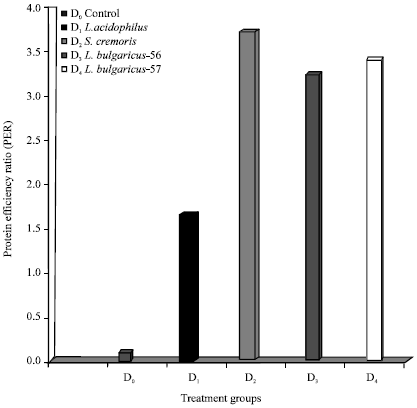 | |
| Fig. 2: | Effect of LAB probionts on protein efficiency ratios (PER) |
The control feed, though being dry and pelleted gave a much higher (Poor) FCR of 3.1 in comparison to the experimental feeds (Fig. 1). The Protein Efficiency Ratios of the experimental and control feeds elicited a similar pattern obtained in the case of Feed conversion Ratios.
| Table 6: | Apparent total digestibility (%) of the control and probiont incorporated feeds in P.indicus juveniles |
 | |
| Column means with same superscripts are not significantly different (p>0.05) | |
| Table 7: | Changes in the nutritional quality of the control and treatment groups after feeding probiotic incorporated feeds for four weeks |
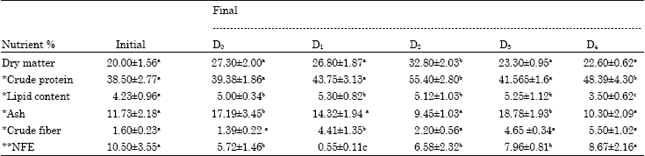 | |
| *% Dry matter basis. **NFE-Nitrogen free extractives calculated as (100-% crude protein+crude lipid+ash+ crudefiber+moisture). Row means with same superscripts are not significantly different (p>0.05) | |
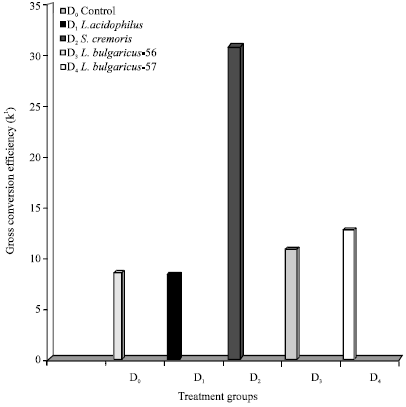 | |
| Fig. 3: | Effect of LAB probionts on gross conversion efficiency (K1) |
Feed D2 gave the best PER of 3.8 followed by D4 and D3 with values of 3.47 and 2.37, respectively. Comparatively lower PER of 1.69 was obtained in feed D1. The control feed D0 gave a very low PER of 0.08 (Fig. 2). The highest K1 value of 32 was obtained for D2 and lowest value of 7.6 for feed D1 (Fig. 3).
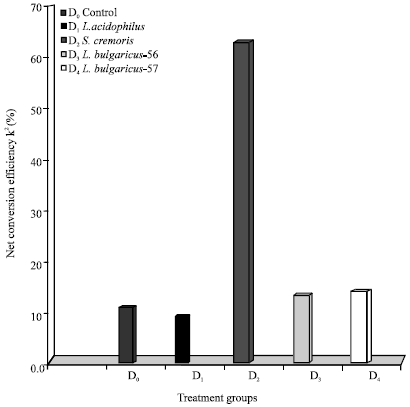 | |
| Fig. 4: | Effect of LAB probionts on net conversion efficiency (K2) |
Feed D4 gave a K1 value of 12.8 and feed D3 a value of 10.1. Again the lowest gross conversion efficiency value of 89 was obtained in the control feed D0. The net conversion efficiency values (Fig. 4) showed the maximum conversion efficiency of 65.25 was obtained with feed D2. Feeds D1, D2 and D3 gave comparatively poor conversion efficiencies of 13.9, 11.8 and 18.1, respectively. A conversion efficiency of 12.19 was obtained on the control feed. The apparent total digestibility of the probiont incorporated feeds in P. indicus juveniles given in Table 6 showed the probiont incorporated feeds to give higher and better digestibility ranging from 93.98% for feed D2 to 90.68% for feed D1. Digestibility obtained for feed D3 (88.98%) and feed D4 (82.15%) were also higher than 70.46% the total digestibility obtained for the control feed D0 (p<0.05).
Changes in the nutritional quality of the control and LAB fed P. indicus: An increase was recorded in the dry matter content in the carcasses of all the treatment groups as compared to the initial dry matter content of the shrimp at the start of the experiment (Table 7). Crude carcass protein increased in all the treatment groups upon feeding LAB from 38.5% the value recorded at the start of the experiment to 39.38% in the control group (D0). The highest increase of 55.4% was obtained for D2 group fed with S. cremoris biomass followed by 48.39% for D4, 43.75% for D1 and 41.505% for D3, respectively. Lipid content recorded an increase from the initial 3.5 to 4.23% for the control group and 5, 5.3, 5.12 and 5.25% for the treatment groups D1, D2, D3 and D4, respectively. Ash content of the animals did not show any particular trend as the control group D0 and treatment groups D3 and D1 showed higher values of 17.19, 18.78 and 14.32%, respectively as compared to the initial value of 11.73% at the start of the experiment. Comparatively low values of 9.00 and 10.3% were recorded for the ash content in treatments D2 and D4. Though the fiber and carbohydrate contents of the carcasses recorded mild variations but these were however not significant.
Total plate counts (TPC) of sea water and gut micro flora: The sea water used in the animal experiments was subjected to TPC. Flexibacter spp. Enterobactereaceae spp. and Pseudomonas spp. formed the dominant bacterial flora in the sea water before subjecting it to sterilization using ultraviolet radiation. The TPC of the gut micro flora is presented in Table 8. The predominance of LAB was observed in all treated groups.
DISCUSSION
The use of probionts as feed additives has been preferred over that of antibiotics as they do not exhibit any of the undesirable effects associated in the use of antibiotics viz., toxicity, allergy, residues in food, bacterial drug resistance and indiscriminate suppression of intestinal micro flora. Metchnikoff (1907) worked with Bulgarian bacillus, an organism closely related to lactobacillus starter of yogurt (L. delbreuckii subspecies bulgaricus) and to this day Lactobacilli have remained the most commonly used probiotic organisms.
Several mechanisms have been investigated whereby bacteria could function as a probiotic. These include adhesion to digestive tract wall to prevent colonization of pathogens (competitive attachment) neutralization of toxins, bactericidal activity and increased immune competence.
Fish are continuously exposed to a wide range of microorganisms present in the environment and the micro biota of fish has been the subject of several reviews. Moriarty (1998) evaluated lactic acid bacteria in fish and demonstrated that Streptococcus, Leuconostoc, Lactobacillus and Carnobacterium belonged to the normal micro biota of the gastrointestinal tract in healthy fish. However, it is well known that the population level of lactic acid bacteria associated with the digestive tract is affected by nutritional and environmental factors like dietary polyunsaturated fatty acids, chromic oxide, stress and salinity. Pathogenic lactic acid bacteria such as Streptococcus, Enterococcus, Lactobacillus, Carnobacterium and Lactococcus have been detected from ascites, kidney, liver, heart and spleen. Some antibiotic treatments and vaccinations have been proposed to cure or prevent these diseases that seem however, to spread with the development of fish culture. It has also been reported that some lactic acid bacteria isolated from the gastrointestinal tract of fish can act as probiotics. These candidates are able to colonize the gut and act antagonistic against Gram-negative fish pathogens. These harmLess bacteriocin-producing strains may reduce the need to use antibiotics in future aquaculture (Piard and Desmazeaud, 1992).
Production of lactic acid by lactic acid bacteria reduces the pH of the stomach contents in endothermic animals. Studies in vitro have shown that an acid condition of less than pH 4.5 prevents the growth of many bacteria including coli forms but still allows the growth of some strains of lactobacilli (Sissons, 1989). LAB are also known to produce hydrogen peroxide which has bactericidal actions in vitro and produces a metabolite thought to neutralize the effect of enter toxin released from coli forms.
| Table 8: | Bacterial flora of gut wall in the control and experimental animals at the end of feeding trial. The bacterial counts are given as colony forming units (CFU)/shrimp |
 | |
Clearly LAB is the best studied among all probionts, even so the effect of its use in crustaceans has not been studied. Although LAB is not the dominant gut micro flora in marine fish as compared to endothermic animals, researchers have been able to introduce LAB into larval and juvenile fish with pronounced protection against Vibro infections. As in the case of fish a strain of Vibro alginolyticus is used as probiotic agent for larval rearing of Penaeus vannamei in Ecuador. Use of this strain increased survival of P. vannamei post-larvae by competitive exclusion of pathogenic virblos and reduced antibiotic prophylaxis in intensive larval culture systems (Griffith, 1995), Uma et al. (1999) reported that the growth and survival of P. indicus juveniles were significantly improved by the addition of Lacto-saccTM (a commercial livestock probiotic feed supplement composed of Saccharomyces sp. Lactobacillus acidophilus and Streptococcus faecium) at levels ranging from 2.5 to 7.5 g kg-1 basal feed. A challenge with Vibro alginolyticus resulted in low mortality rate in Lacto-saccTM fed animals than in control group. However, the presence of these organisms in the gut of the shrimp after feeding was not ascertained. Sridhar and Paulraj (2003) introduced strains of Bacillus and Micrococcus isolated from shrimp guts by coating them onto compounded diets into P. indicus post-larvae. They observed significantly higher specific growth rates and survival in treatment groups than control. Up to 106 CFU/shrimp of probiotic organisms was detected in the gut of the post-larvae. The results of the present study are also in agreement where 65 to 80% colonization of LAB was obtained in the gut due to oral feeding. However, the permanency of these LAB cultures in the gut should be ascertained by estimating their numbers after stopping the oral feeding.
The improvement of the production rate of rotifers was observed with Adjulact, a spray -dried whey with two strains of live lactic bacteria (S. thermophilus and L. helveticus). As the heat killing of the probiotic increased further the rotifer production rate, it was assumed that this effect was due to a growth factor(Gatesoupe et al., 1989a). This growth factor may have come from L. helveticus, since the whey containing S. thermophilus as the only live bacteria did not improve the production (Gatesoupe, 1991a). In the same study the efficiency of L.plantarum was found to be higher than that of L. helveticus. Acosil a spray dried extract from sprouting cereal grain, fermented with selected strains of lactic acid bacteria, limited the bacterial proliferation in rotifers during their overnight enrichment with fish oil emulsion (Gatesoupe et al., 1989b) Adjulact with L. plantarum decreased the bacterial counts in rotifers, especially the count of the dominant Vibrionaceae (A. salmonicida). The spores of Bacillus IP5832 (Paciflor) did not decrease the bacterial counts but the proportion of Vibrionaceae dropped dramatically while a very diversified flora set up. Therefore, the effect on the rotifer flora depends on the probiotic but there is a general trend to reduce the bacterial counts, at least those of Vibrionaceae (Gatesoupe, 1991b). On the other hand no further improvement was observed when probiotics were given to Artemia nauplli .The improvement of the rotifer production and/or the improvement of their bacterial environment may account for the increase of the growth rate of larval fish.
The most promising result of probiotics use was reported in the improvement of the survival rate of turbot due to rotifers fed with spores of Bacillus (Gatesoupe, 1993). This improvement was observed in the event of an infection with A. hydrophila whether the infection was casual or experimental. As the spores are fast ingested then digested by rotifers, the hypothesis was that a Vibrostatic substance from Bacillus hinders the proliferation of Vibrionaceae in rotifers and thereby in turbot. The advantage of this treatment over antibiotics was that it worked at a very low concentration (about 20 spores per rotifer, daily) and the medium also was not affected by any drug.
Metaillier and Hollocou (1993) fed triplicate lots of young 30 g European sea bass (Dicentrarchus labrax) three different diets each including a different probiotic ad libitum. They were compared with lots fed on a standard diet. Food consumption was quite the same for all the lots. When compared with the standard, growth was slightly improved (5%) for two diets but not significantly. Nutritional parameters obtained with probiotics were not different from the standard a finding contradictory to the results of the present study. They concluded that only through a new study carried out on younger animals, during weaning period for example, would it be possible to point out a potential probiotic effect for this species.
Bacillus S11 bacterium isolated from black tiger shrimp habitats was added to shrimp feed as a probiotic in three forms: Fresh cells, fresh cells in normal saline solution and a lyophilized form (Rengpipat et al., 1998). After a 100-day feeding trial with probiotic supplemented and non-supplemented (control) feeds, Penaeus monodon (from PL30) exhibited no significant difference (p>0.05) in growth, survival nor external appearance between all three probiotic treatments but significant differences (p<0.05) occurred between probiotic and control groups also a finding in agreement with the results of our study. After challenging shrimps with a shrimp pathogen, Vibro harveyi, by immersion for 10 days, all probiotic treatment groups had 100% survival whereas the control group had only 26% survival. In addition, the control group had an unhealthy external appearance and deformed texture of the hepatopancreas and intestine, while treatment group shrimp appeared healthy and normal. In another experiment that was performed by Rengpipat et al. (2003), the growth and resistance to Vibro in black tiger shrimp (P. monodon) fed with a Bacillus probiotic (BS11) were studied. As reported by us in the present study they found that the growth and survival rates of shrimps fed on the probiotic supplement were significantly greater than those of the controls.
Seven putative probionts isolated from the gut were mixed with an equal volume of a lipid oralizer and incorporated into compounded feed pellets to achieve a dose of 5x10 bacterial cells g-1 of fish food and fed to rainbow trout Oncorhynchus mykiss fingerlings for a period of 25 days (Sridhar et al., 2006). Superior growth and feed efficiencies were obtained in animals fed the probiotic feeds clearly demonstrating the beneficial effects of orally administered probionts in enhancing growth of culture organisms as well as in increasing their resistance to diseases. The present study also shows that commercial cultures of LAB accorded superior growth and digestibility upon oral administration in shrimp larvae.
Isolated LAB was used as a probiotic in fresh water fish tilapia (Oreochromis mossambicus) against the most common fish pathogen Aeromonashy drophila (Vijayabaskar and Somasundaram, 2008). Higher antagonistic activity recorded from Extra Cellular Protein (ECP) or bacteriocin compared to the Intra Cellular Protein (ICP) against A. hydrophila. After feeding with the potential probiotics for 25 days, challenge by immersion indicated effectiveness at reducing disease caused by A. hydrophilain fishes. Tilapia exhibited no significant difference in growth, survival nor external appearance between the probiotic fed treatments but significant differences (p<0.05) occurred between probiotic and control groups. Askarian et al. (2008) analyzed the composition of Lactic Acid Bacteria (LAB) in intestine of two species of sturgeon, beluga (Huso huso) and Persian sturgeon (Acipenser persicus) and observed that the LAB population levels in the two sturgeon species was not similar and in beluga it was significantly higher than Persian sturgeon. Two species of LAB including Enterococcus seriolicida and Leuconostoc mesenteroides were isolated from GI tract of Persian sturgeon and the predominant species was L. mesenteroide. Furthermore, Lactobacillus curvatus, Lactococcus raffinolactis, Lactococcus lactisandStreptococcus sp. were isolated from the GI tract of beluga and the counts of L. curvatuswas significantly higher in the GI tract of beluga than other species.
Wang (2007) reported Daily Gain (DWG) and Relative Gain Rate (RGR) in all groups of Penaeus vannamei treated with probiotics at all concentrations to be significantly higher than those of the Control. However, mean values of DWG and RGR were not significantly different among treatment groups. These results show that probiotics composed of equal weight of photosynthetic bacteria and Bacillus sp. increased growth performance in shrimps. The results of our study also clearly demonstrate the positive role of LAB orally monitored via compounded feed on growth in P. indicus juveniles. It is thus possible to seed the gut with harm Less bacteria which occupy the attachment sites and prevent infection by pathogenic bacteria by maintaining artificially these bacterial populations at high level by regular intake with feed. Such treatments would be beneficial to improve health and quality of shrimp in culture without the use of antibiotics.
CONCLUSION
Probiotics have been successfully used in aquaculture to enhance both internal and external microbial environments. The role of beneficial bacteria to limit and to control environmental pathogens will become particularly important in the future of aquaculture, especially with regard to the increasing number of antibiotic resistant strains of bacteria, the tightening of governmental and environmental regulations of treatments and the cost-effectiveness. The use of probiotics in aquaculture is now an acceptable practice and is on the increase. The present investigation clearly demonstrates the role of LAB as probionts and orally monitored via compounded feed on growth in P. indicus juveniles. It is thus possible to seed the gut with harmLess bacteria which occupy the attachment sites and prevent infection by pathogenic bacteria by maintaining artificially these bacterial populations at high level by regular intake with feed. Such treatments would be beneficial to improve health and quality of shrimp in culture. These LAB Probiotics could be complementary therapy for restoring balance to the intestinal flora. Further research on these lines using molecular and immunological approaches will provide conclusive evidence for these studies.
ACKNOWLEDGMENTS
The authors are grateful to the Director, C.M.F.R. Institute, Head, Physiology, Nutrition and Pathology Division and Officer in charge, Post-graduate Programme in Mariculture for providing the necessary facilities for carrying out the research work.
REFERENCES
- Askarian, F., A. Matinfar, A. Kousha, M. Bahmani, K. Khorshidi, A. Shenavar and E. Ringo, 2008. Diversity of lactic acid bacteria in the gastrointestinal tracts of reared beluga (Huso huso) and Persian sturgeon (Acipenser persicus): A comparative study. J. Fish. Aquatic Sci., 3: 302-311.
CrossRefDirect Link - Gatesoupe, F.J., 1991. The effect of three strains of lactic bacteria on the production rate of rotifers, Brachionus plicatilis and their dietary value for larval turbot, Scophthalmus maximus. Aquaculture, 96: 335-342.
CrossRefDirect Link - Gatesoupe, F.J., 1993. Bacillus sp. Spores as Food Additive for their Rotifer Brachionus Plicatilis: Improvement of their Bacterial Environment and their Dietary Value for Larval Turbot, Scophthalmus maximus L. In: Fish Nutrition in Practice, Kaushik, S.J. and P. Luquet (Eds.). Institute National de la Recherche Agronomique, Paris, France, pp: 561-568.
- Gatesoupe, F.J., T. Arakawa and T. Watanabe, 1989. The effect of bacterial additives on the production rate and dietary value of rotifers as food for Japanese flounder, Paralichthys olivaceus. Aquaculture, 83: 39-44.
CrossRef - Gatesoupe, F.J., 1999. The use of probiotics in aquaculture. Aquaculture, 180: 147-165.
CrossRefDirect Link - Gomez-Gil, B., A. Roque and J.F. Turnbull, 2000. The use and selection of probiotic bacteria for use in the culture of larval aquatic organisms. Aquaculture, 191: 259-270.
CrossRefDirect Link - Gursoy, O. and O. Kinik, 2006. Probiotics: A new popular option for cancer inhibition. Int. J. Dairy Sci., 1: 100-103.
CrossRefDirect Link - Irianto, A. and B. Austin, 2002. Use of probiotics to control furunculosis in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis., 25: 333-342.
CrossRefDirect Link - Joborn A., J.C. Olsson, A. Westerdahl, P.L. Conway and S. Kjelleberg, 1997. Colonization in the fish intestinal tract and production of inhibitory substances in intestinal mucus and faecal extracts by Carnobacterium sp. strain K1. J. Fish Dis., 20: 383-392.
CrossRef - Kabir, S.M.L., M.M. Rahman, M.B. Rahman, M.M. Rahman and S.U. Ahmed, 2004. The dynamics of probiotics on growth performance and immune response in broilers. Int. J. Poult. Sci., 3: 361-364.
CrossRefDirect Link - Moriarty, D.J.W., 1998. Control of luminous Vibrio species in penaeid aquaculture ponds. Aquaculture, 164: 351-358.
CrossRefDirect Link - Rengpipat, S., W. Phianphak, S. Piyatiratitivorakul and P. Menasveta, 1998. Effects of a probiotic bacterium on black tiger shrimp Penaeus monodon survival and growth. Aquaculture, 167: 301-313.
CrossRefDirect Link - Rengpipat, S., A. Tunyanun, A.W. Fast, S. Piyatiratitivorakul and P. Menasveta, 2003. Enhanced growth and resistance to Vibrio challenge in pond-reared black tiger shrimp Penaeus monodon fed a Bacillus probiotic. Dis. Aquat. Organ., 55: 169-173.
CrossRefPubMedDirect Link - Ringo, E. and F.J. Gatesoupe, 1998. Lactic acid bacteria in fish: A review. Aquaculture, 160: 177-203.
CrossRefDirect Link - Sissons, J.W., 1989. Potential of probiotic organisms to prevent diarrhoea and promote digestion in farm animals: A review. J. Sci. Food Agric., 49: 1-13.
CrossRefDirect Link - Soundarapandian, P. and S. Sankar, 2008. Effect of probiotics on the survival and production of black tiger shrimp Penaeus monodon (Fabricius). Int. J. Zool. Res., 4: 35-41.
CrossRefDirect Link - Sridhar, M., N. Sridhar, P.A.W. Robertson and B. Austin, 2006. Role of gut probionts in enhancing growth and disease resistance in rainbow trout (Oncorhynchus mykiss, Walbaum) fingerlings. Asian Fisher. Sci., 19: 1-13.
Direct Link - Vaseeharan, B. and P. Ramasamy, 2003. Control of pathogenic Vibrio spp. by Bacillus subtilis BT23, a possible probiotic treatment for black tiger shrimp Penaeus monodon. Lett. Applied Microbiol., 36: 83-87.
Direct Link - Verschuere, L., G. Rombaut, P. Sorgeloos and W. Verstraete, 2000. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev., 64: 655-671.
CrossRefPubMedDirect Link - Vijayabaskar, P. and S.T. Somasundaram, 2008. Isolation of bacteriocin producing lactic acid bacteria from fish gut and probiotic activity against common fresh water fish pathogen Aeromonas hydrophila. Biotechnology, 7: 124-128.
CrossRefDirect Link - Vine, N.G., W.D. Leukes and H. Kaiser, 2006. Probiotics in marine larviculture. FEMS Microbiol. Rev., 30: 404-427.
CrossRefPubMedDirect Link - Wang, Y.B., 2007. Effect of probiotics on growth performance and digestive enzyme activity of the shrimp Penaeus vannamei. Aquaculture, 269: 259-264.
CrossRefDirect Link - Wang, Y. and Z. Xu, 2006. Effect of probiotics for common carp (Cyprinus carpio) based\on growth performance and digestive enzyme activities. Anim. Feed Sci. Technol., 127: 283-292.
CrossRefDirect Link - Piard, J.C. and M. Desmazeaud, 1992. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances. Lait, 72: 113-142.
CrossRefDirect Link