
P.D. Belur
Department of Chemical Engineering, National Institute of Technology Karnataka, Srinivasnagar, Surathkal, Karnataka, 575 025, India
G. Mugeraya
Department of Chemical Engineering, National Institute of Technology Karnataka, Srinivasnagar, Surathkal, Karnataka, 575 025, India
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 25-40







ABSTRACT
Tannin acyl hydrolase (E.C.3.1.1.20) is commonly referred as tannase, hydrolyses ester and depside bonds of hydrolysable tannins to produce gallic acid, glucose and galloyl esters. Tannase finds application in many industrial sectors which includes pharmaceutical, food, chemical and beverages industry. The enzyme has potential uses in the treatment of tannery effluents and pre-treatment of tannin containing animal feed. Since, the discovery of tannase in 1867, a great deal of research did happen on production aspects of tannase. Most of the research was focused on fungal tannase, as tannin was earlier considered as bacteriostatic. After the discovery of bacterial tannase in 1983, several studies on bacterial tannase were published. Despite the long history and numerous publications, tannase is still considered as one of the costly industrial enzymes. This is due to less titer and long fermentation time of the processes. In view of the growing demand, it is imperative to isolate high productive strains and develop economically feasible processes. This study reviews the microbial sources, isolation and screening methods, modes of production, substrates and media, temperature and pH of fermentation, duration of fermentation and location of tannase enzyme. An attempt is also made to give an outline of historical development which has taken place in tannase research.
PDF Abstract XML References Citation
Received: April 05, 2010;
Accepted: June 15, 2010;
Published: August 18, 2010
How to cite this article
P.D. Belur and G. Mugeraya, 2011. Microbial Production of Tannase: State of the Art. Research Journal of Microbiology, 6: 25-40.
URL: https://scialert.net/abstract/?doi=jm.2011.25.40
URL: https://scialert.net/abstract/?doi=jm.2011.25.40
INTRODUCTION
Tannin acyl hydrolase (E.C.3.1.1.20) is commonly referred as tannase, catalyses the hydrolysis of ester bond (galloyl ester of an alcohol moiety) and the depside bond (galloyl ester of gallic acid) of hydrolysable tannins (tannic acid) (Haslam and Stangroom, 1966). Tannase activity is found to be sequential, whereby cleavage of depside bonds occurs first, followed by the hydrolysis of the ester bonds. Hydrolysis of tannic acid by tannase results in the liberation of glucose, gallic acid and various galloyl esters of glucose (Van de Lagemaat and Pyle, 2006). Tannins are toxic and bacteriostatic compounds making non-reversible complex with proteins. Tannin protects the vulnerable parts of plants from microbial attack by inactivating the invasive microbial extracellular enzymes. Despite the antimicrobial properties of tannins, many fungi, bacteria and yeasts are quite resistant to tannins and can grow and develop on them (Bhat et al., 1998). The mechanisms by which bacteria can overcome inhibition include tannin modification, degradation, dissociation of tannin-substrate complexes, tannin inactivation by high affinity binders, membrane modification/repair and metal ion sequestration (Smith et al., 2005). Degradation of tannin is achieved by the enzyme tannase. Many microorganisms evolved the ability to produce tannase. Tannase hydrolyses tannins thereby reduce the concentration and also produce the products, some of which are used as energy source. Gallic acid is one of the major products of tannic acid degradation. The gallic acid is readily utilized as substrate by oxidative breakdown to simple aliphatic acids, which then enter citric acid cycle (Bhat et al., 1998).
Major applications of this enzyme are in the production of gallic acid. Gallic acid is used for the manufacture of an antimalarial drug Trimethoprim. Gallic acid is a substrate for the chemical and enzymatic synthesis of propyl gallate, used as antioxidants in fats and oils, as well as in beverages industry (Van de Lagemaat and Pyle, 2006).
Tannase is also used as clarifying agent in some wines, juices of fruits and refreshing drinks with coffee flavor. In case of wines, the tannins often get oxidized to quinones by contact with the air, which lead to undesirable turbidity poses a serious quality problem. Tannase usage offers a viable solution for this. In the manufacture of beer, the tannins are added in the form of hops. Higher quantities of proteins in the beer, results in an undesirable turbidity. This is due to complex formation between protein and the hops tannin. This problem could be resolved by the usage of tannase (Belmares et al., 2004).
Tannase is also being used for the production of instant tea preparations, which are cold water soluble (Lekha and Lonsane, 1997). Further, it has been proved that the tannase treated green tea shows higher antioxidant properties than the normal green and black tea. Tannase treated green tea effectively inhibit the N-nitrosamines, which are carcinogenic, mutagenic and teratogenic found in most of the preserved meat (Lu and Chen, 2007). Several studies also reported a better color stability and organoleptic properties in tannase treated green tea (Lu et al., 2009).
Tannase applications in food and beverage industrial products contribute to remove the undesirable effects of tannins. Enzymatic treatment of fruit juices to reduce the bitterness has got advantages such as the higher quality of juice due to the lower haze and non-deterioration of juice quality. New fruit juices (pomegranate, cranberry, raspberry, cold tea, etc.) have recently been acclaimed for their health benefits, in particular, for its disease-fighting antioxidant potential. The presence of high tannin content in those fruits is responsible for haze and sediment formation, as well as for color, bitterness and astringency of the juice upon storage. Due to the inability of conventional fruit juice debittering processes to remove the bitterness effectively, enzymatic debittering should be preferred. Preliminary results reported by Rout and Banerjee (2006) for pomegranate juice demonstrated that tannase treatment resulted in 25% degradation of tannin, while a combination of tannase and gelatin (1:1) resulted in 49% of tannin degradation.
Tannins form insoluble complexes with proteins. This interaction plays an important role in nonruminants (rabbits, poultry etc.) industry. The tannin in many feed materials complex with dietary and endogenous proteins as well as digestive enzymes, there by interfering with normal digestion, leading to a drain of high quality protein from the body. Tannins are also known to interfere with the absorption of iron. Tannins also damage the mucosal lining of the gastrointestinal tract. The use of tannase as an ingredient of animal feed would improve the digestibility of the feed (Lekha and Lonsane, 1997). The enzyme has potential uses in the treatment of tannery effluents and pre-treatment of tannin containing animal feed (Aguilar et al., 2007; Murugan and Al-Sohaibani, 2010).
However, few of these potential applications have been commercially exploited mainly due to comparatively high cost of the enzyme (Van de Lagemaat and Pyle, 2006). Tannase is being produced and marketed by Biocon (India), Kikkoman (Japan) and ASA Specilaenzyme GmbH (Germany) (Belmares et al., 2004). Several patents regarding tannase applications have been taken over the years, which are listed in Table 1.
| Table 1: | Published patents on tannase application (Aguilar et al., 2007) |
 | |
Last 100 years has seen over 75 publications pertaining to tannase production. This review tries to summarize these contributions which enable us to understand and appreciate the advances, achievements and developments taken place in tannase production research.
TANNASE RESEARCH: HISTORICAL DEVELOPMENTS
An excellent review on historical developments of production and applications of tannase was reported by Lekha and Lonsane (1997). Scheele observed the presence of gallic acid in the aqueous extract of gallnut in 1786 (Knudson, 1913). Robiquet suspected the activity of an organism which causes fermentation and subsequent release of gallic acid in gall nuts. Loraque considered the formation of gallic acid from tannic acid to be due either to an organism or oxidation. To support this theory he further found that several toxic substances inhibit the formation of gallic acid from tannic acid in gall nut (Knudson, 1913). Van Teighem was the first to demonstrate that the formation of gallic acid is due to the action of fungus organism and neither to enzymes pre-existing in the galls, nor to oxidation by the air in 1867 (Knudson, 1913). He stated further that the organisms were Penicillium glaucum and the new organism which he named as Aspergillus niger. He further studied the submerged and surface growth of this organism and the extent of tannic acid degradation. Fernbach grew Aspergillus niger in Raulin’s solution with the sugar replaced by tannic acid and then extracted from the organism the enzyme tannase in 1901 (Knudson, 1913). The first extensive studies on tannase properties, sources, applications, reaction mechanisms and specificity were conducted by workers such as Fernbach, Pottevin, Dykerhoff and Ambruster and Thom and Raper at the start of the twentieth century. These studies moreover showed that tannase was an inducible enzyme and could be synthesized in solid-state fermentation by filamentous fungi such as Aspergillus and Penicillium. The application of tannase for the manufacture of gallic acid from tannin-containing materials was soon recognized. In 1943, Toth and Barsony demonstrated that tannase consists of an esterase and depsidase activity (Lekha and Lonsane, 1997). In 1960’s, tannase was characterized and purified from plant and fungal source by Madhavakrishna-Bose and Dhar-Bose, respectively (Lekha and Lonsane, 1997). Japanese workers Iibuchi et al. (1967) and Yamada et al. (1968) carried out purification and characterization studies on Aspergillus tannase. Iibuchi et al. (1967) also developed a spectrophotometric assay for the determination of tannase activity, which was formerly based on titration. In the early seventies, several patents were filed for potential application of tannase in food and beverage industry (Van de Lagemaat and Pyle, 2006). Around 1980’s many studies on the sources, assay, applications, immobilization, purification and characterization of tannase were published. It was discovered that apart from filamentous fungi, tannase was also found to be produced by animals (Lekha and Lonsane, 1997) and bacteria (Deschamps et al., 1983). Later on methods were developed to determine/detect tannase activity using gas chromatography (Jean et al., 1981).
There was a strong focus on the production of tannase by bacterial strains from 1990 onwards (Mondal et al., 2000, 2001; Mondal and Pati, 2000; Osawa et al., 2000; Ayed and Hamdi, 2002; Das Mohapatra et al., 2006; Selwal et al., 2010; Belur et al., 2010). Several studies demonstrated the potential advantages of solid state fermentation over submerged state fermentation using fungal cultures (Lekha and Lonsane, 1994; Kar and Banerjee, 2000; Aguilar et al., 2001b, 2002). Van de Lagemaat and Pyle (2001) developed a continuous solid state fermentation process for the production of fungal tannase. Hatamoto et al. (1996) were the first to clone sequence and amplify the tannase gene leading to elevated levels of tannase activity in Aspergillus oryzae transforms. Osawa and Walsh (1993) developed visual reading method for the detection of bacterial tannase. Bradoo et al. (1996) developed a rapid and simple plate assay method to screen extracellular tannase producing fungi. Sharma et al. (2000) developed a spectrophotometric method for the assay of tannase using rhodanine.
Several workers reported the purification and characterization of fungal tannase (Rajkumar and Nandy, 1983; Beverini and Metche, 1990; Lekha and Lonsane, 1994; Kar et al., 2003; Sabu et al., 2005; Kasieczka-Burnecka et al., 2007; Sharma et al., 2008; Belur et al., 2005, 2011). Several reports of productivity optimization studies are also available (Bradoo et al., 1997; Seth and Chand, 2000; Saxena and Saxena 2004; Purohit et al., 2006; Guzman-Lopez et al., 2009; Beniwal and Chhokar, 2010). Many workers carried out immobilization studies using fungal tannase (Abdel-Naby et al., 1999; Boadi and Neufeld, 2001; Sharma et al., 2002; Yu et al., 2004).
SOURCES OF TANNASE
Research over past 140 years resulted in the discovery of great variety of tannase producers. Bacteria, fungi and yeasts are the prominent producers. Few animals also have been found to be the producers of tannase. Further research showed that the colonizing microorganisms of these animals are the actual producers, not the animals.
Fungi: Filamentous fungi of the Aspergillus and Penicillium genus have been widely used for tannase production. Majority of the research work used these organisms. A brief list of these molds is given in Table 2.
Yeasts: There are very few reports of tannase production from yeasts, which are listed in Table 3.
Bacteria: Report on tannase from bacterial origin is scanty in literature before 1980’s. In last 25 years or so, about a dozen reports were published on bacterial tannase and about 25 new tannase positive bacteria have been isolated. Deschamps et al. (1983) isolated number of bacterial strains that can utilize tannic acid as the sole carbon source.
| Table 2: | Fungal sources of Tannase |
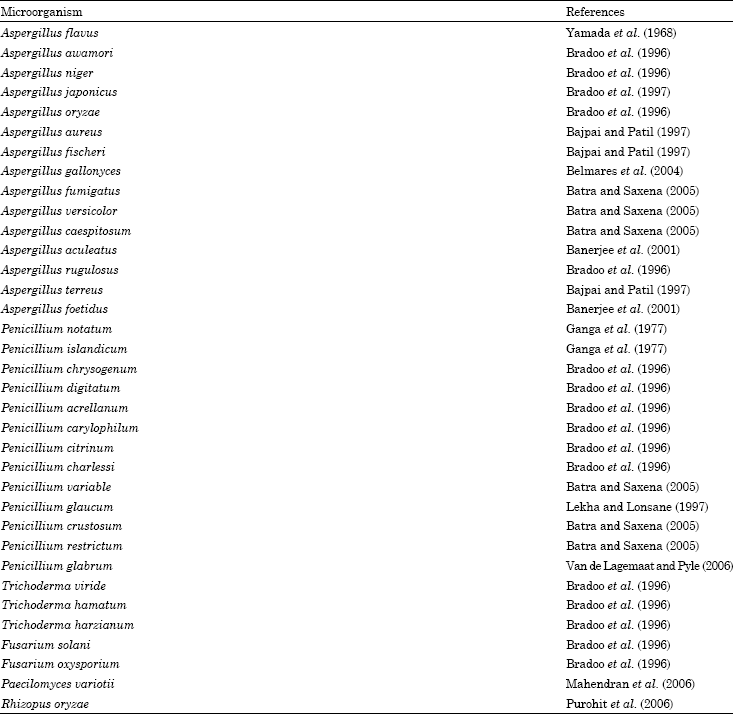 | |
| Table 3: | Yeast sources of tannase |
 | |
They were able to produce tannase from four strains using chestnut tannin and observed that gallic acid was the only degradation product. From the nineties onwards, there has been a strong focus on the production of tannase by bacterial strains. Several reports are now available which has been listed in Table 4.
| Table 4: | Bacterial sources of tannase |
 | |
Many gastrointestinal bacteria of adopted domesticated and wild animals have been found to produce tannase. Several species of these bacteria have been isolated from faeces of Koalas, goats, cows and human beings (Sabu et al., 2006; Nishitani et al., 2004; Goel et al., 2005).
Plants: Tannase has been reported to be present in many tannin-rich plant materials, such as Terminalia chebula (myrobolan) fruits, Caesalpinia coriaria (divi-divi) pods, Angeissus latifolia (dhawa) leaves and the bark of Cassia fistula (konnam) and Acacia Arabica (babul). Plant tannase is found to be less stable compared to microbial sources and purification is more cumbersome (Lekha and Lonsane, 1997).
ISOLATION AND SCREENING OF TANNASE POSITIVE MICROORGANISMS
Most of the literatures on fungal tannase production have reported the screening of fungal isolates procured from authentic culture collection centers. A selective solidified medium was designed to screen these cultures for their tannase activity. Few reports also mentioned the use of second screening in selective liquid cultures.
Yamada et al. (1968) had taken their own preserved cultures of fungus. They carried out first screening in a medium consists of tannic acid (1% w/v) as the sole carbon source and Sodium nitrate (0.2% w/v) as the sole nitrogen source and several salts. Lekha and Lonsane (1994) isolated Aspergillus niger PKL 104 from the soil by a baiting technique. Bradoo et al. (1996) had taken about 50 isolates from their stock culture collection and screened for their tannase producing ability. Screening was done in Czapek Dox’s minimal medium containing commercial tannic acid (1% w/v) as the sole carbon source and Sodium nitrate as the sole nitrogen source. They also developed a simple plate assay method to screen the potent producers and validated the technique. Pinto et al. (2001) took 30 strains of Aspergillus niger from EMBRAPA/Food technology stock collection and screened for their potential in tannase synthesis. Screening was performed in solidified selective medium consisting of tannic acid (1% w/v) as the sole carbon source and sodium nitrate (0.3% w/v) as the sole nitrogen source and salts. Batra and Saxena (2005) reported the screening of 60 Aspergilli and Penicilli procured from a laboratory stock culture collection and a few isolated from tannery effluents. They carried out screening using the method developed by Bradoo et al. (1996). Aissam et al. (2005) reported the isolation of Aspergillus niger HA37 from olive mill waste water. A synthetic selective liquid medium was used; inoculum from olive mill waste water was added and incubated for 72 h. The culture from this flask was subjected to four rounds of sub culturing in the same synthetic liquid medium. Murugan et al. (2007) isolated a strain of Aspergillus niger from composite tannery effluents of a tannery where skin and hides were processed by vegetable tanning method. The fungal forms were isolated using serial dilution and spread plate method on PDA medium.
All the available literature on bacterial tannase suggests that most of the authors isolated the organisms on their own, screened and identified them. The method of choice of isolation was liquid enrichment technique followed by isolation on selective solid media. Deschamps et al. (1983) isolated the five organisms from forest litter and decaying bark samples of oak and pine by enrichment culture using a mineral medium having condensed tannin as the sole carbon source. Kumar et al. (1999) isolated Citrobacter freundii from tannery effluents using a mineral medium having tannic acid by enrichment technique. Osawa et al. (2000) isolated three species of Lactobacillus from human feces and fermented foods. Mondal et al. (2000) isolated Bacillus licheniformis KBR 6 from forest soil by enrichment technique. Mondal et al. (2001) isolated number of tannase producing bacterial strains by plate dilution technique. They collected microbial samples form different forest soils of Midnapore district, W.B, India. They used a selective mineral medium consists of tannic acid (1% w/v) as the sole carbon source and Ammonium nitrate (0.3% w/v) as the sole nitrogen source with 3% agar. Mondal et al. (2001) isolated Bacillus cereus KBR9 from lateritic forest soil on selective tannic acid agar medium. Ayed and Hamdi (2002) isolated Lactobacillus plantarum from stored olive wastes. Nishitani et al. (2004) isolated 8 species of Lactobacillus from human feces and fermented foods based on the technique of Osawa et al. (2000). Sabu et al. (2006) isolated Lactobacillus sp. ASR-S1 from sheep excreta. Guzman-Lopez et al. (2009) purchased pure cultures from UAM-IRD collection (Institut de Recherche pour le Developpement, France). They were screened by culturing them on the selective medium having coffee pulp silage. Belur et al. (2010) have reported the isolation of five strains of bacteria from microbial samples collected from the effluent pit of a tea processing factory and rotting skin of grapes. The selection medium had tannic acid as the major carbon source along with yeast extract and other minerals.
PRODUCTION ASPECTS OF TANNASE
Filamentous fungi of the Aspergillus and Penicillium genus have been widely used for tannase production. Majority of the research work used these molds. Report on tannase from bacteria and yeast is scanty in literature.
Modes of production: Tannase production has been carried out in liquid-surface, submerged (SmF) and Solid-State Fermentation (SSF) in case of filamentous fungi. Bacteria and yeast studies were confined to SmF process, as these organisms prefer SmF process. In terms of productivity, location and stability of the enzyme, SmF may not hold advantage over other methods. Comparative studies by Lekha and Lonsane (1994) and Aguilar et al. (2001b) indicated that the productivity of an Aspergillus sp., was significantly higher in SSF than SmF. Lekha and Lonsane (1994) also found out that the enzyme was completely secreted out from the cell in SSF while it was partly intracellular in SmF. The enzyme from SSF showed better thermal and pH stability.
Almost all the reported studies used shake flasks to carry out SmF process. Very few laboratory studies have been carried out in laboratory fermenters (Pourrat et al., 1982; Deschamps et al., 1983; Seth and Chand, 2000; Banerjee et al., 2007). The SSF studies have reported the use of Petri dishes (Kar and Banerjee, 2000), Erlenmeyer flasks (Lekha and Lonsane, 1994), Column reactors (Aguilar et al., 2002) and a continuous solid state fermenter (Van de Lagemaat and Pyle, 2001). Surface liquid culture was carried out in Erlenmeyer flasks (Pourrat et al., 1982; Lekha and Lonsane, 1994).
Bacterial tannase production studies were carried out mostly by SmF in shaker flasks (Belur et al., 2010; Selwal et al., 2010; Das Mohapatra et al., 2006; Ayed and Hamdi, 2002; Mondal et al., 2000, 2001). However, Deschamps et al. (1983) carried out SmF trials in 2 L jar fermenter making use of their bacterial isolates. Sabu et al. (2006) carried out SSF trials in 250 mL Erlenmeyer flasks using Lactobacillus sp. ASR-S1.
Substrates and media: Both defined (synthetic) and undefined media have been employed for tannase production studies. Lekha and Lonsane (1994) used a synthetic medium consisting of only inorganic salts with 2% (w/v) of tannic acid in their shake flask experiment. Bradoo et al. (1997) reported the usage of Czapek-Dox’s minimal medium supplemented with tannic acid in their shake flask trials with Aspergillus japonicus organism. Various concentrations of tannic acid (0.5-10% w/v) were used as the sole carbon source. 2% (w/v) tannic acid gave highest enzyme productivity. They studied the effect of various sugar additives (0.2% w/v) viz., arabinose, fructose, galactose, xylose, lactose, sucrose, carboxy methyl cellulose, starch and mannitol on the enzyme production. They have reported that all the sugar additives except Glucose decreased the enzyme yield. Glucose concentration of 1% found to be optimal. Seth and Chand (2000), used a defined medium consisting of inorganic salts and tannic acid as the sole carbon source for their fermenter study on Aspergillus awamori, in an effort optimize the process parameters. The tannic acid concentration was varied between 2.5-4.5% (w/v), found 3.5% (w/v) as the optimum concentration. Saxena and Saxena (2004) reported the use of chebulic myrobalan powder as the source of tannic acid (32%) in their experiment on Penicillium variable. They optimized the production of tannase using this natural tannin by response surface methodology in shake flask experiments. 4-12% (w/v) of substrate powder has been used and found that 11.6% (w/v) was the optimum concentration of substrate powder. Lekha and Lonsane (1994), reported the use of sugar cane bagasse as the inert medium for solid state fermentation of spergillus niger PKL 104. Sugar cane pith bagasse was supplemented with a synthetic medium containing 6% (w/v) tannic acid. Aguilar et al. (2001b), reported the usage of Polyurethane foam as a support for the solid state fermentation of Aspergillus niger. A synthetic medium was used, having tannic acid concentration up to 20% (w/v). They reported that 10% (w/v) as the optimum concentration of tannic acid in the medium. Further, they used Glucose concentration of 0.65-20% (w/v) to supplement the medium having the tannic acid content of 2.5% (w/v). They reported a strong catabolite repression of tannase synthesis due to glucose supplementation. Ramirez-Coronel et al. (2003) have reported the usage of polyurethane foam as the support for carrying out solid state fermentation for Aspergillus niger. They had used a synthetic medium consists of mineral salts and 0.1% (w/v) of tannic acid as the sole carbon source. Sabu et al. (2005), have reported the use of Palm kernel cake and tamarind seed powder as the substrate for the production of tannase from Aspergillus niger ATCC 16620 by solid state fermentation. These media were supplemented with NH4NO3, MgSO4 and NaCl solution. Supplementation of Palm kernel cake media have been tried with 1.0% (w/v) of Glucose, Starch, Sucrose, Maltose, Glycerol, Tannic acid, Methyl gallate and Gallic acid. Battestin and Macedo (2007) have reported the use of Coffee husk and rice bran as the media ingredient for solid state fermentation of Paecilomyces variotii. These substances were supplemented with inorganic salt solution. Temperature, % of rice bran/coffee husk, tannic acid and salt solution have been chosen as variable to optimize the process by response surface methodology. Tannic acid at a concentration of 3-15%( w/w) was chosen. Optimum results were obtained at the tannic acid concentration of 15% (w/w) and residue of 50:50 of coffee husk: rice bran. Banerjee et al. (2007) have reported the use of raw tannin of Cassia siamea in 5 L fermenter. They reported good enzyme yield with 2.0% (w/v) of tannin in the synthetic medium.
Nitrogen sources often employed in tannase studies in defined media include sodium nitrate, ammonium chloride, ammonium di-hydrogen phosphate, ammonium oxalate, ammonium sulphate, monosodium glutamate and glutamic acid (Yamada et al., 1968; Lekha and Lonsane, 1997). Banerjee at al. (2007) have reported tannase production using ammonium phosphate as the sole nitrogen source. Most reported salts and trace elements in defined media were Na2HPO4, KCl, K2HPO4, KHPO4, MgSO4, FeSO4, AlCl3 (Van de Lagemaat and Pyle, 2006).
Deschamps et al. (1983) used a mineral medium with 1% (w/v) commercial chestnut extract as the main carbon source to produce tannase from Bacillus polymyxa, Corynebacterium sp. and Klebisella pneumoniae in a laboratory Fermentor. Kumar et al. (1999) used a mineral medium consists of tannic acid as high as 5% (w/v) to produce tannase from Citrobacter freundii. Mondal and Pati (2000) have reported the use of a synthetic medium having ammonium nitrate as the nitrogen source and tannic acid (1% w/v) as the sole carbon source for the shake flask studies of Bacillus lichiniformis KBR 6. Mondal et al. (2001) used a synthetic medium having tannic acid (1% (w/v) and Ammonium chloride as the nitrogen source for shake flask studies on Bacillus cereus KBR9. Ayed and Hamdi (2002) have reported the use of a semi synthetic medium for their shake flask study on Lactobacillus plantarum. They used tannic acid as the sole carbon source along with Ammonium sulfate and Casamino acids as the nitrogen sources. They used tannic acid in the concentration of 0.1-2.0% (w/v) and found highest activity in the medium having 1.5% (w/v) tannic acid. They also tried glucose supplementation and found that it has a positive effect on enzyme productivity. Das Mohapatra et al. (2006) used nine different plant extracts as the tannin source for tannase production from Bacillus lichiniformis KBR6. The extract was used with out any additives for the shake flask study. They obtained highest activity from the crude extract of Anacardium occidentale. Sabu et al. (2006) cultured Lactobacillus sp., ASR-S1 on tamarind seed powder, wheat bran, palm kernel cake and coffee husk. They used ammonium nitrate as the nitrogen source and mineral salts in water for adjusting the moisture content of the solid medium. They reported good productivity with coffee husk medium. Belur et al. (2010) have reported that semi synthetic medium having organic nitrogen source gave better productivity compared to inorganic nitrogen sources in case of their isolate Serratia ficaria DTC have reported that semi synthetic medium having organic carbon source gave better productivity compared inorganic nitrogen sources in case of their isolate Serratia ficaria. Selwal et al. (2010) reported that their strain, Pseudomonas aeruginosa III B 8914 was assessed for tannase productivity on the leaves of Phylanthus emblica (amla), Acacia nilotica (keekar), Eugenia cuspidate (Jamoa) and Syzygium cumini (Jamun) in shake flask trials. Powdered leaves (2% w/v) alone was used as the carbon source, Ammonium nitrate was used as the nitrogen source along with inorganic salts in the medium. They reported highest productivity in Phylanthus emblica (amla) containing medium. They also studied the effect of different carbon sources (0.2% w/v) viz., Dextrose, Mannitol, Glucose, Xylose, starch, lactose, glycerol, maltose, galactose and sucrose. They reported lesser enzyme yield compared to non-supplemented medium. Further, they tried various inorganic nitrogen sources, viz., sodium nitrate, ammonium chloride, urea, ammonium nitrate and ammonium sulphate, as an additive to the medium having amla leaves. They found better enzyme yield only in medium supplemented with ammonium nitrate. Additional carbon sources such as glucose, fructose and sucrose were employed in concentrations of 0.06-7.0% (w/v). This was presumably used to promote initial growth of biomass, as tannic acid is harder to metabolize than simple sugars. Most of these studies suggest that optimal concentration of tannic acid for growth and enzyme synthesis varies depending on the microorganism and fermentation condition used.
Fermentation temperature: Incubation temperature in the range of 25-40°C has been employed with most processes being conducted at around 30°C for fungal systems (Yamada et al., 1968; Rajkumar and Nandy, 1983; Lekha and Lonsane, 1994). Similar trend was observed in bacterial studies too. Kumar et al. (1999) reported maximum tannase activity at 30°C by Citrobacter freundii while Das Mohapatra et al. (2006) reported maximum tannase production at 35°C by Bacillus lichiniformis KBR6. Mondal et al. (2001) reported an optimum temperature of 40°C for their strain Bacillus cereus KBR9, while tannase production was optimum at 30°C in case of Lactobacillus sp. ASR-S1 (Sabu et al., 2006). Selwal et al. (2010) have reported an optimum temperature of 37°C for their culture Pseudomonas aeruginosa III B 8914.
Initial medium pH: The optimum initial medium pH appears to be between 4.5-7.0 for SmF and 5.0-6.5 for SSF for most of the fungal cultures (Van de Lagemaat and Pyle, 2006). In case of bacterial cultures, Selwal et al. (2010) have reported an optimum pH of 5.5 for their culture Pseudomonas aeruginosa III B 8914, while Mondal et al. (2001) reported an optimum pH of 5.0 for their strain Bacillus cereus KBR9. For Bacillus lichiniformis KBR6, Mondal and Pati (2000) have reported the optimum pH as 5.0. Ayed and Hamdi (2002) reported maximum tannase production at pH 6.0 for Lactobacillus Plantarum. Selwal et al. (2010) have found that initial medium pH of 7 was optimum for their strain Psuedomonas aeroginosa. Belur et al. (2010) have found that initial medium pH of 6.0 was the optimum for their strain Serratia ficaria DTC.
Duration of fermentation: In case of Fungi, maximum tannase productivity was observed during 12-24 h in SmF (Bradoo et al., 1997; Aissam et al., 2005; Aguilar et al., 2007) and 72-86 h in SSF (Lekha and Lonsane, 1994; Kar and Banerjee, 2000; Saxena and Saxena, 2004; Sabu et al., 2005). Banerjee et al. (2007) have reported peak tannase production at 36 h in 5 L fermentor using Aspergillus aculeatus DBF9.
In case of bacteria, the maximum productivity was reported with in 24 h of incubation. Maximum tannase activity was observed at the stationary phase of growth (24 h) by Selwal et al. (2010) for their culture Pseudomonas aeruginosa III B 8914, Ayed and Hamdi (2002) for their culture Lactobacillus plantarum and Mondal et al. (2001) for their culture Bacillus cereus KBR9. Same workers reported maximum tannase production at the late exponential phase of growth (21 h) in Bacillus lichiniformis KBR6 (Mondal et al., 2000; Mondal and Pati, 2000). Kumar et al. (1999) reported maximum tannase productivity in the early exponential phase (6 h) by Citrobacter freundii. Deschamps et al. (1983) have recorded optimum production after 6 h of incubation in their fermenter for Bacillus plumilus, Bacillus polymyxa, Corynebacterium sp. and Klebisella pneumoniae. Belur et al. (2010) have reported maximum enzyme production in the late stationary phase of growth , which was at 24th hour of incubation in case of Serratia ficaria DTC. Selwal et al. (2010) also have reported the similar trend in case of their isolate Pseudomonas aeroginosa IIIB 8914. Thus literature review suggests that optimal conditions can vary considerably depending on the microorganisms and type of fermentation.
Induction and inhibition mechanism: Tannase is reported to be an inducible enzyme produced in the presence of certain tannins or their degradation products. In case of fungal cultures, low constitutive levels of the enzyme were measured in the absence of tannic acid and tannase production increased significantly when the tannic acid was added to the culture medium (Bradoo et al., 1997; Aguilar et al., 2001a). Enzyme titer also found to increase with the increase in tannic acid concentration (Lekha and Lonsane, 1997). It is therefore thought that tannase production was induced by tannic acid or some of its derivatives, but the exact mechanism is unclear. Lekha and Lonsane (1997) have questioned the inductive role of tannic acid due to its large molecular size and reactivity and suggested that gallic acid acted as an inducer. However, other workers demonstrated that gallic acid did not induce tannase formation and in some cases even showed end-product repression (Bradoo et al., 1997; Aguilar et al., 2001a). In a study of Bajpai and Patil (1997), both tannic acid and gallic acid could function as inducer. Apart from tannic acid and gallic acid, methyl gallate has also been reported to possess inductive properties (Bajpai and Patil, 1997; Sabu et al., 2005). Inhibition of tannase formation by catabolite repression or feedback inhibition was not observed. Presence of additional carbon sources such as glucose, fructose and sucrose gave better productivity (Aguilar et al., 2001a; Van de Lagemaat and Pyle, 2001). This was presumably used to promote initial growth of biomass, as tannic acid is harder to metabolize than simple sugars. However, Bradoo et al. (1997) and Aguilar et al. (2001b) showed end-product repression on tannase with gallic acid and glucose, respectively.
In case of bacteria, Mondal et al. (2000) and Mondal and Pati (2000) observed that addition of low concentration of glucose, lactose and sucrose (0.1% w/v) were not repressive, but at high concentrations (0.3 and 0.5%), repressive effect was witnessed. Sabu et al. (2006) reported the inhibition of tannase production when the medium was supplemented with 1% (w/v) carbon sources in solid state fermentation of Lactobacillus sp. ASR-S1. Selwal et al. (2010) reported that carbon source supplementation suppressed the enzyme yield. These contradictions indicate that the regulatory mechanism may vary for different microorganisms and fermentation conditions.
TANNASE LOCATION
In case of fungi, tannase is found to be intracellular in submerged fermentation and extracellular in solid state fermentation. Several authors have reported that enzyme was intracellular when SmF was employed. For, example, Penicillium Chrysogenum, (Rajkumar and Nandy 1983), Aspergillus niger (Pourrat et al., 1982), Aspergillus flavus (Yamada et al., 1968). Tannase produced by Aspergillus niger PKL 104 in SmF was completely intracellular during initial 48 h of growth. Subsequently enzyme was secreted into the medium with progress of the fermentation. However, tannase produced by the same culture in SSF was completely extracellular through out the course of fermentation (Lekha and Lonsane, 1994). Seth and Chand (2000) have found that tannase was completely intracellular even after 60 h of incubation in laboratory fermenter in case of Aspergillus awamori.
In case of bacteria, except one report by Belur et al. (2010) all the reports are pertaining to extracellular tannase. They have reported for the first time that all the five strains of bacteria isolated by them showed cell-associated tannase activity, where as only three among them showed extracellular activity.
CONCLUSION
Since, the discovery of tannase in 1867, many research groups worked on microbial tannase production. In the last hundred years a great deal of research did happened on production aspects. Most of the research activities were focused on fungal tannase till the discovery of bacterial tannase by Deschamps et al. (1983). After this breakthrough research in bacterial tannase, about dozen papers were published on tannase production using various bacterial isolates. The discovery of cell-associated tannase in several bacterial isolates by Belur et al. (2010) could be of significant interest due to the fact that cell-associated enzyme generally exhibit higher stability and unique properties. The review suggests that the optimum fermentation conditions and regulatory mechanism of enzyme production seems to vary from one organism to other and generalization may not be possible. In view of growing demand for the tannase and its potential applications in food, pharmaceutical and chemical industries, it is imperative to isolate high productive strains and develop economically feasible processes.
REFERENCES
- Abdel-Naby, M.A., A.A. Sherif, A.B. El-Tanash and A.T. Mankarios, 1999. Immobilization of Aspergillus oryzae tannase and properties of the immobilized enzyme. J. Applied Microbiol., 87: 108-114.
CrossRefDirect Link - Aguilar, C.N., C. Augur, E. Favela-Torres and G. Viniegra-Gonzalez, 2001. Induction and repression pattern of fungal tannase in solid state and submerged cultures. Process Biochem., 36: 565-570.
CrossRefDirect Link - Aguilar, C.N., C. Augur, E. Favela-Torres and G. Viniegra-Gonzalez, 2001. Production of tannase by Aspergillus niger Aa-20 in submerged and solid-state fermentation: Influence of glucose and tannic acid. J. Ind. Microbiol. Biotechnol., 26: 296-302.
Direct Link - Aguilar, C.N., R. Rodriguez, G. Gutierrez-Sanchez, C. Augur and E. Favela-Torres et al., 2007. Microbial tannases: Advances and perspectives. Applied Microbiol. Biotechnol., 76: 47-59.
CrossRefDirect Link - Aissam, H., F. Errachidi, M.J. Penninckx, M. Merzouki and M. Benlemlih, 2005. Production of tannase by Aspergillus niger HA37 growing on tannic acid and olive mill waste waters. World J. Microbiol. Biotechnol., 21: 609-614.
CrossRefDirect Link - Aoki, K., R. Shinke and H. Nishira, 1976. Purification and some properties of yeast tannase. Agric. Biol. Chem., 40: 79-85.
Direct Link - Ayed, L. and M. Hamdi, 2002. Culture conditions of tannase production by Lactobacillus plantarum. Biotechnol. Lett., 24: 1763-1765.
CrossRefDirect Link - Bajpai, B. and S. Patil, 1997. Introduction of tannin acyl hydrolase activity in some members of fungi imperfecti. Enzyme Microbiol. Technol., 20: 612-614.
CrossRef - Banerjee, D., K.C. Mondal and B.R. Pati, 2001. Production and characterization of extracellular and intracellular tannase from newly isolated Aspergillus aculeatus DBF 9. J. Basic Microbiol., 41: 313-318.
CrossRefPubMedDirect Link - Banerjee, D., S. Mahapatra and B.R. Pati, 2007. Gallic acid production by submerged fermentation of Aspergillus aculeatus DBF9. Res. J. Microbiol., 2: 462-468.
CrossRefDirect Link - Batra, A. and R.K. Saxena, 2005. Potential tannase producers from the genera Aspergillus and Penicillium. Process Biochem., 40: 1553-1557.
CrossRefDirect Link - Battestin, V. and G.A. Macedo, 2007. Tannase production by Paecilomyces variotii. Bioresour. Technol., 98: 1832-1837.
PubMedDirect Link - Belmares, R., J.C. Conttreras-Esquivel, R. Rodriguez-Herrera, A.R. Coronel and C.N. Aguilar, 2004. Microbial production of tannase: An enzyme with potential use in food industry. Lebensm. Wiss. Technol., 37: 857-864.
CrossRefDirect Link - Belur, P.D., G. Mugeraya, K.R. Nirmala and N. Basavaraj, 2010. Production of novel cell-associated tannase from newly isolated Serratia ficaria DTC. J. Microbiol. Biotechnol., 20: 722-726.
PubMed - Belur, P.D., G. Mugeraya and B. Nainegali, 2011. Release of cell-associated tannase of Serratia ficaria DTC by sonication, surfactants and solvents. Asian J. Biotechnol., 3: 91-97.
CrossRefDirect Link - Beniwal, V. and V. Chhokar, 2010. Statistical optimization of culture conditions for tannase production by Aspergillus awamori MTCC 9299 under submerged fermentation. Asian J. Biotechnol., 2: 46-52.
CrossRefDirect Link - Beverini, M. and M. Metche, 1990. Identification, purification and physicochemical properties of tannase of Aspergillus oryzae. Sci. Aliments, 10: 807-816.
Direct Link - Bhat, T.K., B. Singh and O.P. Sharma, 1998. Microbial degradation of tannins-a current perspective. Biodegradation, 25: 343-357.
CrossRefPubMedDirect Link - Boadi, D.K. and R.J. Neufeld, 2001. Encapsulation of tannase for the hydrolysis of tea tannins. Enzyme Microb. Technol., 28: 590-595.
CrossRef - Bradoo, S., R. Gupta and R.K. Saxena, 1997. Parametric optimization and biochemical regulation of extracellular tannase from Aspergillus japonicus. Process Biochem., 32: 135-139.
CrossRefDirect Link - Das Mohapatra, P.K., K.C. Mondal and B.R. Pati, 2006. Production of tannase through submerged fermentation of tannin containing plant extracts by Bacillus lichiniformis KBR6. Polish J. Microbiol., 55: 297-301.
PubMedDirect Link - Deschamps, A.M., G. Otuk and J.M. Lebeault, 1983. Production of tannase and degradation of chestnut tannin by bacteria. J. Ferment. Technol., 61: 55-59.
Direct Link - Goel, G., A.K. Puniya, C.N. Aguilar and K. Singh, 2005. Interaction of gut microflora with tannins in feeds. Naturwissenschaften, 92: 497-503.
CrossRefDirect Link - Guzman-Lopez, O., O. Loera, J.L. Parada, A. Castillo-Morales and C. Martinez-Ramirez et al., 2009. Microcultures of lactic acid bacteria: Characterization and selection of strains, optimization of nutrients and gallic acid concentration. J. Ind. Microbiol. Biotechnol., 36: 11-20.
PubMedDirect Link - Haslam, E. and J.E. Stangroom, 1966. The esterase and depsidase activities of tannase. J. Biochem., 99: 28-31.
Direct Link - Hatamoto, O., T. Watarai, M. Kikuchi, K. Mizusawa and H. Sekine, 1996. Cloning and sequencing of the gene encoding tannase and a structural study of the tannase subunit from Aspergillus oryzae. Gene, 175: 215-221.
PubMedDirect Link - Jean, D., H. Pourrat, A. Pourrat and A. Carnat, 1981. Assay of tannase (tannin acyl hydrolase EC 3.1.1.20) by gas chromatography. Anal. Chem., 110: 369-372.
CrossRef - Kar, B. and R. Banerjee, 2000. Biosynthesis of tannin acyl hydrolase from tannin-rich forest residue under different fermentation conditions. J. Ind. Microbiol. Biotechnol., 25: 29-38.
CrossRefDirect Link - Kar, B., R. Banerjee and B.C. Bhattacharyya, 2003. Effect of additives on the behavioral properties of tannin acyl hydrolase. Process Biochem., 38: 1285-1293.
CrossRefDirect Link - Kasieczka-Burnecka, M., K. Kuc, H. Kalinowska, M. Knap and M. Turkiewicz, 2007. Purification and characterization of two cold-adopted extracellular tannin acyl hydrolases from an Antarctic strain Verticillium sp. P9. Appl. Microbiol Biotechnol., 77: 77-89.
Direct Link - Kumar, R.A., P. Gunasekaran and M. Lakshmanan, 1999. Biodegradation of tannic acid by Citrobacter freundii isolated from a tannery effluent. J. Basic Microbiol., 39: 161-168.
PubMedDirect Link - Lekha, P.K. and B.K. Lonsane, 1997. Production and application of tannin acyl hydrolase: State of the art. Adv. Applied Microbiol., 44: 215-260.
PubMedDirect Link - Lu, M.J. and C. Chen, 2007. Enzymatic tannase treatment of green tea increases in vitro inhibitory activity against N-nitrosation of dimethylamine. Process Biochem., 42: 1285-1290.
CrossRefDirect Link - Lu, M.J., S.C. Chu, L. Yan and C. Chen, 2009. Effect of tannase treatment on protein-tannin aggregation and sensory attributes of green tea infusion. Food Sci. Technol., 42: 338-342.
CrossRefDirect Link - Mahendran, B., N. Raman and D.J. Kim, 2006. Purification and characterization of tannase from Paecilomyces variotii: Hydrolysis of tannic acid using immobilized tannase. Applied Microbiol. Biotechnol., 70: 444-450.
CrossRef - Mondal, K.C., R. Banerjee and B.R. Pati, 2000. Tannase production by Bacillus licheniformis. Biotechnol. Lett., 22: 767-769.
CrossRefDirect Link - Mondal, K.C. and B.R. Pati, 2000. Studies on the extracellular tannase from newly isolated Bacillus licheniformis KBR6. J. Basic Microbiol., 40: 223-232.
Direct Link - Murugan, K., S. Saravanababu and M. Arunachalam, 2007. Screeing of tannin acyl hydrolase (E.C.3.1.1.20) producing tannery effluent fungal isolates using simple agar plate and SmF process. Bioresour. Technol., 98: 946-949.
CrossRef - Nishitani, Y., E. Sasaki, T. Fujisawa and R. Osawa, 2004. Genotypic analyses of Lactobacilli with a range of tannase activities isolated from human feces and fermented foods. Syst. Applied Microbiol., 27: 109-117.
CrossRef - Osawa, R. and T.P. Walsh, 1993. Visual reading method for detection of bacterial tannase. Appl. Environ. Microbiol., 59: 1251-1252.
PubMedDirect Link - Osawa, R., K. Kuroiso, S. Goto and A. Shimizu, 2000. Isolation of tannin degrading Lactobacilli from humans and fermented foods. Applied Environ. Microbiol., 66: 3093-3097.
Direct Link - Pinto, G.A.S., S.G.F. Leite, S.C. Terzi and S. Couri, 2001. Selection of tannase-producing Aspergillus niger strains. Braz. J. Microbiol., 32: 24-26.
CrossRefDirect Link - Pourrat, H., F. Regerat, A. Pourrat and D. Jean, 1982. Production of tannase (tannin acyl hydrolase E.C. 3.1.1.20) by a strain of Aspergillus niger. Biotechnol. Lett., 4: 583-588.
CrossRefDirect Link - Purohit, J.S., J.R. Dutta, R.K. Nanda and R. Banerjee, 2006. Strain improvement for tannase production from co-culture of Aspergillus foetidus and Rhizopus oryzae. Bioresour. Technol., 97: 795-801.
CrossRef - Rajkumar, G.S. and S.C. Nandy, 1983. Isolation, purification and some properties of Penicillium chrysogenum tannase. Applied Environ. Microbiol., 46: 525-527.
Direct Link - Ramirez-Coronel, M.A., G. Viniegra-Gonzalez, A. Darvill and C. Augur, 2003. A novel tannase from Aspergillus niger with β-glucosidase activity. Microbiology, 149: 2941-2946.
CrossRefDirect Link - Rout, S. and R. Banerjee, 2006. Production of tannase under mSSF and its application in fruit juice debittering. Ind. J. Biotechnol., 5: 346-350.
Direct Link - Sabu, A., G.S. Kiran and A. Pandey, 2005. Purification and characterization of tannin acyl hydrolase from Aspergillus niger ATCC 16620. Food Technol. Biotechnol., 43: 133-138.
Direct Link - Sabu, A., C. Augur, C. Swati and A. Pandey, 2006. Tannase production by Lactobacillus sp. ASR-S1 under solid-state fermentation. Process Biochem., 41: 575-580.
CrossRef - Saxena, S. and R.K. Saxena, 2004. Statistical optimization of tannase production from Penicillium variable using fruits (chebulic myrobalan) of Terminalia chebula. Biotechnol. Applied Biochem., 39: 99-106.
PubMed - Selwal, M.K., A. Yadav, K.K. Selwal, N.K. Aggarwal, R. Gupta and S.K. Gautam, 2010. Optimization of culture conditions for tannase production by Pseudomonas aeruginosa III B 8914 under submerged fermentation. Wolrld J. Micrbiol. Biotechnol., 26: 599-605.
CrossRefDirect Link - Seth, M. and S. Chand, 2000. Biosynthesis of tannase and hydrolysis of tannins to gallic acid by Aspergillus awamori-optimisation of process parameters. Process Biochem., 36: 39-44.
CrossRefDirect Link - Sharma, S., T.K. Bhat and R.K. Dawra, 2000. A spectrophotometric method for assay of tannase using rhodanine. Anal. Biochem., 279: 85-89.
CrossRefPubMedDirect Link - Sharma, S., T.K. Bhat and M.N. Gupta, 2002. Bioaffinity immobilization of tannase from Aspergillus niger on concanavalin A-Sepharose CL-4B. Biotechnol. Applied Biochem., 35: 165-169.
PubMed - Sharma, S., L. Agarwal and R.K. Saxena, 2008. Purification, immobilization and characterization of tannase from Penicillium variable. Bioresour. Technol., 99: 2544-2551.
CrossRef - Smith, A.H., E. Zoetendal and R.I. Mackie, 2005. Bacterial mechanism to overcome inhibitory effects of dietary tannins. Microb. Ecol., 50: 197-205.
PubMed - Van de Lagemaat, J. and D.L. Pyle, 2001. Solid state fermentation and bioremediation: Development of continuous process for the production of fungal tannase. Chem. Eng. J., 84: 115-123.
CrossRef - Yamada, H., O. Adachi, M. Watanabe and N. Sato, 1968. Studies on fungal tannase: Formation, purification and catalytic properties of tannase of Aspergillus flavus. Agric. Biol. Chem., 32: 1070-1078.
Direct Link - Yu, X., Y. Li, C. Wang and D. Wu, 2004. Immobilization of Aspergillus niger tannase by microencapsulation and its kinetic characteristics. Biotechnol. Applied Biochem., 40: 151-155.
PubMed - Mondal, K.C., D. Banerjee, R. Banerjee and B.R. Pati, 2001. Production and characterization of tannase from Bacillus cereus KBR9. J. Gen. Applied Microbiol., 47: 263-267.
CrossRefPubMedDirect Link - Lekha, P.K. and B.K. Lonsane, 1994. Comparative titres, location and properties of tannin acyl hydrolase produced by Aspergillus niger PKL 104 in solid-state, liquid surface an submerged fermentations. Process Biochem., 29: 497-503.
CrossRef