
H.A. Murad
Department of Dairy Science, National Research Center, Dokki, Cairo, Egypt
H.H. Azzaz
Department of Dairy Science, National Research Center, Dokki, Cairo, Egypt
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 246-269







ABSTRACT
Pectinases are widely distributed in higher plants and microorganisms. They are today one of the upcoming enzymes of the commercial sector. It has been estimated that microbial pectinases account for 25% of the global food enzymes sales. Microbial pectinases can be produced from bacteria including actinomycetes, yeast and fungi. Aspergillus niger is the most commonly used fungal species for industrial production of pectinolytic enzymes. Among industrial applications of pectinases are using these enzymes as an animal feed supplementation. This usage of pectinases for ruminant's feed production can reduces the feed viscosity, which increases absorption of nutrients, liberates nutrients, either by hydrolysis of non-biodegradable fibers or by liberating nutrients blocked by these fibers and reduces the amount of faeces. This study deals with types and classification of pectinolytic enzymes, their mode of action, production techniques and the methods of activity assay. Furthermore, it provides a bird’s eye view of the possible applications of these enzymes in ruminant nutrition.
PDF Abstract XML References Citation
Received: June 01, 2010;
Accepted: July 25, 2010;
Published: November 04, 2010
How to cite this article
H.A. Murad and H.H. Azzaz, 2011. Microbial Pectinases and Ruminant Nutrition. Research Journal of Microbiology, 6: 246-269.
URL: https://scialert.net/abstract/?doi=jm.2011.246.269
URL: https://scialert.net/abstract/?doi=jm.2011.246.269
INTRODUCTION
Pectinases or pectinolytic enzymes are today one of the upcoming enzymes of the commercial sector. It has been reported that microbial pectinases account for 25% of the global food enzymes sales (Jayani et al., 2005). Primarily, these enzymes are a heterogeneous group of related enzymes that hydrolyze the pectic substances, present mostly in plants.
Pectinolytic enzymes are widely distributed in higher plants and microorganisms (Whitaker, 1990). They are of prime importance for plants as they help in cell wall extension (Ward and Moo-Young, 1989) and softening of some plant tissues during maturation and storage (Sakai, 1992; Aguilar and Huirton, 1990). They also aid in maintaining ecological balance by causing decomposition and recycling of waste plant materials (Jayani et al., 2005).
Microbial production of pectinases has been extensively studied during recent years (Kashyap et al., 2001; Torres et al., 2006). Pectinase production has been reported from bacteria including actinomycetes (Cao et al., 1992; Bruhlmann et al., 1994; Beg et al., 2000), yeast (Blanco et al., 1999; Reid and Ricard, 2000) and fungi (Elegado and Fujio, 1994; Huang and Mahoney, 1999). However, almost all the commercial preparations of pectinases are produced from fungal sources (Singh et al., 1999). Aspergillus niger is the most commonly used fungal species for industrial production of pectinolytic enzymes (Naidu and Panda, 1998; Gummadi and Panda, 2003).
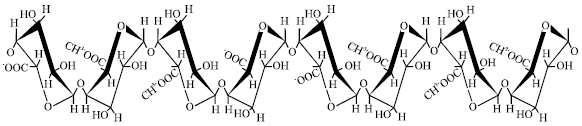 | |
| Fig. 1: | Primary structure of pectic substances (Rexova -Benkova and Markovic , 1976) |
Higher cost of the production is perhaps the major constraint in commercialization of new sources of enzymes. Though, using high yielding strains, optimal fermentation conditions and cheap raw materials as a carbon source can reduce the cost of enzyme production for subsequent applications in industrial processes. Among industrial applications of pectinases are using these enzymes as an animal feed supplementation. This usage of pectinases for ruminant's feed production can reduces the feed viscosity, which increases absorption of nutrients, liberates nutrients, either by hydrolysis of non-biodegradable fibers or by liberating nutrients blocked by these fibers and reduces the amount of faeces (Hoondal et al., 2002).
This study mainly concentrates on the types and classification of pectinolytic enzymes, their mode of action, production techniques and the methods of activity assay. It also provides a bird’s eye view of the possible applications of these enzymes in ruminant nutrition.
THE SUBSTRATE
Pectic substance is the generic name used for the compounds that are acted upon by the pectinolytic enzymes. They are high molecular weight, negatively charged, acidic, complex glycosidic macromolecules (polysaccharides) that are present in the plant kingdom. They are present as the major components of middle lamella between the cells in the form of calcium pectate and magnesium pectate (Jayani et al., 2005).
Chemically, pectic substances are essentially branched heteropolysaccharides containing between a few hundred and about one thousand building blocks per molecule, with a backbone consisting of galacturonic acid residues part of which are methylesteritied. The pectin molecule (Fig. 1) is generally agreed to consist of a chain structure of axial-axial E-(1, 4)-linked D-galacturonic acid units, containing blocks of I.-rhamnosc rich regions, with mainly arabinose, galactose and xylose as side chains. The carboxyl groups of the galacturonic acid are partly esterified by methyl groups and partly or completely neutralized by sodium, potassium or ammonium ions. Some of the hydroxyl groups on C2 and C3 may be acetylated (Pilnik and Voragen, 1970).
The American Chemical Society classified pectic substances into four main types as reported by Alkorta et al. (1998) as follows:
| • | Protopectin is the water insoluble pectic substance present in intact tissue. Protopectin on restricted hydrolysis yields pectin or pectic acids |
| • | Pectic acid is the soluble polymer of galacturonans that contains negligible amount of methoxyl groups Normal or acid salts of pectic acid are called pectates |
| • | Pectinic acids is the polygalacturonan chain that contains >0 and <75% methylated galacturonate units. Normal or acid salts of pectinic acid are referred to as pectinates |
| • | Pectin (Polymethyl galacturonate) is the polymeric material in which, at least, 75% of the carboxyl groups of the galacturonate units are esterified with methanol. It confers rigidity on cell wall when it is bound to cellulose in the cell wall |
THE PECTINASES
The pectinases may be divided in three broader groups as reported by Sakai (1992) and Palomaki and Saarilahti (1997) as follows:
| • | Protopectinases: Degrade the insoluble protopectin and give rise to highly polymerized soluble pectin |
| • | Esterases: Catalyze the de-esterification of pectin by the removal of methoxy esters |
| • | Depolymerases: Catalyze the hydrolytic cleavage of the α-(1-4)-glycosidic bonds in the D-galacturonic acid moieties of the pectic substances |
Depolymerases act on pectic substances by two different mechanisms, hydrolysis, in which they catalyze the hydrolytic cleavage with the introduction of water across the oxygen bridge and trans-elimination lysis, in which they break the glycosidic bond by a trans-elimination reaction without any participation of water molecule (Fig. 2) (Albersheim et al., 1960; Codner, 2001). Depolymerases can be subdivided into four different categories, depending on the preference of enzyme for the substrate, the mechanism of cleavage and the splitting of the glycosidic bonds (Rexova -Benkova and Markovic , 1976).
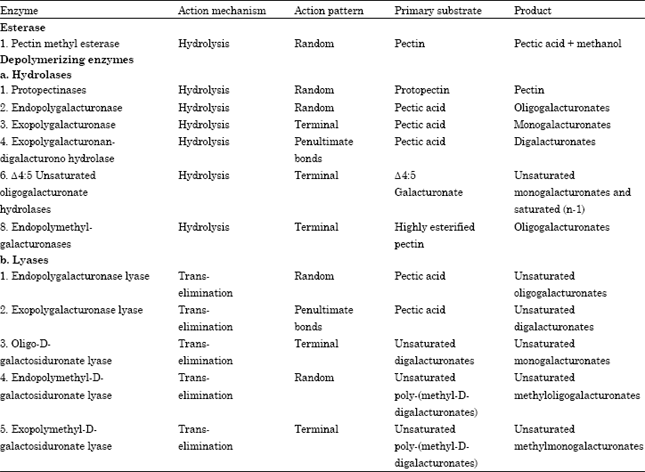
Polygalacturonase and polymethylgalacturonase breakdown pectate and pectin, respectively by the mechanism of hydrolysis. However, polygalacturonate lyase and polymethylgalacturonate lyase breakdown pectate and pectin by β elimination, respectively. Depending upon the pattern of action, i.e., random or terminal, these enzymes are termed as Endo or Exo enzymes, respectively. During the course of investigations and research conducted in recent years, the above classification of pectinolytic enzymes has slowly become obsolete. A more recent and elaborated classification of these enzymes has been provided in Table 1 (Rexova -Benkova and Markovic , 1976; Ward and Moo-Young, 1989; Whitaker, 1990; Kashyap et al., 2001).
Protopectinases: The enzyme that catalyzes the solubilization of protopectin was originally named protopectinase by Brinton et al. (1927). Pectinosinase is also synonymous with protopectinase (PPase). Protopectinase catalyzes the following reaction:
PPases are classified into two types, on the basis of their reaction mechanism (Sakai and Okushima, 1982; Sakamoto et al., 1994). A-type PPases react with the inner site, i.e., the polygalacturonic acid region of protopectin, whereas B-type PPases react on the outer site, i.e., on the polysaccharide chains that may connect the polygalacturonic acid chain and cell wall constituents.
A-type PPase are found in the culture filtrates of yeast and yeast-like fungi. They have been isolated from Kluyveromyces fragilis IFO 0288, Galactomyces reesei L. and Trichosporon penicillatum SNO 3 and are referred to as PPase-F, -L and -S, respectively (Whitaker,1990). B-type PPases have been reported in Bacillus subtilis IFO 12113 (Sakai and Ozaki, 1988), B. subtilis IFO3134 (Sakai and Sakamoto, 1990) and Trametes sp. (Sakai et al., 1993) and are referred to as PPase-B, -C and -T, respectively. B-type PPases have also been found in the culture filtrate of a wide range of Bacillus sp. (Sakai, 1992).
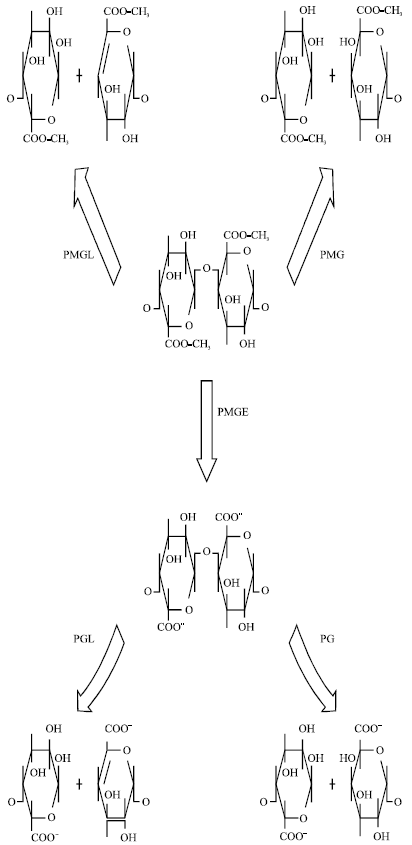 | |
| Fig. 2: | Enzymatic mode of action of the most frequent depolymerases on the pectin molecule. PMGL: Polymethylgalacturonate lyase (pectin lyase); PMG: Polymethylgalacturonase (pectin hydrolase); PMGE: Polymethylgalacturonate esterase (pectinestcrase); PGL: Polygalacturonate lyase (pectate lyase); PC: Polygalacturonase (pectate hydrolase). (Serra et al., 1992) |
Ppase activity is assayed by measuring the amount of pectic substance liberated from protopectin by the carbazole-sulphuric acid method (Siebert and Anto, 1946). The pectin concentration is measured as D-galacturonic acid from its standard curve. One unit of PPase activity is defined as the enzyme that liberates pectic substance corresponding to 1 mmol of Dgalacturonic acid per millilitre of reaction mixture under assay conditions.
Polygalacturonases: Polygalacturonases (PGases) are the pectinolytic enzymes that catalyze the hydrolytic cleavage of the polygalacturonic acid chain with the introduction of water across the oxygen bridge. They are the most extensively studied among the family of pectinolytic enzymes. The PGases involved in the hydrolysis of pectic substances are endo-PGase (E.C. 3.2.1.15) and exo-PGase (E.C. 3.2.1.67) (Table 1). PGases have the biological, functional and technical applications in food processing and plant-fungal interactions.
Endo-PGases are widely distributed among fungi, bacteria and many yeasts (Luh and Phaff, 1951). They are also found in higher plants and some plant parasitic nematodes (Sakai et al., 1993). They have been reported in many microorganisms, including Aureobasidium pullulans (Sakai and Takaoka, 1984), Rhizoctonia solani Kuhn (Marcus et al., 1986), Fusarium moniliforme (De Lorenzo et al., 1987), Neurospora crassa (Polizeli et al., 1991), Rhizopus stolonifer (Manachini et al., 1987), Aspergillus sp. (Nagai et al., 2000), Thermomyces lanuginosus (Kumar and Palanivelu, 1999), Peacilomyces clavisporus (Souza et al., 2003). Endo-PGases have also been cloned and genetically studied in a large number of microbial species (Laing and Pretorius, 1993; Naumov et al., 2001). In contrast, exo-PGases occur less frequently. They have been reported in Erwinia carotovora (Palomaki and Saarilahti, 1997), Agrobacterium tumefaciens (Rodrigues-Palenzuela et al., 1991), Bacteroides thetaiotamicron (Tierny et al., 1994), E. chrysanthemi (Kobayashi, et al., 2001), Alternaria mali (Nozaki et al., 1997), Fusarium oxysporum (Maceira et al., 1997), Ralstonia solanacearum (Huang and Allen, 1997), Bacillus sp. (Kobayashi et al., 2001). Exo-PGases can be distinguished into two types: fungal exo-PGases, which produce monogalacturonic acid as the main end product and the bacterial exo-PGases, which produce digalacturonic acid as the main end product (Sakai et al., 1993). Occurrence of PGases in plants has also been reported (Pressey and Avants, 1973; Alonso et al., 2003).
PGase activity is determined on the basis of measuring, during the course of the reaction: (1) the rate of increase in number of reducing groups and (2) the decrease in viscosity of the substrate solution (Rexova -Benkova and Markovic , 1976). The amount of reducing sugar can be readily measured by colorimetric methods like 3, 5-dinitrosalicylate reagent method (Miller, 1959) and the arsenomolybdate-copper reagent method (Nelson, 1944; Collmer et al., 1988). One unit of enzyme activity is defined as the enzyme that releases 1 mmol/mL/min galacturonic acid under standard assay conditions.
Viscosity reduction measurements have also found widespread use in determining the PGase activity (Roboz et al., 1952):
| Table 1: | An extensive classification of pectinolytic enzymes (Jayani et al., 2005) |
 | |
where, To, Tt and Tw represent the flow rate (in a capillary viscometer) in seconds for the reaction mixture without enzyme, the test mixture and water, respectively. The unit of enzyme activity is mostly selected as the amount of enzyme required for attaining a certain decrease of viscosity per unit time. However, this method has met with limited success. There is no direct correlation between viscosity reduction and number of glycosidic bonds hydrolyzed. PGase activity can also be determined by the cup-plate method (Dingle et al., 1953). Cups are cut out from the solidified agar containing the substrate and are filled with the enzyme solution. After lapse of a certain period of time, the zones of degraded substrate are stained with iodine and quantified.
Lyases: Lyases (or transeliminases) perform non-hydrolytic breakdown of pectates or pectinates, characterized by a trans-eliminative split of the pectic polymer (Sakai et al., 1993). The lyases break the glycosidic linkages at C-4 and simultaneously eliminate H from C-5, producing a D 4:5 unsaturated products (Codner, 2001; Albersheim et al., 1960).
Lyases can be classified into following types on the basis of the pattern of action and the substrate acted upon by them (Table 1):
(I) Endopolygalacturonate lyase (EndoPGL, E.C. 4.2.2.2); (II) exopolygalacturonate lyase (ExoPGL, E.C. 4.2.2.9); (III) endopolymethylgalacturonate lyase (EndoPMGL, E.C. 4.2.2.10); (IV) exopolymethylgalacturonate lyase (ExoPMGL).
The reactions catalyzed by lyases can be illustrated as follows:
Polygalacturonate lyases (Pectate lyases or PGLs) are produced by many bacteria and some pathogenic fungi with endo-PGLs being more abundant than exo-PGLs. PGLs have been isolated from bacteria and fungi associated with food spoilage and soft rot. They have been reported in Colletotrichum lindemuthionum (Wijesundera et al., 1984), Bacteroides thetaiotaomicron (McCarthy et al., 1985), Erwinia carotovora (Kotoujansky, 1987), Amucala sp. (Bruhlmann, 1995), Pseudomonas syringae pv. Glycinea (Margo et al., 1994), Colletotrichum magna (Wattad et al., 1995), E. chrysanthemi (Favey et al., 1992; Shevchik et al., 1997), Bacillus sp. (Kobayashi et al., 1999; Takao et al., 2001), Bacillus sp. DT-7 (Kashyap et al., 2000), C. gloeosporioides (Yakoby et al., 2000; Drori et al., 2003)
Very few reports on the production of polymethylgalacturonate lyases (pectin lyases or PMGLs) have been reported in literature. They have been reported to be produced by Aspergillus japonicus (Ishii and Yokotsuka, 1975), Penicillium paxilli (Szajer and Szajer, 1982, 1985), Penicillium sp. (Diaz Borras et al., 1987; Alana et al., 1990; Sapunova et al., 1995), Pythium splendens (Chen et al., 1998) Pichia pinus (Moharib et al., 2000), Aspergillus sp. (Sunnotel and Nigam, 2002), Thermoascus auratniacus (Martins et al., 2002).
The most convenient method of following the activity of lyases is to measure the increase in absorbance at 235 nm due to formation of the Δ4:5 double bonds produced at the non-reducing ends of the unsaturated products (Albersheim, 1966; Whitaker, 1990; Liao et al., 1999). The molar extinction coefficients for PGL and PMGL are 4.6x103 and 5.5x103/M/cm, respectively. One unit of enzyme activity is defined as the amount of enzyme that releases 1 mmol of unsaturated product per minute under assay conditions. Reducing group methods (Miller, 1959; Collmer et al., 1988) are also useful in determining the lyase activity. Viscosity reduction method (Roboz et al., 1952), in conjunction with a reducing group method or along with intermediate product analysis by HPLC or GC, can be used to distinguish between endo-and exo-splitting enzymes (Albersheim, 1966).
Pectinesterase: Pectinesterase (PE, Pectin pectlylhydrolase, E.C. 3.1.1.11), often referred to as pectinmethylesterase, pectase, pectin methoxylase, pectin demethoxylase and pectolipase, is a carboxylic acid esterase and belongs to the hydrolase group of enzymes (Whitaker, 1984). It catalyzes the deesterification of methyl ester linkages of galacturonan backbone of pectic substances to release acidic pectins and methanol (Cosgrove, 1997). The resulting pectin is then acted upon by polygalacturonases and lyases (Sakai et al., 1993; Prade et al., 1999). The mode of action of PE varies according to its origin (Micheli, 2001). Fungal PEs act by a multi-chain mechanism, removing the methyl groups at random. In contrast, plant PEs tends to act either at the non-reducing end or next to a free carboxyl group and proceed along the molecule by a single chain mechanism. The reaction catalyzed by PE can be represented as follows (Forster, 1988):
PE activity is implicated in cell wall metabolism including cell growth, fruit ripening, abscission, senescence and pathogenesis (Gaffe et al., 1997; Dorokhov et al., 1999). Commercially PE can be used for protecting and improving the texture and firmness of several processed fruits and vegetables as well as in the extraction and clarification of fruit juices (Fayyaz et al., 1993). PE is found in plants, plant pathogenic bacteria and fungi (Hasunuma et al., 2003). It has been reported in Rhodotorula sp. (Vaughan et al., 1969), Phytophthora infestans (Forster, 1988), Erwinia chrysanthemi B341 (Pitkanen et al., 1992), Saccharomyces cerevisiae (Gainvors et al., 1994), Lachnospira pectinoschiza (Cornick et al., 1994), Pseudomonas solanacearum (Schell et al., 1994), Aspergillus niger (Maldonado et al., 1994; Maldonado and de Saad,1998), Lactobacillus lactis subsp. Cremoris (Karam and Belarbi, 1995), Penicillium frequentans (Kawano et al., 1999), E. chrysanthemi 3604 (Laurent et al., 2000), Penicillium occitanis (Hadj-Taieb et al., 2002), A. japonicus (Semenova et al., 2003) and others. There are many reports of occurrence of PE in plants, viz., Carica papaya (Fayyaz et al., 1993; Innocenzo and Lajalo, 2001), Lycopersicum esculentum (Warrilow et al., 1994), Prunus malus (Macdonald and Evans, 1996), Vitis vinifera (Corredig et al., 2000), Citrus sp. (Arias and Burns, 2002), Pouteria sapota (Arenas-Ocampo et al., 2003) and Malpighia glabra L. (Assis et al., 2004).
PE activity is most readily followed by gel diffusion assay, as described by Downie et al. (1998). Increased binding of ruthenium red to pectin, as the number of methyl esters attached to the pectin decreases, is used as the basis of the assay. The unit of activity in nano-or picokatals is calculated, based on the standard curve generated from the log-transformed commercial enzyme activity versus stained zone diameter. The sensitivity, specificity and simplicity of this PE assay are superior to all others. PE activity can also be measured by using a pH stat because ionization of the carboxyl group of the product releases a proton, which causes a change in pH (Whitaker, 1984).
MICROBIAL PECTINASES PRODUCTION
In developing nations, there is noticeable growth of food and feed processing industries, leading to an increase in the demand for pectic enzymes. Unfortunately, many of these nations depend largely on imported pectinase, which has become more expensive because of higher cost of foreign exchange. Therefore, with a view of improving the utilization of horticultural product in the depressed economy of a nation like Egypt, it is necessary to carry out research on the production of pectic enzymes locally.
Microbial pectinases production techniques: There were two fermentation techniques we can use for pectinases production, as many other enzymes (Murad and Foda, 1992) these techniques are Solid State Fermentation (SSF) and submerged fermentation (SmF).
Solid state fermentation is defined as the cultivation of microorganisms on moist solid supports, either on inert carriers or on insoluble substrates that can be used as carbon and energy source. This process occurs in the absence or near absence of free water in the space between substrate particles. In this system, water is present in the solid substrate whose capacity for liquid retention varies with the type of material (Lonsane et al., 1985; Pandey et al., 2001). In contrast, in submerged fermentation (SmF) the nutrients and microorganisms are both submerged in water (Grigelmo-Migeul and Martin-Belloso, 1998).
Approximately 90% of all industrial enzymes are produced in SmF, frequently using specifically optimized, genetically manipulated microorganisms. In this respect SmF processing offers an insurmountable advantage over SSF. On the other hand, almost all these enzymes could also be produced in SSF using wild-type microorganisms (Filer, 2001; Pandey et al., 2001). Interestingly, fungi, yeasts and bacteria that were tested in SSF in recent decades exhibited different metabolic strategies under conditions of solid state and submerged fermentation.
| Table 2: | Biotechnological advantages of Solid State Fermentation (SSF) against submerged technology (SmF) (Holker et al., 2004) |
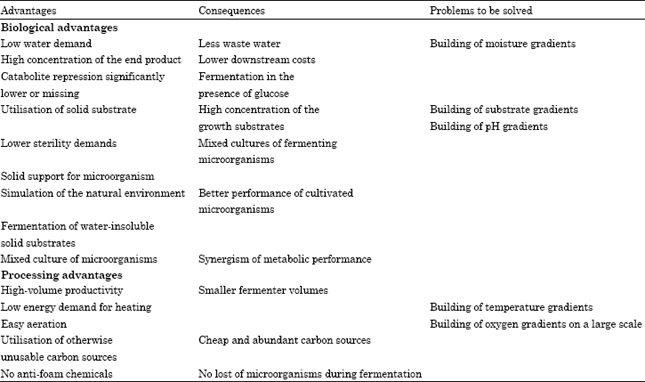 | |
The aim of SSF is to bring the cultivated fungi or bacteria into tight contact with the insoluble substrate and thus to achieve the highest substrate concentrations for fermentation. This technology results, only on a small scale, in several processing advantages of significant potential economic and ecological importance as compared with SmF (Table 2).
However, there are also several disadvantages of SSF, which have discouraged use of this technique for industrial production. The main obstructions are due mainly to the build-up of gradients of temperature, pH, moisture, substrate concentration or CO2 during cultivation, which are difficult to control under limited water availability. It has become clear (as mentioned in nearly every review cited) that the cost-factor for the production of bulk-ware enzymes in most cases favors SSF over SmF. The low estimated costs of SSF are due to the rather traditional preferential claim of SSF, viz. SSF utilises complex, heterogenous agricultural wastes as substrates and uses low-cost technology regarding sterility and regulation demands. However, attempts to reduce costs progress in SSF because of the strongly increased diversity in SSF research.
There is no consensus on the methods, the microorganisms or the substrates used, that would allow comparison with other cultivation technologies. The broad spectrum of substrates used represents an especially severe problem. As already mentioned, one great advantage of SSF has always been the possibility of using substrates that are abundant, cheap and not applicable to SmF. However, regardless of the differences in process up-scaling, the scientific and technological impact of research data is difficult to compare when results are obtained with different microorganisms producing different products and using a vast variety of substrates (Rafae et al., 2006; Abd El-Salam et al., 1994). There are many substrates could be used as, pineapple, mixed fruit, maosmi waste, orange bagasse, mango waste, corn tegument, almond meal, apple pomace, molasses, banana peels, amaranth grain, eucalyptus kraft pulp, coffee residues and some food industry wastes (Holker et al., 2004; Murad, 1998).
To facilitate comparison of results, the use of inert substrates as solid supports is becoming increasingly important (Gautam et al., 2002; Ooijkaas et al., 2000). Surprisingly, biological parameters, such as the stability of the produced enzymes at high temperature or extreme pH, have also been reported to be better in SSF (Deschamps and Huet, 1985; Acuna-Arguelles et al., 1995). Capability repression or protein degradation by proteases severe problems in SmF were often reduced or absent in SSF (Pereira et al., 1993; Aguilar et al., 2001). In contrast, much less research has been carried out to evaluate the metabolic differences of microorganisms when cultivated in SSF or SmF.
The factors affecting microbial pectinases production: Environmental and nutritional factors are known to have marked effects on enzyme production by microorganisms. There are, therefore, variations in optimum conditions for pectic enzyme production. Some of the cultural factors that affect the production of pectic enzymes are presented in this study.
Initial pH of growth medium: According to Shoichi et al. (1985) the initial pH of the medium has a great effect on the growth of the organism, on the membrane permeability, also on the biosynthesis and stability of the enzymes (Murad, 1998; Murad and Salem, 2001). Optimum production of pectic enzymes from many moulds has been reported to be within the acidic pH range (Zetelaki-Horvath, 1980; Shin et al., 1983). Zheng and Shetty (1999), reported that, polygalacturonase produced from Lentinus edodes has a relatively lower optimum pH (pH 5.0) in addition, Piccoli-Valle et al. (2001) observed that a high polygalacturonase and pectin esterase activity was showed by P. griseoroseum in more acid pH of 4.5 and 5. Also, Silva et al. (2002) found that P. viridicatum showed maximum production of polygalacturonase and pectinlyase at a pH of 4.5 and 5, respectively. Fawole and Odunfa (2003) reported that the optimum pectolytic activity observed was at pH 5. Phutela et al. (2005) concluded that the thermophilic fungi A. fumigatus Fres expressed maximum pectinase (1116 Ug-1) activity at pH 4.0 while polygalacturonase was active at pH 5.0 (1270 Ug-1). Also, Debing et al. (2005) found that the pH 6.5 was the optimal pH for pectinase production from A. niger by solid state fermentation. Reda et al. (2008) found that the polygalacturonase productivity by Bacillus firmus-I-10104 reached its maximum at initial pH 6.0 and 6.2. Rasheedha et al. (2010) found that P. chrysogenum exhibited maximum polygalacturonase production at initial pH of 6.5. However, the mechanism by which the pH acts on the production pectic enzyme is not known.
Incubation period: The time of fermentation had a profound effect on microbial product formation (Murad and Foda, 1992; Murad, 1998; Murad and Salem, 2001). Maximum production of pectic enzyme from different moulds varies from 1 to 6 days (Ghildyal et al., 1981). Castilho et al. (2000) reported that the highest polygalacturonase activities were obtained by A. niger after 70 h of fermentation period. In addition, Fawole and Odunfa (2003) reported that optimum production of pectinmethylesterase was obtained after 4 days of fermentation under submerged fermentation condition. Moreover, Sarvamangala and Dayanand (2006) observed a gradual increase in the production of pectinase from deseeded sunflower head by A. niger after 72 h of fermentation period in submerged and up to 96 h in solid-state conditions. Reda et al. (2008) found that the level of polygalacturonase increased gradually with increasing the incubation period up to a maximum of 96 h by Bacillus firmus-I-10104 under solid state fermentation conditions.
Nitrogen source: The effects of organic and inorganic nitrogen sources on the production of pectinase were extensively studied. The observations of Hours et al. (1988) suggested that lower levels of (NH4)2SO4 (0.16%), or K2HPO4 (0.1%) added to the growth medium as inorganic nitrogen sources did not influence pectinase yield. In addition Galiotou-Panayotou and Kapantai (1993) observed that ammonium phosphate and ammonium sulphate did influence production of pectinase positively but also recorded the inhibitory effects of ammonium nitrate and potassium nitrate on pectinase production. Moreover, Sarvamangala and Dayanand (2006) revealed that both ammonium phosphate and ammonium sulphate did influence production of pectinase positively in both submerged and solid-state conditions. In contrast, Sapunova (1990) found that ammonium salts stimulated the pectinolytic enzyme production in A. alliaceus BIM-83. Moreover, Sapunova et al. (1997) has also observed that (NH4)2SO4 stimulated pectinase synthesis, as in its absence fungus displayed a slight proteolytic activity and did not produce extracellular pectinases. In addition, Fawole and Odunfa (2003) found that ammonium sulphate and ammonium nitrate were good nitrogen sources for pectic enzyme production from A. niger while glycine and tryptophan did not support enzyme production. Also, Phutela et al. (2005) reported that (NH4)2SO4 stimulated pectinase production, as in its absence fungus displayed a slight proteolytic activity and did not produce extracellular pectinases. Phutela et al. (2005) found that the presence of yeast extract + (NH4)2 SO4 in growth medium supported maximal production of pectinase (925 Ug-1) followed by malt sprouts + (NH4)2SO4 (785 Ug-1), which also supported maximal polygalacturonase activity (938 Ug-1). In addition, Rasheedha et al. (2010) found that ammonium sulphate has enhanced the production of P. chrysogenum pectinase.
On the other hand, report of Aguilar et al. (1991) showed yeast extract (organic nitrogen source) as the best inducer of exopectinases by Aspergillus sp. Moreover Kashyap et al. (2003) found that, yeast extract, peptone and ammonium chloride were found to enhance pectinase production up to 24% and addition of glycine, urea and ammonium nitrate inhibited pectinase production. Also, Reda et al. (2008) found that the maximum value of polygalacturonase productivity by Bacillus firmus-I-10104 reached up to 350 U mL-1 in the presence of peptone as a nitrogen source in the growth medium. In addition, Vivek et al. (2010) found that organic nitrogen sources showed higher endo, exo pectinases activities than inorganic nitrogen sources. Also the increasing trend in the enzymes activity with the increase in nitrogen source content was observed in the case of organic nitrogen sources while decreasing trend observed for inorganic nitrogen sources. Soybean meal (4%) showed the maximum Exopectinase activity of 5128 IU g-1 and endo-pectinase activity of 793 IU g-1.
Carbon source: An adequate supply of carbon as energy source is critical for optimum growth affecting the growth of organism and its metabolism. Aguilar and Huitron (1987) reported that the production of pectic enzymes from many moulds is known to be enhanced by the presence of pectic substrates in the medium. Fawole and Odunfa (2003) found that pectin and polygalacturonic acid promoted the production of pectic enzyme and they observed the lack of pectolytic activity in cultures with glucose as sole carbon source reflects the inducible nature of pectic enzyme from the strain of A. niger. However, when different concentrations of glucose were added to the medium containing pectin, production of pectic enzymes was inhibited at high glucose concentration while low glucose concentrations (0.5% w/v) stimulated enzyme production Also, the reducing sugar content of the culture filtrate increased with increase in the amount of glucose added to the growth medium. The ability of high concentrations of glucose in the medium to meet growth requirement of the organism probably made the breakdown of pectin in the medium unnecessary or minimal and thus the low pectic activities observed in cultures. Phutela et al. (2005) stated that wheat bran supported maximum pectinase production (589 U g-1) while pure pectin give the maximum production of polygalacturonase (642 U g-1). Sarvamangala and Dayanand (2006) reported that glucose (4-6%) increase the production of pectinase in submerged condition whereas 6-8% sucrose gives better yield of pectinase in solid-state condition. Reda et al. (2008) reported that Solanum tuberosum (ST) peels was the best carbon source for polygalacturonase production by Bacillus firmus-I-10104 under solid state condition.
PURIFICATION OF MICROBIAL PECTINASES
In order to characterize and study the properties of microbial pectinases the enzymes must be purified. Important purification methods for the isolation of different pectinases are briefly summarized in this section. Pectinases from various sources of microorganisms have been purified to homogeneity. An exo-PG has been separated from mycelial extracts of Aspergillus niger by eluting from DEAE cellulose with 0.2 M sodium acetate buffer at pH 4.6. Purification was efficient with 209-fold increase in specific activity with a recovery of 8.6% and the enzyme displayed its full activity only in the presence of Hg2_ ions (Mill, 1966a). A second PG was isolated with 205-fold increase in specific activity with a recovery of 1%.These two PGs are differentiated by their optimum pH and PG II was not inhibited by chelating agents and did not require Hg2_ for activity (Mill, 1966b).
Benkova and Slezarik (1966) developed a purification strategy for the isolation of extracellular PMG, PG and PE. The enzyme was salted out with ammonium sulphate and precipitated with ethanol after gel filtration through Sephadex G-25. Repeated chromatography on DEAE-cellulose column yielded a homogeneous preparation of enzyme. Exo-PG, Endo-PG and pectinesterase have been separated from the culture filtrate of Trichoderma reesei by Sephadex chromatography (Markovic et al., 1985). Polygalacturonase from Rhizopus stolonifer has been purified up to 10-fold by ethanol precipitation followed by CM-Sepharose 6B ion exchange chromatography and gel filtration by Sephadex G-100 (Manachini et al., 1987). PG and PL (pectinlyase) from Aureobasidium pullulans LV10 have been separated by CMSepharose 6B followed by column chromatography (DEAE-cellulose column) and gel filtration on Sephadex G-100 (Manachini et al., 1988). PG and PL (pectinlyase) have been separated into PG I and PG II and PL I and PL II, respectively. Pectatelyase (PGL) was synthesized by Amycolata species and the extracellular crude enzyme has been purified to homogeneity by both cation and anion exchange columns and hydrophobic interaction chromatography (Bruhlmann, 1995). It has been observed that purification resulted in a 4-fold increase in specific activity with 37% recovery. Pectinases from Clostridium aectobutylicum ID 91-36 a UV mutant, has been purified by cation exchange chromatography on a Sepharose column by eluting with NaCl (Seethaler and Hartmeier, 1992).
Endopectate lyase synthesized by Bacillus macerans has been purified by ammonium sulphate precipitation followed by DEAE-Sephadex A-50 chromatography and CM-cellulofine chromatography (Miyazaki, 1991). Similarly endopectate lyase I_/IV have been isolated from the culture filtrate of Erwinia carotovora by CM Sepharose CL 6B chromatography, Sephadex S-200 gel filtration and isoelectric focusing (Tanabe et al., 1984). Kobayashi et al. (2001) purified the first bacterial; exo-PG from Bacillus sp. strain KSM-P443 to homogeneity. This enzyme releases exclusively mono-glacturonic acid from polygalacturonic acid (PGA), di-, tri-, tetra-and penta-galacturonic acids. They also determined the N-terminal sequence and concluded that no sequence matched with other pectinases reported to-date.
An extracelluar endo-PG produced by Aspergillus awamori IFO 4033 was purified homogeneity using cation-exchange and size-exclusion chromatographic columns (Nagai et al., 2000). Sakamoto et al. (1994) isolated protopectinase-N (PPN) and protopectinase-R (PPR) from the culture filtrate of Bacillus subtilis IFO3134. These enzymes have been purified by hydrophobic interaction chromatography on butyl-toyopearl 650 M, cation exchange chromatography on CMtoyopearl650 M and gel filtration on sepharose 12HR. These enzymes have been found to be stable over a wide range of pH and temperature. Endopectate lyase produced by Erwinia caratovara FERM P-7576 has been selectively co-sedimented with an extracellularly produced lipopolysaccharide lipid complex (Fukoka et al., 1990). The cell free broth was precipitated and the enzyme separated by gel chromatography with a specific activity of 710 U mg-1 of protein. Co-sedimentation has been affected by pH and ionic strength. Denis et al. (1990) studied the effect of shear stress on purification of five isozymes of pectate lyase produced by Erwinia chrysanthemi 3937 in ultrafiltration equipment. Activity was not affected during 7 h of pumping and 36% activity was lost after 25 000 passes.
New affinity matrices have been developed for the purification of pectinases, which possess better mechanical and chemical stability than those cross-linked one with pectic acid (Lobarzewski et al. 1985). The culture filtrate was desalted on a Sephadex G-25 column. The supports used were silanized controlled pore glass, silica gel silanized with 5-aminopropyl triethoxysilane. All supports were activated with 3-(3-dimethylaminopropyl) carbodiimide and best results were obtained with silanized controlled pore glass. Gupta et al. (1996) developed an affinity precipitation technique for separation of selective proteins using heterobifunctional ligands. They used a soluble form of the ligand for affinity binding and then precipitation was induced for separating the protein complex. Alginate was used as successful ligand for pectinases. Wu et al. (2000) studied the partitioning behavior of endo-PG and total protein from Kluveromyces marxianus culture broth in polyethylene glycol (PEG) and PEG-potassium phosphate aqueous two-phase systems (ATPS). Both enzyme and total protein partitioned into the bottom phase in both systems. Since, the enzyme protein content in the total protein of the fermentation broth was higher, they proposed that separating endo-PG was concentration rather than separation.
An efficient concentration was achieved using ATPS with polymer recycling and dialysis. Immobilized metal ion affinity polysulphone hollow-fiber membranes with a high capacity for protein adsorption were tested for commercial pectic enzyme fractionation (Camperi et al., 2000). The flow through (unbound) fraction has higher activity for pectinlyase and PE was retained on the column. Similarly, Savary (2001) developed a rapid and simple method to separate PE from PG and other pectinases in potato enzyme preparations using perfusion chromatography (Poros HS). PE was eluted at low salt concentrations (-/80 mM NaCl) and PG at high salt concentrations (-/300 mM NaCl). The development of such columns looks promising for economical purification strategies for pectinases. A literature survey showed that extensive work has been carried out on the purification of PG. PE and PGL and less attention has been focused on PMG protopectinases and oligogalacturonases.
RUMINANT AND MICROBIAL PECTINASES
Ruminants have an extensive array of microbial enzymes produced in the rumen and these enzymes play an important role in the ruminant digestive process. Pectinolytic activity in the rumen arises from protozoal, fungal and bacterial sources, one of the major pectinolytic bacterial species inhabiting the rumen, Lachnospira multiparus, produces a pectin lyase and a pectin methylesterase (Silley, 1985). Ruminal fungi and protozoa also express one or more of these enzymes (Orpin, 1984; Chesson and Forsberg, 1988; Bonhomme, 1990).
On the other hand, specific enzyme preparations have become valuable tools for economically improving digestive processes in the ruminant (Gado et al., 2009; Murad et al., 2009; Azzaz, 2009). Use supplemental enzymes in the rumen may be important since the digestibility of organic matter in the rumen does not reach 100% and even small changes in digestibility can influence the efficiency of ruminal fermentations. As a result, many strategies have been developed to stimulate the digestion of the fibrous components in ruminant feeds. One particularly promising technology is supplementation of fibrolytic enzymes (Cellulases, xylanases and pectinases) to ruminant's diets (Selinger et al., 1996). Although extensive studies were carried out to investigate the effects of cellulases and xylanases supplementation to ruminant diets on their productivity, study effects of pectinases supplementation to ruminant diets is lacking. Hoondal et al. (2002) reported that use of pectinases as ruminant's feed supplementation can reduces the feed viscosity, which increases absorption of nutrients, liberates nutrients, either by hydrolysis of non-biodegradable fibers or by liberating nutrients blocked by these fibers and reduces the amount of faeces. In addition, Jacob et al. (2008) reported that the reducing sugar level was increasing gradually for banana fiber treatment with polygalacturonase indicating the effectiveness of the treatment and it was evident from the photographs that the cells were separated after treatment as a result of pectin hydrolysis. This may be reflecting the importance of use pectinases as a fiber degrading enzymes in animal feed production.
To date, little is known about the way that exogenous fibrolytic enzymes improve feed by rumen microorganisms. Several potential modes of action have been proposed. These include: a) increase in microbial colonization of feed particles. Yang et al. (1999), enhancing attachment and/or improve access to the cell wall matrix by ruminal microorganisms and by doing so, accelerate the rate of digestion (Nsereko et al., 2000) enhancing the hydrolytic capacity of the rumen due to added enzyme activities and /or synergy with rumen microbial enzymes (Morgavi et al., 2000). Also to date, it has not been possible to predict the effectiveness of enzyme products for ruminants based on their enzymatic activities because the effectiveness of enzyme products is highly variable (Colombatto et al., 2003). Part of this variability may be due to the types and activities of enzyme, which can vary widely depending on the source organism, growth substrate and culture conditions employed, type of diet fed to animals, enzyme application method and even the level of animal productivity (Beauchemin et al., 2003). Lineweaver and Jansen (1951) study effect of low and high concentration of pectic substances (citrus pulp) on polygalacturonase activity in steer's rumen fluid (in vitro). They found that polygalacturonase activity in steer's rumen fluid increased by ten folds compared with basal diet and there was no significant difference in polygalacturonase activity between the low and high level of citrus pulp. Both levels reached about the same maximum activity of 2,000 units mL-1) and returned to about 200 units on the basal diet. This ten fold increase in activity of polygalacturonase activity attributed to citrus pulp suggests that the formation of extracellular polygalacturonase was induced by the increase in total pectic substances in the diet.
Finally, we recommended in this study giving more attention to production of microbial pectinases and more attention to their application especially in animal feed production.
REFERENCES
- Abd El-Salam, M.H., M.A. Fadel, H.A. Murad, 1994. Bioconversion of sugarcane molasses into xanthan gum. J. Biotechnol., 33: 103-106.
CrossRefDirect Link - Acuna-Arguelles, M.E., M. Gutierrez-Rojas, G. Viniegra-Gonzalez and E. Favela-Torres, 1995. Production and properties of three pectinolytic activities produced by Aspergillus niger in submerged and solid-state fermentation. Applied Microbiol. Biotechnol., 43: 808-814.
PubMedDirect Link - Aguilar, C.N., C. Augur, E. Favela-Torres and G. Viniegra-Gonzalez, 2001. Production of tannase by Aspergillus niger Aa-20 in submerged and solid-state fermentation: Influence of glucose and tannic acid. J. Ind. Microbiol. Biotechnol., 26: 296-302.
Direct Link - Albersheim, P., H. Neukom and H. Deuel, 1960. Splitting of pectin chain molecules in neutral solutions. Arch. Biochem. Biophysiol., 90: 46-51.
CrossRef - Alkorta, I., C. Garbisu, M.J. Llama and J.L. Serra, 1998. Industrial applications of pectic enzymes: A review. Process Biochem., 33: 21-28.
CrossRefDirect Link - Alonso, J., W. Canet, N. Hawell and R. Alique, 2003. Purification and characterization of carrot (Daucas carota L.) pectinesterase. J. Sci. Food. Agric., 83: 1600-1606.
CrossRef - Arenas-Ocampo, M.L., S. Evangelista-Lozano, R. Arana-Errasquin, A. Jimenez-Aparicio and G. Davila-Ortız, 2003. Softening and biochemical changes of sapote mamey fruit (Pouteria sapota) at different development and ripening stages. J. Food. Chem., 27: 91-107.
CrossRef - Arias, C. and J. Burns, 2002. A pectinmethylesterase gene associated with a heatstable extract from citrus. J. Agric. Food. Chem., 50: 3465-3472.
CrossRef - Assis, S.A., P. Fernandes, B.S. Ferreira, H.C. Trevisan, J.M.S. Cabral and O.M.M.F. Oliveira, 2004. Screening of supports for the immobilization of pectin-methylesterase from acerola (Malpighia glabra L.). J. Chem. Technol. Biotechnol., 79: 277-280.
Direct Link - Beauchemin, K.A., D. Colombatto, D.P. Morgavi and W.Z. Yang, 2003. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J. Anim. Sci., 81: E37-E47.
Direct Link - Beg, Q.K., B. Bhushan, M. Kapoor and G.S. Hoondal, 2000. Effect of amino acids on production of xylanase and pectinase from Streptomyces sp. QG-11-3. World J. Microbiol. Biotechnol., 16: 211-213.
CrossRefDirect Link - Blanco, P., C. Sieiro and T.G. Villa, 1999. Production of pectic enzymes in yeasts. FEMS Microbiol. Lett., 175: 1-9.
CrossRefDirect Link - Bonhomme, A., 1990. Rumen ciliates: their metabolism and relationships with bacteria and their hosts. Anim. Feed Sci. Technol., 30: 203-266.
Direct Link - Bruhlmann, F., 1995. Production and characterization of an extracellular pectate lyase from an Amycolata sp. Applied Environ. Microbiol., 61: 3580-3585.
Direct Link - Camperi, S.A., M. Grasselli and O. Cascone, 2000. High-speed pectic enzyme fractionation by immobilized metal ion affinity membranes. Bioseparation, 9: 173-177.
CrossRef - Cao, J., L. Zheng and S. Chen, 1992. Screening of pectinase producer from alkalophilic bacteria and study on its potential application in degumming of ramie. Enzyme Microbial Technol., 14: 1013-1016.
CrossRef - Chen, W.C., H.J. Hsieh and T.C. Tseng, 1998. Purification and characterization of a pectin lyase from Pythium splendens infected cucumber fruits. Botanical. Bull. Academia. Sinica, 39: 181-186.
Direct Link - Colombatto, D., D.P. Morgavi, A.F. Furtado and K.A. Beauchemin, 2003. Screening of exogenous enzymes for ruminant diet: relationship between biochemical characteristics and in vitro ruminal degradation. J. Anim. Sci., 81: 2628-2638.
Direct Link - Cornick, N.A., N.S. Jensen, D.A. Stahl, P.A. Hartman and M.J. Allison, 1994. Lachnospira pectinoschiza sp. nov., an anaerobic pectinophile from the pig intestine. Int. J. Syst. Bacteriol., 44: 87-93.
CrossRef - Corredig, M., W. Kerr and L. Wicker, 2000. Separation of thermostable pectinmethylesterase from marsh grapefruit pulp. J. Agric. Food. Chem., 48: 4918-4923.
CrossRefPubMedDirect Link - Cosgrove, D.J., 1997. Assembly and enlargement of the primary cell wall in plants. Ann. Rev. Cell Dev. Biol., 13: 171-201.
PubMed - De Lorenzo, G., G. Salvi, L. Degra, R. D`Ovidio and F. Cervone, 1987. Introduction of extracellular polygalacturonases and its mRNA in the phytopathogenic fungus Fusarium moniliforme. J. Gen. Microbiol., 133: 3365-3373.
Direct Link - Denis, S., S. Terre, Y. Bertheau and P. Boyaval, 1990. Factors affecting pectatelyase activity during membrane filtration. Biotechnol. Technol., 4: 127-132.
CrossRef - Deschamps, F. and M.C. Huet, 1985. Xylanase production in solid-state fermentation: A study of its properties. Applied Microbiol. Biotechnol., 22: 177-180.
Direct Link - Dingle, J., W.W. Reid and G.L. Solomons, 1953. The enzymatic degradation of pectin and other polysaccharides. II. Application of the `Cup-plate` assay to the estimation of enzymes. J. Sci. Food. Agric., 4: 149-155.
CrossRef - Dorokhov, Y.L., K. Mankinen, O.Y. Frolova, A. Mertis, J. Saarinen and N. Kalkkinen, 1999. A novel function for a ubiquitous plant enzyme pectin methylesterase: the host-cell receptor for the tobacco mosaic virus movement protein. FEBS Lett., 461: 233-238.
PubMed - Downie, B., L.M.A. Dirk, K.A. Hadfield, T.A. Wilkins, A.B. Bennet and K.J. Bradford, 1998. A gel diffusion assay for quantification of pectinmethylesterase activity. Anal. Biochem., 264: 149-157.
CrossRef - Drori, N., H. Kramer-Haimovich, J. Rollins, A. Dinoor, Y. Okon and O. Pines, 2003. External pH and nitrogen source affect secretion of pectate lyase by Colletotrichum gloeosporioides. Applied Environ. Microbiol., 69: 3258-3262.
CrossRef - Elegado, F.B. and Y. Fujio, 1994. Purification and some properties of polygalacturonase from Rhizopus sp. LKN. World J. Microbiol. Biotechnol., 10: 256-259.
CrossRef - Fawole, O.B. and S.A. Odunfa, 2003. Some factors affecting production of pectic enzymes by Aspergillus niger. Int. Biodeterior. Biodegrad., 52: 223-227.
CrossRefDirect Link - Fayyaz, A., B.A. Asbi, H.M. Ghazali, T.B. Che Man and S. Jinap, 1993. Pectinesterase extraction from papaya. Food Chem., 47: 183-185.
CrossRef - Forster, H., 1988. Pectinesterase from Phytophthora infestans. Methods Enzymol., 161: 355-357.
CrossRef - Fukoka, S., H. Kamishima, L. Sode and I. Karube, 1990. Facile isolation of endopectate lyase from Erwinia carotovaro based on electrostatic interaction. Applied Biochem. Biotecnol., 26: 239-248.
PubMed - Gado, H.M., A.Z.M. Salem, P.H. Robinson and M. Hassan, 2009. Influence of exogenous enzymes on nutrient digestibility, extent of ruminal fermentation as well as milk production and composition in dairy cows. Anim. Feed Sci. Technol., 154: 36-46.
CrossRefDirect Link - Gaffe, J., M.E. Tizando and A.K. Handa, 1997. Characterization and functional expression of a ubiquitously expressed tomato pectin methylesterase. Plant Physiol., 114: 1547-1556.
Direct Link - Gainvors, A., V. Frezier, H. Lemaresquier, C. Lequart, M. Aigle and A. Belarbi, 1994. Detection of polygalacturonase, pectin lyase and pectinesterase activities in Saccharomyces cerevisiae strain. Yeast, 10: 1311-1319.
PubMed - Galiotou-Panayotou, M., P. Rodis and M. Kapantai, 1993. Enhanced polygalacturonase production by Aspergillus niger NRRL-364 grown on supplemented citrus pectin. Lett. Applied Microbiol., 17: 145-148.
CrossRefDirect Link - Gautam, P., A. Sabu, A. Pandey, G. Szakacs and C.R. Soccol, 2002. Microbial production of extra-cellular phytase using polystyrene as inert solid support. Bioresour. Technol., 83: 229-233.
CrossRef - Ghildyal, N.P., S.V. Ramakrishna, P.N. Devi, B.K. Lonsane and H.A. Asthana, 1981. Large scale production of pectolytic enzyme by solid state fermentation. Food Sci. Technol., 18: 248-251.
Direct Link - Grigelmo-Migeul, N. and O. Martin-Belloso, 1998. Characterization of dietary fiber from orange juice extraction. Food Res. Int., 31: 355-361.
CrossRef - Gummadi, S.N. and T. Panda, 2003. Purification and biochemical properties of microbial pectinases: A review. Process Biochem., 38: 987-996.
Direct Link - Hadj-Taieb, N., M. Ayadi, S. Trigui, F. Bouabdollah and A. Gargouri, 2002. Hyper production of pectinase activities by fully constitutive mutant (CT 1) of Penicillium occitanis. Enzyme Microbial Technol., 30: 662-666.
CrossRef - Hasunuma, T., E.I. Fukusaki and A. Kobayashi, 2003. Methanol production is enhanced by expression of an Aspergillus niger pectin methylesterase in tobacco cells. J. Biotechnol., 106: 45-52.
PubMed - Holker, U., M. Hofer and J. Lenz, 2004. Biotechnological advantages of laboratory-scale solid state fermentation with fungi. Applied Microbiol. Biotechnol., 64: 175-186.
Direct Link - Hoondal, G., R. Tiwari, R. Tewari, N. Dahiya and Q. Beg, 2002. Microbial alkaline pectinases and their industrial applications: A review. Applied Microbiol. Biotechnol., 59: 409-418.
CrossRefDirect Link - Hours, R.A., C.E. Voget and R.J. Ertola, 1988. Some factors affecting pectin-ase production from apple pomace in solid-state-cultures. Biol. Wastes, 24: 147-157.
Direct Link - Huang, L.K. and R.R. Mahoney, 1999. Purification and characterization of an endo-polygalacturonase from Verticillum albo-atrum. J. Applied Microbiol., 86: 145-146.
Direct Link - Innocenzo, M.D. and F.M. Lajalo, 2001. Effect of gamma irradiation on softening changes and enzyme activities during ripening of papaya fruit. J. Food Biochem., 25: 425-438.
CrossRef - Ishii, S. and T. Yokotsuka, 1975. Purification and properties of pectin lyase from Aspergillus japonicus. Agric. Biol. Chem., 39: 313-321.
Direct Link - Jacob, N., K.N. Niladevi, G.S. Anisha and P. Prema, 2008. Hydrolysis of pectin: An enzymatic approach and its application in banana fiber processing. Microbiol. Res., 163: 538-544.
CrossRefPubMedDirect Link - Jayani, R.S., S. Saxena and R. Gupta, 2005. Microbial pectinolytic enzymes: A review. Process Biochem., 40: 2931-2944.
CrossRefDirect Link - Karam, N.E. and A. Belarbi, 1995. Detection of polygalacturonase and pectinesterases in lactic acid bacteria. World J. Microbiol. Biotechnol., 11: 559-563.
CrossRef - Kashyap, D.R., S. Chandra, A. Kaul and R. Tewari, 2000. Production, purification and characterization of pectinase from a Bacillus sp. DT7. World J. Microbiol. Biotechnol., 16: 277-282.
CrossRef - Kashyap, D.R., P.K. Vohra, S. Chopra and R. Tewari, 2001. Applications of pectinases in the commercial sector: A review. Bioresour. Technol., 77: 215-227.
CrossRefPubMedDirect Link - Kashyap, D.R., K.S. Soni and R. Tewari, 2003. Enhanced production of pectinase by Bacillus sp. DT7 using solid-state fermentation. Bioresour. Technol., 88: 251-254.
CrossRef - Kawano, C.Y., M.A.S.C. Chellegatti, S. Said and M.J.V. Fonseca, 1999. Comparative study of intracellular and extracellular pectinases produced by Penicillium frequentans. Biotechnol. Applied Biochem., 29: 133-140.
PubMed - Kobayashi, T., K. Koike, T. Yoshimatsu, N. Higaki, A. Suzumatsu and T. Ozawa, 1999. Purification and properties of a low-molecular weight, high-alkaline pectate lyase from an alkaliphilic strain of Bacillus. Biosci. Biotechnol. Biochem., 63: 72-75.
PubMed - Kotoujansky, A., 1987. Molecular genetics of pathogenesis by soft-rot Erwinia. Annu. Rev. Phytopathol., 25: 405-430.
Direct Link - Laurent, F., A. Kotoujansky and Y. Bertheau, 2000. Overproduction in Escherichia coli of the pectin methylesterase A from Erwinia chrysanthemi3937: one-step purification, biochemical characterization and production of polyclonal antibodies. Can. J. Microbiol., 46: 474-480.
PubMed - Castilho, L.R., R.A. Medronho and T.L.M. Alves, 2000. Production and extraction of pectinases obtained by solid state fermentation of agroindustrial residues with Aspergillus niger. Bioresourc. Technol., 71: 45-50.
CrossRefDirect Link - Liao, C.H., L. Revear, A. Hotchkiss and B. Savary, 1999. Genetic and biochemical characterization of an exo polygalacturonase and a pectate lyase from Yersinia enterolitica. Can. J. Microbiol., 45: 396-403.
CrossRef - Lobarzewski, J., J. Fiedurek, G. Ginalska and J. Wolski, 1985. New matrices for the purification of pectinases by affinity chromatography. Biochem. Biophys. Res. Commun., 131: 66-74.
PubMed - Lonsane, B.K., N.P. Ghildyal, S. Budiatman and S.V. Ramakrishna, 1985. Engineering aspects of solid state fermentation. Enzyme Microb. Technol., 7: 258-265.
CrossRefDirect Link - Luh, B.S. and H.J. Phaff, 1951. Studies on polygalacturonase of certain yeasts. Arch. Biochem. Biophys., 33: 212-227.
CrossRef - Maceira, F.I.G., A.D. Pietro and M.I.G. Roncero, 1997. Purification and characterization of a novel exopolygalacturonase from Fusarium oxysporum f. sp. lycopersici. FEMS Microbiol. Lett., 154: 37-43.
CrossRef - Maldonado, M.C., A.M.S. Saad and D.A.S. Callieri, 1994. Purification and characterization of pectinesterase produced by a strain of Aspergillus niger. Curr. Microbiol., 24: 193-196.
CrossRef - Maldonado, M.C. and S.A.M. de Saad, 1998. Production of pectinesterase and polygalacturonase by Aspergillus niger in submerged and solid-state systems. J. Ind. Microbiol. Biotechnol., 20: 34-38.
Direct Link - Manachini, P.L., C. Parini and M.C. Fortina, 1988. Pectic enzymes from Aureobasidium pullulans, LV10. Enzyme Microbiol Biotechnol., 10: 682-685.
CrossRef - Marcus, L., I. Barash, B. Sneh, Y. Koltin and A. Finkler, 1986. Purification and characterization of pectolytic enzymes produced by virulent and hypovirulent isolates of Rhizoctonia solani Kuhn. Physiol. Mol. Plant Pathol., 29: 325-336.
CrossRef - Martins, E.S., D. Silva, R. Da Silva and E. Gomes, 2002. Solid state production of thermostable pectinases from thermophilic Thermoascus aurantiacus. Process Biochem., 37: 949-954.
CrossRef - McCarthy, R.E., S.F. Kotarski and A.A. Salyers, 1985. Location and characteristics of enzymes involved in the breakdown of polygalacturonic acid by Bacteroides thetaiotaomicron. J. Bacteriol., 161: 493-499.
Direct Link - Micheli, F., 2001. Pectin methylesterase: cell wall enzymes with important roles in plant physiology. Trends Plant Sci., 6: 414-419.
PubMed - Mill, P.J., 1966. The pectic, enzymes of Aspergillus niger mercury activated exopolygalacturonase. Biochem. J., 99: 557-561.
Direct Link - Mill, P.J., 1966. The peptic enzymes of Aspergillus niger: A second exopolygalacturonase. Biochem. J., 99: 562-565.
Direct Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Miyazaki, Y., 1991. Purification and characterization of an endo-pectate lyase from Bacillus macerans. Agric. Biol. Chem., 55: 25-30.
Direct Link - Morgavi, D.P., K.A. Beauchemin, V.L. Nsereko, L.M. Rode and A.D. Iwaasa et al., 2000. Synergy between ruminal fibrolytic enzymes and enzymes from Trichoderma longibrachiatum. J. Dairy Sci., 83: 1310-1321.
CrossRefDirect Link - Murad, H.A. and M.M.E. Salem, 2001. Utilization of Uf-permeate for producing exopolysaccharides from lactic acid bacteria. J. Agric. Mansoura Univ., 26: 2167-2175.
Direct Link - Murad, H.A and M.S. Foda, 1992. Production of yeast polygalacturonase on dairy wastes. Bioresour. Technol., 41: 247-250.
CrossRefDirect Link - Murad, H.A., 1998. Utilization of ultrafiltration permeate for production of Beta-galactosidase from Lactobacillus bulgaricus. Milchwissenschaft, 53: 273-276.
Direct Link - Murad, H.H., M.A. Hanfy, A.M. Kholif, M.H. Abdel Gawad and H.A. Murad, 2009. Effect of cellulases supplementation to some low quality roughages on digestion and milk production by lactating goats. J. Biol. Chem. Environ. Sci., 4: 791-809.
Direct Link - Nagai, M., T. Katsuragi, T. Terashita, K. Yoshikawa and T. Sakai, 2000. Purification and characterization of an endo-polygalacturonase from Aspergillus awamori. Biosci. Biotechnol. Biochem., 64: 1729-1732.
PubMed - Naidu, G.S.N. and T. Panda, 1998. Production of pectolytic enzymes-a review. Bioprocess Eng., 19: 355-361.
CrossRefDirect Link - Naumov, G.I., E.S. Naumova, M. Aigle, I. Masneuf and A. Belarbi, 2001. Genetic reidentification of the pectinolytic yeast strain SCPP as Saccharomyces bayanus var. uvarum. Applied Microbiol. Biotechnol., 55: 108-111.
PubMed - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Nsereko, V.L., D.P. Morgavi, L.M. Rode, K.A. Beauchemin and T.A. McAllister, 2000. Effects of fungal enzyme preparations on hydrolysis and subsequent degradation of alfalfa hay fiber by mixed rumen microorganisms in vitro. Anim. Feed Sci. Technol., 88: 153-170.
CrossRefDirect Link - Ooijkaas, L.P., F. Weber, R.M. Buitelaar, J. Tramper and A. Rinzema, 2000. Defined media and inert supports: Their potential as solid-state fermentation production system. Trends Biotechnol., 18: 356-360.
CrossRef - Orpin, C.G., 1984. The role of ciliate protozoa and fungi in the rumen digestion of plant cell walls. Anim. Feed Sci. Technol., 10: 121-143.
Direct Link - Palomaki, T. and H.T. Saarilahti, 1997. Isolation and characterization of new C-terminal substitution mutation affecting secretion of polygalacturonases in Erwinia carotovora ssp. carotovora. FEBS Lett., 400: 122-126.
PubMed - Solis-Pereira, S., E. Favela-Torres, G. Viniegra-Gonzalez and M. Gutierrez-Rojas, 1993. Effects of different carbon sources on the synthesis of pectinase by Aspergillus niger in submerged and solid state fermentations. Applied Microbiol. Biotechnol., 39: 36-41.
CrossRefDirect Link - Phutela, U., V. Dhuna, S. Sandhu and B.S. Chadha, 2005. Pectinase and polygalacturonase production by a thermophilic Aspergillus fumigatus isolated from decomposting orange peels. Braz. J. Microbiol., 36: 63-69.
CrossRefDirect Link - Piccoli-Valle, R.H., F.M.L. Passos, F.J.V. Passos and D.O. Silva, 2001. Production of pectin lyase by Penicillium griseoroseum in bioreactors in the absence of inducer. Braz. J. Microbiol., 32: 135-140.
Direct Link - Pitkanen, K., R. Heikinheimo and R. Pakkanen, 1992. Purification and characterization of Erwinia chrysatnthemi B374 pectin methylesterase produced by Bacillus subtilis. Enzyme Microbial. Technol., 14: 832-836.
CrossRef - Polizeli, M.L.T.M., J.A. Jorge and H.F. Terenzi, 1991. Pectinase production by Neurospora crassa: purification and biochemical characterization of extracellular polygalacturonase activity. J. Gen. Microbiol., 137: 1815-1823.
Direct Link - Prade, R.A., D. Zohan, P. Ayoubi and A.J. Mort, 1999. Pectins, pectinases and plant-microbe interactions. Biotechnol. Genet. Eng. Rev., 16: 361-391.
PubMed - Banu, A.R., M.K. Devi, G.R. Gnanaprabhal, B.V. Pradeep and M. Palaniswamy, 2010. Production and characterization of pectinase enzyme from Penicillium chrysogenum. Indian J. Sci. Technol., 3: 377-381.
Direct Link - Reda, A.B., M.Y. Hesham, A.S. Mahmoud and Z.A. Ebtsam, 2008. Production of bacterial pectinase(s) from agro-industrial wastes under solid state fermentation conditions. J. Applied Sci. Res., 4: 1708-1721.
Direct Link - Reid, I. and M. Ricard, 2000. Pectinase in papermaking: solving retention problems in mechanical pulps bleached with hydrogen peroxide. Enzyme Microbial Technol., 26: 115-123.
CrossRef - Rexova-Benkova, L. and O. Markovic, 1976. Pectic enzymes. Adv. Carbohydr. Chem., 33: 323-385.
CrossRefDirect Link - Roboz, E., R.W. Barratt and E.L. Tatum, 1952. Breakdown of pectic substances by a new enzyme from Neurospora. J. Biol. Chem., 195: 459-471.
Direct Link - Rodrigues-Palenzuela, P., T.J. Burr and A. Collmer, 1991. Polygalacturonase is a virulence factor in Agrobacterium tumifaciens biovar 3. J. Bacteriol., 173: 6547-6552.
PubMed - Sakai, T. and M. Okushima, 1982. Purification and crystallisation of a protopectin- solubilizing enzyme from Trichosporon penicillatum. Agric. Biol. Chem., 46: 667-776.
Direct Link - Sakai, T. and Y. Ozaki, 1988. Protopectin solubilizing enzyme that does not catalyze the degradation of polygalacturonic acid. Agric. Biol. Chem., 52: 1091-1093.
Direct Link - Sakai, T., T. Sakamoto, J. Hallaert and E.J. Vandamme, 1993. Pectin, pectinase and protopectinase: Production, properties and applications. Adv. Applied Microbiol., 39: 231-294.
PubMedDirect Link - Sakamoto, T., R.A. Hours and T. Sakai, 1994. Purification, characterization and production of two pectic-transeliminases with protopectinase activity from Bacillus subtilis. Biosci. Biotechnol. Biochem., 58: 353-358.
PubMed - Patil, S.R. and A. Dayanand, 2006. Production of pectinase from deseeded sunflower head by Aspergillus niger in submerged and solid-state conditions. Bioresour. Technol., 97: 2054-2058.
CrossRef - Savary, B.J., 2001. Perfusion chromatography separation of the tomato fruit-specific pectin methylesterase from a semi purified commercial enzyme preparation. Prep. Biochem. Biotechnol., 31: 241-258.
PubMed - Selinger, L.B., C.W. Forsberg and K.J. Cheng, 1996. The rumen: A unique source of enzymes for enhancing livestock production. Anaerobe, 2: 263-284.
CrossRef - Semenova, M.V., S.G. Grishutin, A.V. Gusakov, O.N. Okunev and A.P. Sinitsyn, 2003. Isolation and properties of pectinases from the fungus Aspergillus japonicus. Biochemistry, 68: 559-569.
CrossRefDirect Link - Shevchik, V.E., J. Robert-Baudouy and N. Hugouvieux-Cotte-Pattat, 1997. Pectate lyase PelI of Erwinia chrysanthemi 3937 belongs to a new family. J. Bacteriol., 179: 7321-7330.
Direct Link - Shin, I., S. Dowmez and O. Kilic, 1983. Study on pectolytic enzyme production from some agricultural wastes by fungi. Chem. Microbiol. Technol. Lebensm, 8: 87-90.
Direct Link - Siebert, F.B. and J. Anto, 1946. Determination of polysaccharides in serum. J. Biol. Chem., 163: 511-522.
Direct Link - Silley, P., 1985. A note on the pectinolytic enzymes of Lachnospira multiparus. J. Applied Bacteriol., 58: 145-149.
CrossRef - Silva, D., E.D. Silva, R.D. Silva and E. Gomes, 2002. Pectinase production by Penicillium viridicatum RFC3 by solid state fermentation using agricultural wastes and agro industrial by-products. Braz. J. Microbiol., 33: 318-324.
Direct Link - Souza, J.V.B., T.M. Silva, M.L.S. Maia and M.F.S. Teixeira, 2003. Screening of fungal strains for pectinolytic activity: endopolygalacturonase production by Peacilomyces clavisporus 2A.UMIDA.1. Process Biochem., 4: 455-458.
CrossRef - Sunnotel, O. and P. Nigam, 2002. Pectinolytic activity of bacteria isolated from soil and two fungal strain during submerged fermentation. World J. Microbiol. Biotechnol., 18: 835-839.
Direct Link - Szajer, I. and C. Szajer, 1985. Formation and release of pectin lyase during growth of Penicillium paxilli. Biotechnol. Lett., 7: 105-108.
CrossRef - Szajer, I. and C. Szajer, 1982. Pectin lyase of Penicillium paxilli. Biotechnol. Lett., 4: 549-552.
CrossRef - Takao, M., T. Nakaniwa, K. Yoshikawa, T. Terashita and T. Sakai, 2001. Molecular cloning, DNA sequence, and expression of the gene encoding for thermostable pectate lyase of thermophilic Bacillus sp. TS 47. Biosci. Biotechnol. Biochem., 65: 322-329.
PubMed - Tierny, Y., M. Bechet, J.C. Joncquiert, H.C. Dubourguier and J.B. Guillaume, 1994. Molecular cloning and expression in Escherichia coli of genes encoding pectate lyase and pectin methylesterase activities from Bacteroides thetaiotaomicron. J. Applied Bacteriol., 76: 592-602.
PubMed - Favela-Torres, E., T. Volke-Sepulveda and G. Viniegra-Gonzalez, 2006. Production of hydrolytic depolymerising pectinases. Food. Technol. Biotechnol., 44: 221-227.
Direct Link - Vaughan, I.L.H., T. Jakubczly, J.D. McMillan, T.E. Higgins, B.A. Dave and V.M. Crampton, 1969. Some pink yeast associated with softening of olives. Applied Microbiol., 18: 771-775.
Direct Link - Ward, O.P., M. Moo-Young and K. Venkat, 1989. Enzymatic degradation of cell wall and related plant polysaccharides. Crit. Rev. Biotechnol., 8: 237-274.
CrossRefPubMedDirect Link - Warrilow, A.G.S., R.J. Turner and M.G. Jones, 1994. A novel form of pectinesterase in tomato. Phytochemistry, 35: 863-868.
PubMed - Wattad, C., S. Freeman, A. Dinoor and D. Prusky, 1995. A nonpathogenic mutant of Colletotrichum magna is deficient in extracellular secretion of pectate lyase. Mol. Plant-Microbe Interac., 8: 621-626.
Direct Link - Whitaker, J.R., 1984. Pectic substances, pectic enzymes and haze formation in fruit juices. Enzyme Microb. Technol., 6: 341-349.
CrossRefDirect Link - Wu, Y.T., M. Pereira, A. Venoncio and J. Teixeira, 2000. Recovery of endopolygalacturonase using polyethylene glycol-salt aqueous two phase extraction with polymer recycling. Bioseparation, 9: 247-254.
Direct Link - Yakoby, N., I. Kobiler, A. Dinoor and D. Prusky, 2000. pH regulation of pectate lyase secretion modulates the attack of Colletotrichum gloeosporioides in avocado fruits. Applied Environ. Microbiol., 66: 1026-1030.
Direct Link - Yang, W.Z., K.A. Beauchemin and L.M. Rode, 1999. Effects of an enzyme feed additive on extent of digestion and milk production of lactating dairy cows. J. Dairy Sci., 82: 391-403.
CrossRefPubMedDirect Link - Diaz Borras, M.A., R. Vila Aguilar and E. Hernandez Gimenez, 1987. Comparacion de la actividad pectinolitica in vitro entre cepas de penicillium sensibles y resistentes a fungicidas comerciales. Alimentaria, 24: 57-58.
Direct Link - Aguilar, G. and C. Huitron, 1990. Constitutive exo-pectinase produced by Aspergillus sp. CH-Y-1043 on different carbon source. Biotechnol. Lett., 12: 655-660.
CrossRef