
M. Karmakar
Microbiology Laboratory, Department of Zoology, Molecular Biology and Genetics, Presidency College, Kolkata, 700 073, India
R.R. Ray
Microbiology Laboratory, Department of Zoology, Molecular Biology and Genetics, Presidency College, Kolkata, 700 073, India
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 41-53







ABSTRACT
Bioconversion of cellulose, nature’s most abundant polysaccharide is accomplished by the enzyme cellulase. Cellulase, a complex of sequentially acting component enzymes are found to be synthesized by a variety of microbes. A number of different catalytic and non catalytic enzyme modules form the cellulase which synergistically acts on their substrates are reported, amongst which in some anaerobic bacteria different cellulases are organized together to form a multifunctional supramolecular complex known as cellulosome. Basic and applied research on microbial cellulases generated great scientific knowledge, their mode of action and their enormous industrial applications. Microbial cellulases are used in food, animal feed, brewery, wine, textile, laundry, paper, pulp and agriculture as well as in research purposes. The ever increasing demand for relatively non expensive cellulases and their enhanced utilization in multiple sectors of industries is the main driving force for research on different microbial cellulases.
PDF Abstract XML References Citation
Received: March 18, 2010;
Accepted: April 26, 2010;
Published: August 18, 2010
How to cite this article
M. Karmakar and R.R. Ray, 2011. Current Trends in Research and Application of Microbial Cellulases. Research Journal of Microbiology, 6: 41-53.
URL: https://scialert.net/abstract/?doi=jm.2011.41.53
URL: https://scialert.net/abstract/?doi=jm.2011.41.53
INTRODUCTION
Cellulose rich plant biomass is one of the foreseeable and sustainable source of fuel, animal feed and feed stock for chemical synthesis (Bhat, 2000). Therefore the conversion of cellulosic biomass to fermentable sugars and alcohols through biocatalyst cellulase produced by various cellulolytic organisms has attracted a world wide attention (Ladisch et al., 1983). Complete enzymatic hydrolysis of cellulose requires synergistic action of three cellulase enzymes: endoglucanase, exoglucanase and beta-glucosidases (Ryu and Mandels, 1980). Extensive basic and applied research on cellulases revealed their commercial significance and industrial applicability (Bajpai, 1999; Bayer et al., 1994; Beguin and Aubert, 1994; Bhat and Bhat, 1997; Gilbert and Hazlewood, 1993; Godfrey and West, 1996; Harman and Kubicek, 1998; Poutanen, 1997; Saddler, 1993; Uhlig, 1998). In spite of their commercial importance, the high cost of production of these enzymes has hindered the industrial application of cellulose bioconversion (Narasimha et al., 2006) therefore, there is an ever increasing demand for more stable, highly active, specific enzymes of nominal cost. In order to increase the production and applicability of microbial cellulase, various biotechnological approaches were adopted by the scientists. Biotechnology of cellulases began in early 1980s, first in animal feed followed by food applications (Voragen, 1992) later in the textile, laundry as well as in the pulp and paper industries (Godfrey and West, 1996). During the last two decades, the use of cellulases has increased considerably, especially in textile, in the bioprocessing of natural fibers, such as for the hydrolysis of cellulose to fermentable sugars and ethanol production (Tolan and Foody, 1999); de-inking of recycled paper (Smook, 1992; Moekerbak and Zimmermann, 1998); biopolishing of cotton fabrics to enhance softness and appearance and treatment of recycled fibers to restore fiber texture and flexibility lost during operations (Smook, 1992; Moekerbak and Zimmermann, 1998; Bhat et al., 1991; Pommier et al.,1989), in pulp and paper industries (Godfrey and West, 1996; Harman and Kubicek, 1998; Saddler, 1993; Uhlig, 1998). It has also been shown that cellulase treatment in combination with physical refining can provide a means for altering the morphology of coarse wood fibers (e.g., Douglas fir) to produce finer paper products (Mansfield et al., 1999). A number of review works are available on the production and application of microbial cellulases (Sukumaran et al., 2005), but the increasing application of cellulases in various sectors of food, textile and allied industries demand the understanding of the exact mechanism of action of these enzymes Therefore, the present review intends to highlight the recent researches on the mode of action of microbial cellulases of commercial significance and their main uses in various industries.
Mode of action: Before studying the applications of the cellulase enzyme, one needs to know the mode of action of the enzyme and the actual role of each component of the enzyme in performing the specific reaction. A cellulose enzyme system consists of three major components: endoglucanase (EC 3.2.1.4), exoglucanase (EC 3.2.1.91) and β-glucosidase (EC 3.2.1.21). of which endoglucanase- (EC 3.2.1.4; 1,4-β-D-glucan glucanohydrolase) acts on carboxy methyl cellulose, causing random scission of cellulose chains yielding glucose and cello-oligosaccharides; exoglucanase-(EC 3.2.1.91; 1,4-β-D-glucan cellobiohydrolase) acts on microcrystalline cellulose (avicel),imparting an exo-attack on the non-reducing end of cellulose, liberating cellobiose as the primary product and beta-glucosidases (EC 3.2.1.21) that facilitates the hydrolysis of cellobiose to glucose (Fig. 1). All these enzymes act synergistically to release glucose as end product. The commonly described mode of action for cellulases on polymers is either exo-or endo-cleavage and all cellulases target the specific cleavage of β-1,4-glycosidic bonds (Wood and McCrae, 1979).
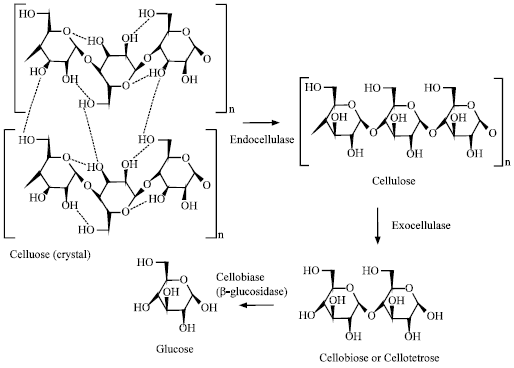 | |
| Fig. 1: | Mode of action of various components of cellulase |
Endo-β-glucanase acts randomly on the cellulose chain, to produce cello-oligosaccharides, while exo-β-glucanase acts on exposed chain ends by splitting off cellobiose. Cellobiose is subsequently hydrolysed by cellobiase to form glucose. This hypothesis is however more applicable and acceptable worldwide for the decomposition of cellulosic biomass.
Cellulases are composed of independently folding, structurally and functionally discrete units called domains or modules, making cellulases module (Henrissat et al., 1998). A free cellulase is composed of a Carbohydrate Binding Domain (CBD) at the C-terminal joined by a short poly-linker region to the catalytic domain at the N-terminal. There are only two modes of action for the hydrolysis of cellulose by cellulases, either inversion or retention of the configuration of the anomeric carbon. At least two amino acids with carboxyl groups located within the active site catalyze the reaction by acid-base catalysis. Using this classification system, cellobiohydrolases (exoglucanases) were classified as exo-acting based on the assumption that they all cleave β-1,4-glycosidic bonds from chain ends. As well, those enzymes truly exo-acting often have a tunnel-shaped closed active site which retains a single glucan chain and prevents it from re-adhering to the cellulose crystal (Rouvinen et al., 1990; Divne et al., 1994, 1998). While endoglucanases on the other hand, are often classified as endo-acting cellulases because they are thought to cleave β-1,4-glycosidic bonds internally only and appear to have cleft-shaped open active sites. Endoglucanase are active on amorphous regions of cellulose and thus their activity can be assayed using soluble cellulose substrates; i.e., the carboxymethylcellulase assay (CMCase). However, there is evidence that some cellulases display both action, endo-and exo-function (Davies and Henrissat, 1995). Thus classification has changed; cellobiohydrolases (exoglucanases) are described as active on the crystalline regions of cellulose; whereas, endoglucanases are typically active on the more soluble amorphous region of the cellulose crystal. There is a high degree of synergy seen between cellobiohydrolases (exoglucanases) and endoglucanases and it is this synergy that is required for the efficient hydrolysis of cellulose. The products of endoglucanases and cellobiohydrolases, that are cellodextrans and cellobiose, respectively, are inhibitory to the enzymes’ activity. Thus, efficient cellulose hydrolysis requires the presence of β-glucosidases which cleaves the final glycosidic bonds producing glucose (end product). Typically cellobiose and cellodextrins are taken up by the microorganism and internally cleaved via cellodextrin phosphorylases or cellobiose phosphorylases to create glucose monophosphate, which is energetically favoured. Some bacteria also produce inter-or extra-cellular β-glucosidases to cleave cellobiose and cellodextrins and produce glucose to be taken up by or assimilated by the cell. Mechanism of cellulose degradation by aerobic bacteria is similar to that of aerobic fungi but it is clear that anaerobic bacteria operate a different system (Coughlan et al., 1988). The cellulolytic systems of anaerobic bacteria have received in-depth study of late. Cellulase complexes located on the cell surface mediate adherence of anaerobic cellulolytic bacteria to the substrate. Evidence indicates that the anaerobic cellulolytic bacteria must attach themselves to cellulose to effect its degradation. Lamed and Bayer (1988) coined the term cellulosome to describe the multicomponent cellulolytic complex produced by C. thermocellum. Aggregates of these cellulosomes, i.e., polycellulosomes, located on the cell surface are responsible for attachment of the cell to the substrate, this adherence being specific for cellulose. The cellulosome, concept was originally proposed in case of the cellulase system of the thermophilic anaerobe Clostridium thermocellum. In this bacterium, the cellulosome is composed of a primary scaffoldin subunit that can bind up to nine enzymes into the complex, a process mediated by very strong intermodular interactions (Fig. 2). Free cellulase system which generally contain individual enzymes that bear a catalytic module together with a Cellulose-Binding Module (CBM).
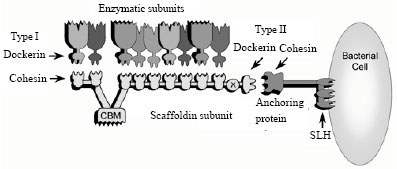 | |
| Fig. 2: | Schematic representation of the structure of cellulosome present in a bacterial cell membrane |
But in case of cellulosomal systems the scaffoldin subunit contains a single CBM together with numerous cohesion modules. The cohesion modules, in turn, bind strongly to a dockerin module borne by each cellulosomal enzyme. Cellulosomes derived from different bacteria show a divergent type of architecture, owing to the number of interacting scaffoldins and the content and specificities of their resident cohesins (Bayer et al., 2004). Cellulosomes of some bacteria like Acetivibrio cellulolyticus and Ruminococcus flavefaciens, can be much more intricate than those of C. thermocellum (Xu et al., 2004; Jindou, et al., 2006); the heterogeneity of cellulosome composition and assembly is still a mystery and an area of current research. A poly-cellulosome isolated from residual cellulose in cultures of C. thermocellum strain JW20 was found to have a diameter of 60 mm and a calculated mass of from 50 to 80x10 6 Da. It is comprised of a number of tightly packed spherical entities, viz., cellulosome, (16 to 18 nm in diameter) with a mass of 2 to 2.5x10 6 Da. The cellulosomes in turn are comprised of about 35 polypeptides ranging from 45 kDa to about 200 kDa. A polypoptide of 200-210 kb, which in the case of C. thermocellum responsible for attachment of the complex to the cell and to the substrate, is present in the complex produced by each organism. Presence of enzymes on the cell surface may make possible the topological arrangement required for hydrolysis of crystalline substrates. Anaerobic cellulolytic bacteria contain a β-glucosidase or cell bound cellobiose phosphorylase or both.
ROLE OF MICROBIAL CELLULASE IN VARIOUS INDUSTRIES
Cellulases have a wide range of enormous potential applications in biotechnology and many thermo stable endoglucanase appeared to have a great potentiality for industrial use (Karmakar and Ray, 2010a). In most of the cases they are used with hemicelluloses, pectinases, ligninase and allied enzymes. Some of the most important applications of cellulases are in food, brewery and wine, animal feed, textile and laundry, pulp and paper industries, as well as in agriculture and for research purposes. Details of most promising applications are discussed below.
Cellulase in food processing industries: Enzyme infusion has the potential of producing fruit and vegetable juices which is very important from commercial standpoint. The production of fruit and vegetable juices requires methods for extraction, clarification and stabilization. During early 1930s, when fruit industries began to produce juice, the yields were low and many difficulties were encountered in filtering the juice to an acceptable clarity (Uhlig, 1998). Subsequently, research on industrially suitable macerating enzymes from food-grade micro-organisms (Aspergillus niger and Trichoderma sp.), together with increased knowledge on fruit components, helped to overcome these difficulties (Grassin and Fauquembergue, 1996). During the production of juice from fruits such as apples and pears, the whole fruits were crushed to pulp mash, which, after mechanical processing (pressing, centrifuging and filtering), resulted into a clear fruit juice and a solid phase called pomace (Galante et al., 1998). Application of macerating enzymes could increase both production and process performance without additional capital investment. Macerating enzymes are generally used after crushing, to macerate the fruit pulp for partial or complete liquifaction, which increases the juice yield, reduces the processing time and improves the extraction of valuable fruit components. Thus, the macerating enzymes, composed of mainly cellulase and pectinase play a key role in food biotechnology and their demand will likely increase for extraction of juice from a wide range of fruits and vegetables including olive oil extraction, that has attracted the world market because of its numerous health claims. In addition, the infusion of pectinases and b-glucosidases increases the aroma and volatile characteristics of specific fruits and vegetables (Humpf and Schrier, 1991; Krammer et al., 1991; Marlatt et al., 1992; Pabst et al., 1991).
Cellulase in pharmaceutical industries: Since, humans poorly digest cellulose fiber, taking a digestive enzyme product, like Digestin, that contains cellulase enzymes is not only necessary, but also vital for healthy cells.
Cellulase in brewery and wine industries: Bioconversion of cellulosic materials to bioalcohol involves a multistep process which first uses cellulolytic enzymes for hydrolysis of polymers to pentose/hexose sugars and fermentation followed by distillation of these sugars into ethanol. In the beer wort production, Pajunen (1986) opined that the enzyme preparation from Trichoderma was the best as judged by its cost /performance ratio. In an earlier study, Oksanen et al. (1985) observed that endoglucanase II and cellobiohydrolase II of the Trichoderma cellulase system were responsible for a maximum reduction in the degree of polymerisation and wort viscosity. Furthermore, a marked improvement in filterability was reported with increasing doses of enzyme when tested in pilot scale (Oksanen et al., 1985). Significant and reproducible improvements in grape pressability, settling rate and total juice yield were achieved using a combination of macerating enzymes. Such improvements were noticeable only with a correct balance of pectinolytic, cellulolytic and hemicellulolytic enzymes. Using three varieties (Soave, Chardonnay and Sauvignon) of white grapes from Northern Italy, Galante et al. (1998) assessed the performance of Cytolase 219 (a commercial enzyme preparation, derived from Trichoderma and Aspergillus, containing pectinase, cellulase and hemicellulase) in wine making. They reported a 10-35% increase in the extraction of the first wine must, a 70-180% increase in the must filtration rate, significant improvement in wine stability, 50–120 min decrease in pressing time, 30-70% decrease in must viscosity and 20-40% energy saving during cooling of fermenters. In fact, the enzyme technology offers enormous benefits to wine industry.
Cellulase in textile industries: Enzymes have been used in the leather industry for many years and more recently have been introduced into modern textile industries. Depending on the aim, the extent of cellulose degradation and the properties of the resulting products can be controlled by adjusting the treatment parameters (like time, enzyme concentration and the composition of cellulase mixture). The textile industries take advantage of both complete and individual cellulase components to achieve partial cellulose hydrolysis and improve fabric properties, where the cellulase would act upon the fibre to reduce the cell wall thickness and would make the fibre more flexible and collapsible. While in case of bio-polishing, endoglucanase rich cellulase would act for depiling and aging of the fabric. For example, complete cellulase mixtures are used in depilling/cleaning of cotton fabrics, whereas, pure endoglucanase (EG) or EG-rich mixtures are used to produce aged and soft fabrics demanded by the fashion market (Tolan and Foody, 1999; Cavaco-Paulo, 1998). It is postulated that during depilling, enzymes attack and hydrolyze the microfibrils that hold the pills to the fiber surface, whereas in fabric ageing, the attack occurs on the fiber surface and results in fiber defibrillation (Cavaco-Paulo, 1998). The accompanying mechanical action removes the dye bound to the surface and imparts an aged appearance (Cavaco-Paulo, 1998). In both cases, the accessibility of cellulose surface to the enzymes plays a key role. Blue jeans and other denim garments have gained remarkable popularity in recent years. In denim fabrics, the indigo dye is mostly attached to the surface of the yarn and to the most exterior short cotton fibres. Repeated washings of denim fabric showed the wash down or aged effect, on which the entire denim industry has been built. Commercial cellulases have also been shown to enhance the whiteness, brightness and color characteristics of cotton fabrics (Csiszar et al., 1998). The cellulase preparations capable of modifying the structure of cellulose fibrils are added to laundry detergents to improve the colour brightness, hand feel and dirt removal from cotton and cotton blend garments. Most cotton or cotton blend garments, during repeated washings, tend to become fluffy and dull. This is mainly due to the presence of partially detached microfibrils on the surface of garments that can be removed by cellulases in order to restore a smooth surface and original colour to the garment. Also, the degradation of microfibrils by cellulase, softens the garment and removes dirt particles trapped in the microfibril network. The retention of water by fibers during refining reduces the softening temperature of hemicellulose and lignin present between adjacent fibers and weakens inter-fiber bonding, hence improving the separation of fibers from one another and reducing the energy consumption during refining operation (Pere et al., 1996). It has been shown that cellobiohydrolase I, a cellulase component from Trichoderma reesei, could selectively reduce the crystallinity of cellulose and subsequently produce more amorphous material with a higher affinity for water. Treatment with CBH I was able to reduce the refining energy demands by 40% (Pere et al., 1996). The finishing of denim jeans has also become a popular application for cellulases in the textile industry. Traditionally denim was stonewashed with pumice stones to fade the surface of the garment. A small application of cellulase can replace many of the stones resulting in less damage to the garments and machinery. This technique has become known as Biostoning and can result in much greater fading without high abrasive damage both to the actual fabric and any other accessories (buttons, rivets) on the fabric. Stonewashing enzymes are usually available as either acid cellulases (optimum activity around 4.5) or neutral cellulases (optimum activity at just below pH 7.0).
Cellulase in detergent industries: Use of cellulase along with protease and lipase is a more recent innovation (Singh et al., 2007). Removal of oil from interfibre space by selective contraction of fibres by the alkaline cellulase increases the cleansing capacity of a detergent. Nowadays, liquid laundry detergent containing anionic or nonionic surfactant, citric acid or a water-soluble salt, proteolytic enzyme, cellulase and a mixture of 1,2 propane diol and boric acid or its derivative. The compositions are prepared by adding the diol and boric acid before adding the citric acid/salt to the composition. This order of addition improves the stability of the cellulase (Boyer and Farwick, 1995) As most of the cellulose fibres in the modern textile industry enzymes are used increasingly in the finishing of fabrics and clothes are arranged as long, straight chains some small fibres can protrude from the yarn or fabric. The correct application of a cellulase enzyme can remove these rough protuberances giving a smoother, glossier brighter colored fabric. This technique has become known biopolishing and results in not only a softer fabric but also improved color brightness. This process of washing has been adapted and included in some laundry detergents.
Applications of cellulase in pulp and paper industries: Use of cellulases along with xylanase and ligninase in the pulp and paper industries has increased considerably during last decade (Mai et al., 2004). Biopulping with the help of cellulases and allied enzymes is a better alternative for mechanical pulping process as the former provides significant energy savings as these enzymes require lower energy input to achieve the required freeness and strength and check the problem of pollution. Refining, of primary or secondary fibers, can generate small particles (fines) that can reduce the drainage rate of pulps during papermaking operations. Cellulases seem to preferentially attack and hydrolyze the fines produced during the refining operation and therefore, improve the pulp's drainage property. Cellulase and hemicellulases helps in modification of coarse mechanical pulp and handsheet strength properties, partial hydrolysis of carbohydrate molecules and the release of ink from fibre surfaces which results into deinking of recycled paper. Cellulases have also been used to remove ink from papers and to enhance papermaking properties of recycled fibers. Enzymatic deinking can lower the need for deinking chemicals and reduce the adverse environmental impacts of the paper industry (Stork and Puls, 1996). While in general, enzymatic deinking results in little or no loss in fiber strength (Stork and Puls, 1996; Sarkar et al., 1995; Sarkar, 1997), the overall effectiveness of the treatment depends on variables, such as toner quality and type, the type and amount of sizing and the presence of other contaminants (Sarkar et al., 1995). Although, strength properties have not been compromised substantially, the excessive use of enzymes must be avoided (Stork and Puls, 1996), as it has been shown that significant hydrolysis of the fines (Stork and Puls, 1996; Jeffries et al., 1996; Ow et al., 1996; Jackson et al., 1993) could reduce the bondability of the fibers (Karnis, 1995; Kibblewhite, 1975). It has been postulated that improvements in dewatering and deinking of various pulps results in the peeling of the individual fibrils and bundles, which have a high affinity for the surrounding water and ink particles (Kibblewhite et al., 1995). It appears that cellulase treatments can release ink particles bound to the fines and to the fiber and enhance the removal of ink by floatation. While cellulases clearly enhance the deinking process, the mechanical agitation still plays a critical role in the efficiency of ink removal. These claims are consistent with similar findings concerning enzymatic stone washing of cotton fabrics, which indicated that enzymatic treatments in combination with mechanical agitation improve the efficacy of the process (Lee and Kim, 1983; Welt and Dinus, 1995; Zeyer et al., 1993).
Application of cellulase in animal feed: Cellulases have potential application in animal feed industry consumed by poultry, pigs, ruminants as well as pet and fish farming. In todays world there is a great deal of interest in using enzyme preparations containing high levels of cellulase and hemicellulase activities for improving the feed utilization, milk yield and body weight gain by ruminants. Nevertheless, the successful use of these enzymes in animal diet is to: eliminate Anti-Nutritional Factors (ANF) present in grains or vegetables; degrade certain cereal components in order to improve the nutritional value of feed; and/or to supplement animals’ own digestive enzymes (e.g., proteases, amylases and glucanases). Moreover, Cellulases and hemicellulases are responsible for partial hydrolysis of lignocellulosic materials, dehulling of cereal grains, hydrolysis of b-glucans and better emulsification and flexibility of feed materials which results in the improvement in the nutritional quality of animal feed (Chesson, 1987; Cowan, 1996; Galante et al., 1998). Cellulases, hemicellulases and pectinases can cause partial hydrolysis of plant cell wall during silage and fodder preservation. They are responsible for the expression of preferred genes in ruminant and monogastric animals for high feed conversion efficiency. These commercially important enzymes can produce and preserve high quality fodder for ruminants; improving the quality of grass silage (Ali et al., 1995; Hall et al., 1993; Selmer-Olsen et al., 1993).
Application of cellulases in research and development: Mixture of different cellulase along with hemi-cellulase and pectinase have immense potential and application in research and development area for controlling plant diseases and enhancing plant growth. A cocktail of different cellullases,hemicellulases and pectinases results in the solubilisation of fungal or plant cell wall to produce protoplast (Beguin and Aubert, 1994; Bhat and Bhat, 1997). Cellulases and related enzymes are used in the biocontrol of plant pathogens and different plant diseases by inhibiting the germination of spores of the plant pathogens (Benitez et al., 1998; Chet et al., 1998). Even the cellobiohydrolase promoters of Tricoderma is used for the expression of the different proteins, enzymes, antibodies in large amount. These are some of the main cellulase application in the research area (Dunn-Coleman et al., 1991; Harkki et al., 1989; Penttila, 1998).
Application in waste utilization: Cellulose is the major part of plant biomass. Therefore, the wastes generated from forests, agricultural fields and agro industries contain a large amount of unutilized or underutilized cellulose. Agricultural and industrial wastes are among the causes of environmental pollution (Milala et al., 2005.) These wastes generally accumulate in the environment causing pollution problem (Abu et al., 2000). Nowadays, these so called wastes are judiciously converted into valuable products such as enzymes (Ray et al., 1994) Sugar (Ghosh and Ray, 2010), biofuels, chemicals, cheap energy sources for fermentation, improved animal feeds and human nutrients (Howard et al., 2003), which is accomplished by cellulase. Therefore, the discarded biomass and agrowastes are successfully utilized for the production of enzymes, sugar and alcohols (Karmakar and Ray, 2010b; Youssef and Berekaa, 2009; Acharya et al., 2008; Milala et al., 2009; Omosajola et al., 2008).
CONCLUSION
It was estimated that in 2000, the world sale of industrial enzymes have already reached a market of 1.6 billion US dollars (Demain, 2000) of which cellulase and allied enzymes occupy a significant position. Microbes are an attractive topic of interest for the production of cellulases and hemicellulases due to their immense potential for cellulase production, enzyme complexity and extreme habitat variability. Microbial cellulases are preferred for their vast industrial applicability and relatively lower cost of production. In fact the craze for these cellulase enzymes is increasing day by day worldwide for their use in food, pharmaceuticals, bioalcohols and other industries. More and more research works are resulting into improved scientific knowledge along with the success of meeting the growing demands of the cellulase and related enzymes for generation of environment friendly textiles, detergents, bio-pulping and bio-alcohols. Moreover, it is opening new avenues for utilization of various agrowastes and organic pollutants as a source of renewable resource instead of dumping them to cause environmental pollution. In near future newer knowledge of excellent cellulolytic and hemicellulolytic systems and adoption of different biotechnological strategies will definitely bring a great prospect in industrial microbiology.
ACKNOWLEDGMENT
The authors wish to thank the Department of Science and Technology (DST), West Bengal, India for the financial assistance.
REFERENCES
- Abu, E.A., P.C. Onyenekwe, D.A. Ameh, A.S. Agbaji and S.A. Ado, 2000. Cellulase (E.C.3.21.3) production from sorghum bran by Aspergillus niger SL 1: An assessment of pretreatment methods. Proceedings of the International Conference on Biotechnology: Commercialization and Food Security, (ICBCFS`00), Abuja, Nigeria, pp: 153-159.
- Acharya, P.B., D.K. Acharya and H.A. Modi, 2008. Optimization for cellulase production by Aspergillus niger using saw dust as substrate. Afr. J. Biotechnol., 7: 4147-4152.
Direct Link - Ali, S., J. Hall, K.L. Soole, C.M.C.A. Fontes, G.P. Hazlewood, B.H. Hirst and H.J. Gilbert, 1995. Targeted Expression of Microbial Cellulases in Transgenic Animals. In: Carbohydrate Bioengineering Progress in Biotechnology, Petersen, S.B., B. Svensson and S. Pedersen (Eds.). Vol. 10. 1st Edn., Elsevier, Amsterdam, ISBN-10: 0444822232, pp: 279-93.
- Bajpai, P., 1999. Applications of enzymes in the pulp and paper industry. Biotechnol. Prog., 15: 147-157.
CrossRef - Bayer, E.A., E. Morag and R. Lamed, 1994. The Cellulosome: A treasure-trove for biotechnology. Trends Biotechnol., 12: 379-386.
CrossRef - Bayer, E.A., J.P. Belaich, Y. Shoham and R. Lamed, 2004. The cellulosomes: Multi-enzyme machines for degradation of plant cell wall polysaccharides. Ann. Rev. Microbiol., 58: 521-554.
CrossRefDirect Link - Bhat, M.K. and S. Bhat, 1997. Cellulose degrading enzymes and their potential industrial applications. Biotechnol. Adv., 15: 583-620.
CrossRefDirect Link - Bhat, M.K., 2000. Cellulases and related enzymes in biotechnology. Biotechnol. Adv., 18: 355-383.
Direct Link - Bhat, G., J. Heitmann and T. Joyce, 1991. Novel techniques for enhancing the strength of secondary fiber. Tappi, 74: 151-156.
Direct Link - Beguin, P. and J.P. Aubert, 1994. The biological degradation of cellulose. FEMS Microbiol. Rev., 13: 25-58.
CrossRef - Benitez, T., C. Limon, J. Delgado-Jarana and M. Rey, 1998. Glucanolytic and Other Enzymes and their Genes. In: Trichoderma and Gliocladium-Enzymes, Biological Control and Commercial Applications, Harman, G.F. and C.P. Kubicek (Eds.). Vol. 2. Taylor and Francis, London, ISBN: 978-0-7484-0805-4, pp: 101-127.
- Cavaco-Paulo, A., 1998. Processing Textile Fibers with Enzymes: An Overview. In: Enzyme Applications in Fiber Processing, Eriksson, K.E. and A. Cavaco-Paulo (Eds.). American Chemical Society Washington, DC., ISBN-13: 9780841235472, pp: 180-189.
CrossRef - Csiszar, E., G. Szakacs and I. Rusznak, 1998. Bioscouring of Cotton Fabrics with Cellulase Enzyme. In: Enzyme Applications in Fiber Processing, Eriksson, K.E. and A. Cavaco-Paulo (Eds.). American Chemical Society, Washington, DC., ISBN-13: 9780841235472, pp: 204-211.
CrossRef - Davies, G.J. and B. Henrissat, 1995. Structures and mechanisms of glycosyl hydrolases. Structure, 3: 853-859.
CrossRef - Demain, A.L., 2000. Microbial biotechnology. Trends Biotechnol., 18: 26-31.
CrossRefPubMedDirect Link - Divne, C., J. Stahlberg, T. Reinikainen, L. Ruohonen and G. Petterson et al., 1994. The three-dimensional crystal structure of the catalytic core of cellobiohydrolase I from Trichoderma reesei. Science, 265: 524-528.
CrossRef - Divne, C., J. Stahlberg, T. Teeri and T.A. Jones, 1998. High-resolution crystal structures reveal how a cellulose chain is bound in the 50 A long tunnel of cellobiohydrolase I from Trichoderma reesei. J. Mol. Biol., 275: 309-325.
CrossRef - Dunn-Coleman, N.S., P. Bloebaum, R.M. Berka, E. Bodie and N. Robinson et al., 1991. Commercial levels of chymosin by Aspergillus. Biotechnology, 9: 976-981.
CrossRefDirect Link - Galante, Y.M., A. De Conti and R. Monteverdi, 1998. Application of Trichoderma Enzymes in Food and Feed Industries. In: Trichoderma and Gliocladium-Enzymes, Biological Control and Commercial Applications, Harman, G.F. and C.P. Kubicek (Eds.). Vol. 2. CRC Press, London, UK., ISBN: 978-0-748-40805-4, pp: 327-42.
- Ghosh, B. and R.R. Ray, 2010. Saccharification of raw native starches by extracellular isoamylase of Rhizopus oryzae. Biotechnology, 9: 224-228.
CrossRefDirect Link - Gilbert, H.J. and G.P. Hazlewood, 1993. Bacterial cellulases and xylanases. J. Gen. Microbiol., 139: 187-194.
Direct Link - Hall, J., A. Simi, M.A. Surani, G.P. Hazlewood and A.J. Clark et al., 1993. Manipulation of the repertoire of digestive enzymes secreted into the gastrointestional tract of transgenic mice. Biol. Technol., 11: 376-379.
CrossRef - Harkki, A., J. Uusitalo, M. Bailey, M. Penttila and J.K.C. Knowles, 1989. A novel fungal expression system: Secretion of active calf chymosin from the filamentous fungus Trichoderma reesei. Nature Biotechnol., 7: 596-603.
CrossRef - Henrissat, B., T.T. Teeri, R.A.J. and A.Warren, 1998. Scheme for designating enzymes that hydrolyse the polysaccharides in the cell walls of plants. FEBS Lett., 425: 352-354.
CrossRef - Howard, R.L., E. Abotsi, R.E.L. Jansen and S. Howard, 2003. Lignocellulose biotechnology: Issue of bioconversion and enzyme production. Afr. J. Biotechnol., 2: 602-619.
Direct Link - Humpf, H.U. and P. Schrier, 1991. Bound aroma compounds from the fruit and the leaves of Blackberry (Rubus laciniata L.). J. Agric. Food Chem., 39: 1830-1832.
CrossRefDirect Link - Jackson, L.S., J.A. Heitmann and T.W. Joyce, 1993. Enzymatic modifications of secondary fiber. Tappi, 76: 147-154.
Direct Link - Jeffries ,T.W., M.S. Sykes, K. Rutledge-Cropsey, J.H. Klungness and S. Abubakr, 1996. Enhanced Removal of Toners form Office Waste Papers by Microbial Cellulases. In: Biotechnology in the Pulp and Paper Industry: Recent Advances in Applied and Fundamental Research, Srebotnik, E. and K. Messner (Eds.). Vol. 1. Vienna, Austria, pp: 141-144.
- Jindou, S., I. Borovok, M.T. Rincon, H.J. Flint and D.A. Antonopoulos et al., 2006. Conservation anddivergence in cellulosome architecture between two strains of Ruminococcus flavefaciens. J. Bacteriol., 188: 7971-7976.
CrossRef - Karmakar, M. and R.R. Ray, 2010. Characterization of extracellular thermostable endoglucanase from Rhizopus oryzae using response surface methodology. Res. Rev. Biosci., 4: 50-55.
Direct Link - Karmakar, M. and R.R. Ray, 2010. Extra cellular endoglucanase production by rhizopus oryzae in solid and liquid state fermentation of agro wastes. Asian J. Biotechnol., 2: 27-36.
CrossRef - Karnis, A., 1995. The role of latent and delatent mechanical pulp fines in sheet structure and pulp properties. Paperi ja Puu-Paper Timber, 77: 491-497.
Direct Link - Kibblewhite, R.P., A.D. Bawden and C.L. Brindley, 1995. TMP fiber and fines qualities of 13 radiata pine wood types. APPITA, 48: 367-377.
Direct Link - Krammer, G., P. Winterhalter, M. Schwab and P. Schrier, 1991. Glycosidically bound aroma compounds in the fruits of prunus species: Apricot (P. armeniaca, L.), Peach (P. persica, L.), Yellow plum (P. domestica, L. ssp. Syriaca). J. Agric. Food Chem., 39: 778-781.
CrossRefDirect Link - Ladisch, M.R., K.W. Lin, M. Voloch and G.T. Tsao, 1983. Process considerations in the enzymatic hydrolysis of biomass. Enz. Microb. Technol., 5: 82-100.
CrossRef - Lee, S.B. and I.H. Kim, 1983. Structural properties of cellulose and cellulase reaction mechanisms. Biotechnol. Bioeng., 25: 33-51.
CrossRef - Mai, C., U. Kues and H. Miltz, 2004. Biotechnology in the wood industry. Applied Microbiol. Biotechnol., 63: 477-494.
CrossRefDirect Link - Marlatt, C., C.T. Ho and M. Chien, 1992. Studies of aroma constituents bound as glycosides in tomato. J. Agric. Food Chem., 40: 249-252.
CrossRefDirect Link - Milala, M.A., B.B. Shehu, H. Zanna and V.O. Omosioda, 2009. Degradation of agro-waste by cellulase from Aspergillus candidus. Asian J. Biotechnol., 1: 51-56.
CrossRefDirect Link - Moekerbak, A.L. and W. Zimmermann, 1998. Applications of Enzymes in Paper Deinking Processes. In: Enzyme Applications in Fiber Processing, Eriksson, K. and A. Cavaco-Paulo (Eds.). Vol. 687. American Chemical Society, Washington, D.C., ISBN-13: 9780841235472e, pp: 133-141.
CrossRef - Narasimha, G., A. Sridevi, B. Viswanath, M.S. Chandra and R.B. Rajasekhar, 2006. Nutrient effects on production of cellulolytic enzymes by Aspergillus niger. Afr. J. Biotechnol., 5: 472-476.
Direct Link - Omosajola, P.F. and O.P. Jilani, 2008. Cellulase production by Trichoderma longi, Aspergillus niger, Saccharomyces cerevisae cultured on Waste Materials from orange. Pak. J. Biol. Sci., 11: 2382-2388.
Direct Link - Pabst, A., D. Barron, P. Etievant and P. Schrier, 1991. Enzymatic hydrolysis of bound aroma constituents from raspberry fruit pulp. J. Agric. Food Chem., 39: 173-175.
CrossRef - Pere, J., M. Siika-Aho, L. Viikari, S. Liukkonen, J. Gullichsen, 1996. Use of purified enzymes in mechanical pulping. TAPPI Pulping Con., 2: 693-696.
Direct Link - Poutanen, K., 1997. Enzymes: An important tool in the improvement of the quality of cereal foods. Trends Food Sci. Technol., 8: 300-306.
CrossRef - Rouvinen, J., T. Bergfors, T.T. Teeri, J.K.C. Knowles and T.A. Jones, 1990. Three-dimensional structure of cellobiohydrolase II from Trichoderma reesei. Science, 249: 380-386.
CrossRef - Ryu, D.D.Y. and M. Mandels, 1980. Cellulases: Biosynthesis and applications. Enzyme. Microb. Technol., 2: 91-102.
CrossRefDirect Link - Sarkar, J., D. Cosper and E. Hartig, 1995. Applying enzymes and polymers to enhance the freeness of recycled fiber. TAPPI J., 78: 89-95.
Direct Link - Selmer-Olsen, I., A.R. Henderson, S. Robertson and R. McGinn, 1993. Cell wall degrading enzymes for silage. 1. The fermentation of enzyme-treated ryegrass in laboratory silos. Grass Forage Sci., 48: 45-54.
CrossRef - Stork, G. and J. Puls, 1996. Changes in Properties of Different Recycled Pulps by Endoglucanase Treatment. In: Biotechnology in the Pulp and Paper Industry: Recent Advances in Applied and Fundamental Research, Srebotnik, E. and K. Mesner (Eds.). Vol. 1. Facultas-Universitatsverlag, Vienna, pp: 145-150.
- Sukumaran, R.K., R.R. Singhania and A. Pandey, 2005. Microbial cellulases: Production, applications and challenges. J. Scient. Ind. Res., 64: 832-844.
Direct Link - Tolan, J.S. and B. Foody, 1999. Cellulase from Submerged Fermentation. In: Advances in Biochemical Engineering/Biotechnology, Scheper, Th. (Ed.). Vol. 65. Springer, Berlin / Heidelberg, ISBN: 978-3-540-65577-0, pp: 41-67.
CrossRef - Welt, T. and R.J. Dinus, 1995. Enzymatic deinking: A review. Progress Paper Recycling, 4: 36-47.
Direct Link - Wood, T.M. and S.I. McCrae, 1979. Synergism between enzymes involved in the solubilization of native cellulose. Adv. Chem. Ser., 181: 181-209.
CrossRef - Xu, Q., Y. Barak, R. Kenig, Y. Shoham, E.A. Bayer and R. Lamed, 2004. A novel Acetivibrio cellulolyticus anchoring scaffoldin that bears divergent cohesins. J. Bacteriol., 186: 5782-5789.
CrossRef - Youssef, G.A. and M.M. Berekaa, 2009. Improved production of endoglucanase enzyme by Aspergillus terreus: Application of plackett burman design for optimization of process parameters. Biotechnology, 8: 212-219.
CrossRefDirect Link
Sukhveer Singh Reply
let me explore my knowledge. thank you