
Mohamed H.M. Al-Agamy
Department of Microbiology, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 193-245







ABSTRACT
Not available
PDF Abstract XML References Citation
Received: June 29, 2010;
Accepted: October 08, 2010;
Published: November 04, 2010
How to cite this article
Mohamed H.M. Al-Agamy, 2011. Tools of Biological Warfare. Research Journal of Microbiology, 6: 193-245.
URL: https://scialert.net/abstract/?doi=jm.2011.193.245
URL: https://scialert.net/abstract/?doi=jm.2011.193.245
INTRODUCTION
Mass destruction weapons include nuclear, chemical and biological weapons (bioweapons). Direction of bioweapons against the military is Biological Warfare (BW). However, direction against civilians is bioterrorism. One of the main goals of the BW is the undermining and destruction of economic progress and stability. BW is the use of pathogens such as viruses, bacteria, other disease-causing Biological Agent (BA), or the toxins produced by them as bioweapons. There is a clear overlap between BW and chemical warfare, as the use of toxins produced by living organisms is considered under the provisions of both the biological and toxin weapons convention and the chemical weapons convention (Wheelis et al., 2006). The ideal agent for BW and bioterrorism would be capable of producing illness in a large percentage of those exposed, be disseminated easily to expose large numbers of people (e.g., through aerosol), remain stable and infectious despite environmental exposure and be available for production in adequate amounts. Fortunately, very few agents have these characteristics. The BAs used in bioweapons can often be manufactured quickly and easily. However, the delivery of BA in an effective form to a vulnerable target is difficult. For example, anthrax is considered an effective agent for several reasons. It forms hardy spores, perfect for dispersal aerosols. Pulmonary infections of anthrax usually do not cause secondary infections in other people. Weaponization of BAs has a long history of use and progressed from crude forms during ancient world and in medieval ages, when snake venom and infected cadavers were used as weapons in battles, to sophisticated preparations for use during and after the Second World War. The use of the bioweapons may be intended to kill, incapacitate, or seriously impair a person, group of people, or even an entire population. In the last six decades the BW is science and technique used mainly by military that can be used by nation-states or non-national groups (Bioterrorism). Development of bioweapons on a military significant scale was initiated in several countries in the period between the World Wars. Several countries had active programs such as US, UK, Canada, Germany, Japan and the Soviet Union (Roffey et al., 2002). There are four kinds of BA: bacteria, viruses, rickettsiae and fungi. Bioweapons are distinguished by being living organisms, that reproduce within their host victims, who then become contagious with a deadly, if weakening, multiplier effect. Toxins in contrast do not reproduce in the victim and need only the briefest of incubation periods; they kill within a few hour (Moran et al., 2008).
The classification of BAs was based on several criteria. Some of them include severity of effect on human health, potential for delivery in a weapon, capacity to cause panic and special needs for development and stockpiling of medication. BAs were classified by the Centers of Disease Control and Prevention (CDC) into three major categories (Table 1). BAs that could cause the greatest harm following deliberate use were placed in category A. However, category B includes BAs capable of producing serious harm and significant mortality but their severity are lower than category A. On the other hand, category C includes emerging pathogens that could be developed for mass dispersion in future and their potential as a major health threat (Hilleman, 2002). Category A Bas includes anthrax, plague, tularemia, botulism and smallpox.
| Table 1: | List of category A, B and C biological agents (Moran et al., 2008) |
 | |
CDC classified Viral Hemorrhagic Fevers (VHF) as category A Bas. On the other hand, category B Bas includes alphavirus (viral encephalitis such as Venezuelan Equine Encephalitis (VEE), Q fever, brucellosis, ricin toxin, Clostridium perfringens epsilon toxin (ETX) and Staphylococcal Enterotoxin B (SEB). Category C BAs includes Nipah viruses, Hantavirus, Tick-borne encephalitis, yellow fever, multi-drug resistant T.B (Lane et al., 2001). The most important bacterial causative agents of serious infections that could be misused for BW purpose are B. anthracis, F. tularensis and Y. pestis (Moran et al., 2008).
There are many tools involved in BW such as BAs (pathogens or toxins) used in BW, production of those agents as weapon (Weaponization) and delivery of the BAs. On other hand the biological defense measures (prevention and prophylaxis by vaccination and treatment) to prevent the disease cause by potential Biological Warfare Agents (BWA). The most important tool of the biological warfare will be discussed in this review. This review aims to summarize the important knowledge about BWAs (Category A, B and C). The basic information of BWA, such as microbiology of the WBA, weaponization of those agents and biological defense against BWA such as vaccine if available was discussed in this review.
HISTORICAL PERSPECTIVE OF BIOLOGICAL AGENTS AS BIOLOGICAL WEAPONS
The use of BAs in bioweapons has progressed from crude forms in early and middle ages to sophisticated preparations developed for use in BW during and after the Second World War (Bhalla and Warheit, 2004).
The ancient world: One of the earliest documented incidents of the intention to use the bioweapons is recorded in 1320 to 1000 B.C on the Philistines when plague is disseminated (Riedels, 2005). After 900 years later, the plague of Athens (430-426 BC) is credited with having played a role in Sparta's winning the Peloponnesian War (Ramirez, 1996). During the First Sacred War in Greece, in about 590 BC, Athens and the Amphictionic League poisoned the water supply of the besieged town of Kirrha with the toxic plant Hellebore. The Roman commander Manius Aquillus poisoned the wells of besieged enemy cities in about 130 BC (Christopher et al., 1997). From 249 to 270 AD, epidemic plague occurred in Egypt and throughout the entire empire (Russell, 1968). During the 4th century BC Scythian archers tipped their arrow tips with snake venom, human blood and animal feces to cause wounds to become infected. There are numerous other instances of the use of plant toxins, venoms and other poisonous substances to create biological weapons in antiquity (Mayor, 2003).
Medieval biological warfare: Tatars inadvertently brought bubonic plague from central Asia to Middle East and Europe. The Black Death (plague) killed millions of people, approximately one half of the population, throughout Europe in the 14th century (McGovern and Friedlander, 1997). Tatar army besieging the city of Kaffa used a combination of psychological warfare and BW (Kornpeter et al., 2001). The ubiquitous rat and an outbreak of the bubonic plague among the troops worked for Tatar army besieging Kaffa in 1346. Tatars catapulted bodies of plague victims over the walls of Kaffa in an attempt to initiate an epidemic upon the residents (Mayor, 1997). The incident of using plague corpses for BW occurred in 1710, when Russian forces attacked the Swedes by flinging plague-infected corpses over the city walls of Reval (Derbes, 1966).
Modern times
The 18th century: Smallpox virus was employed as a weapon during French and Indian war of 1754-1767. Jeffery Amherst is said to have written letters encouraging the use of virus contaminated blanket to inoculate the Indians. Distribution of contaminated blanket resulted in smallpox epidemic in Indians (Noah et al., 2002). The Native American population was decimated after contact with the old world due to the introduction of many different fatal diseases. Smallpox blanket incident in American history took place in the midst of Pontiac's Rebellion during a parley at Fort Pitt on June 24, 1763. Ecuyer gave representatives of the besieging Delawares two blankets and a handkerchief that had been exposed to smallpox, hoping to spread the disease to the Natives in order to end the siege (Fenn, 2000). The roots of diseases that killed millions of indigenous peoples in the Americas can be traced back to Eurasians living for millennia in close proximity with domesticated animals (Eitzen and Takafuji, 1997).
The 19th century: In 1834 Cambridge Diarist Richard Henry Dana visited San Francisco on a merchant ship. His ship traded many items including blankets with Mexicans and Russians who had established outposts on the northern side of the San Francisco Bay. The histories document that the California smallpox epidemic began at the Russian fort soon after they left. Blankets were a popular trading item and the cheapest source of them was second-hand blankets which were often contaminated. During the American Civil War, General Sherman reported that Confederate forces shot farm animals in ponds upon which the Union depended for drinking water. This would have made the water unpleasant to drink, although the actual health risks from dead bodies of humans and animals which did not die of disease are minimal (Fenn, 2000).
The 20th century: During the First World War, Germany is said to have used anthrax and glanders in an effort to destroy animals used during the war. Toward this end, inoculated horses and mules were shipped to allies. Although, some mortality in animals was reported, effects were not significant and did not pose a serious threat. Germany pursued an ambitious biological warfare program using diplomatic pouches and couriers, the German General Staff supplied small teams of saboteurs in the Russian Duchy of Finland and in the then-neutral countries of Romania, US and Argentina (Leitenberg, 2001). German intelligence officer and US citizen Dr. Anton Casimir Dilger established a secret lab in the basement of his sister's home in Chevy Chase, Maryland, that produced Glanders which was used to infect livestock in ports and inland collection points including, at least, Newport News, Norfolk, Baltimore and New York and probably St. Louis and Covington, Kentucky. In response to bioweapons development in Japan and at the time suspected in Germany, the US, UK and Canada initiated a BW development program in 1941 that resulted in the weaponization of tularemia, anthrax, brucellosis and botulism toxin. There was a period of development by many nations and Japanese Unit 731, based primarily at Pingfan in occupied China and commanded by Shiro Ishii, did research on BW, conducted forced human experiments, often fatal, on prisoners and provided bioweapons for attacks in China (Williams and Wallace, 1989). During the Sino-Japanese War (1937-1945) and the Second World War, Japan carried out a large-scale human experimentation on the prisoners at its experimentation station, called Unit 731, in PingFan, Manchuria. These experiments with B. anthracis, Y. pestis, V. cholerae, N. meningitis and other bacteria were carried out on several thousand prisoners. Although, the precise number of casualties is not known, estimates place the number of people killed from 5000 to over 10000 (Christopher et al., 1997; Leitenberg, 2001). Several incidents of Japanese aircrafts dropping grains and flees contaminated with Y. pestis over cities in China in 1940s were reported (Noah et al., 2002). In 1940, a Japanese plane is said to have scattered contaminated rice over the city of Chuhsien. This was followed by an outbreak of plague in a region where the disease was not seen before. There is also an indication that Germany had planned to drop typhus and plague infected rats over England during the Second World War (Lamb, 2001). During the 1948 Arabian-Israel War, Red Cross reports raised suspicion that the Jewish Haganah militia had released Salmonella typhi into the water supply for the city, causing an outbreak of typhoid among the inhabitants. Soviet Union is believed to have instituted a massive biological weapons program in the post the Second World War era. In 1979, an accident at a biological warfare facility in Sverdlvosk resulted in the release of anthrax. This was followed by an epidemic of anthrax and resulted in 77 cases and 66 deaths in a community downwind from a military microbiology facility; deaths in individuals who contracted inhalation anthrax occurred within 28 days (Lamb, 2001). Although the source of anthrax release was not initially acknowledged, in 1992, Soviet President Yeltsin admitted to an accident at the BW facility and outlawed activities that violated the 1972 Biological Weapons Convention (Noah et al., 2002).
The 21st century: On September 18, 2001 and for a few days after several letters were received by members of the US Congress and media outlets containing anthrax spores: the attack killed five people (Bartlett et al., 2002).
TOOLS OF BIOLOGICAL WARFARE
Bas (pathogens or toxins) and its production as weapon (Weaponization) are the most important tools to achieve BW. In addition, the delivery of the BAs to the victims, the biological defense measures to prevent the disease cause by potential BWA and application of biotechnology and genetic engineering are involved within the tools of BW.
Biological agents: BW is ability to use BAs (pathogens or toxin) as weapons. Bacteria, viruses, rickettsiae and fungi are the main kinds of BWAs. Bioweapons are distinguished by being living organisms, that reproduce within their host victims, who then become contagious with a deadly, if weakening, multiplier effect. Toxins in contrast do not reproduce in the victim and need only the briefest of incubation periods; they kill within a few h (Pohanka and Skladal, 2009). BAs that have been identified as posing the greatest threat are variola major (smallpox), B. anthracis (anthrax), Y. pestis (plague), Clostridium botulinum toxin (botulism), F. tularensis (tularaemia), filoviruses (Ebola hemorrhagic fever and Marburg hemorrhagic fever) and arenaviruses Lassa (Lassa fever) and Junin (Argentine hemorrhagic fever) (Broussard, 2001). The list of potential BAs is extensive (Table 1) and based on the risk to national security, these agents have been prioritized into three categories (Broussard, 2001). Agents classified as high-risk or category A can be easily disseminated or transmitted person-to-person, cause high mortality with potential for major public health impact, may cause public panic and social disruption and require special action for public health preparedness. The second highest priority or category B agents include those that are moderately easy to disseminate, cause moderate morbidity and low mortality and require enhanced disease surveillance. Category C includes emerging pathogens that could be engineered for mass dissemination in the future because of availability, ease of production and dissemination and potential for high morbidity and mortality and major health impact (Guarner and Zaki, 2006).
Requirements for an ideal biological warfare agent: Although, many pathogens and toxins cause disease or intoxication of humans, relatively few would actually be effective if employed as bioweapons. For a number of reasons, some that might be useful on a small scale, such as an assassination weapon or a terrorist weapon, would normally not be applicable on a large scale. The key factors that make a biological pathogen or toxin suitable for a large-scale biowarfare attack include:
Availability or ease of production: BAs are often simpler to acquire and produce than nuclear and chemical weapons. Some of BAs such as anthrax and brucellosis occur naturally in animals in certain parts of the world and individually can easily obtain these BAs. Many BAs can be produced in large quantities with modern fermentation and viral production technologies. Some toxins, like ricin, are widely available because their source in nature is ubiquitous and the process necessary to harvest the toxin is technically straightforward. On the other hand, some replicating agents are very difficult to grow in quantity or to a high enough titre and many toxins are produced in nature in such low quantities that harvesting them is impractical. Some of the rare toxins, like saxitoxin, could be used on a very limited scale to target one or a few individuals but currently could not be used as weapons of mass destruction. Some of these limitations could possibly be overcome by future adversaries, who might use recombinant techniques to amplify production of some of these compounds (Garrett, 2000).
Incapacitation and lethality: BWAs are likely to be selected for their ability to either incapacitate or kill the human targets of the attack. BWA does not necessarily have to be lethal to be useful as a military weapon. An agent such as VEE virus could render entire military units mission-ineffective by incapacitating large numbers of unit personnel. If one of an adversary’s aims is to overload our field medical care systems, an incapacitating agent such as VEE virus might be chosen preferentially to a lethal BA. VEE casualties may require hospitalization, tying up beds and medical resources needed for the care of battle-injured soldiers (Calisher, 1994). If lethality is desired, agents such as B. anthracis; the viruses that cause Ebola hemorrhagic fever, Marburg disease and Crimean-Congo hemorrhagic fever; or Y. pestis can be used. Inhalational anthrax, pneumonic plague and certain VHFs have high case-fatality rates once infection is established in nonimmune hosts. Viruses may be particularly attractive as BWAs because specific treatments are not available for many of them (WHO, 1970; Miller et al., 2002).
Appropriate particle size in aerosol: The inhalational or aerosol route of entry into the body is by far the most important to consider when planning defences against BW attacks. An ideal BWA would be of a particle size that would allow it to be carried for long distances by prevailing winds and inhaled deeply into the lungs of the unsuspecting victims. The size range of particles that meets both of these conditions is 1 to 5 μm in diameter (Fig. 1; GlobalSecurity.org).
Ease of dissemination: Dissemination of BAs can be simple, inexpensive and delivered through a variety of means. An ideal BWA would be easily disseminated in the open air by using off-the shelf devices such as industrial sprayers or other types of aerosol-producing devices. These could be mounted on an airplane, boat, car, or other moving vehicle, or even placed in a stationary position. An alternative method would be to disseminate the agent in an enclosed space (e.g., a building), where it could more efficiently infect or intoxicate humans living or working in the area. In addition to having the proper particle size, an ideal agent might also be dried (by freeze-drying or spray-drying), which would make it easier to disseminate widely and over longer distances. As noted above, some toxins, although inherently very toxic, have low enough lethality that massive (ton) amounts would be required to cover any significant area. Delivering such amounts to a battlefield might be logistically very difficult, if not impossible. Although, logistical difficulties militate against using toxins of low lethality as open-air weapons, they could be used on a smaller scale (Stern, 1999).
 | |
| Fig. 1: | Droplet size penetration of respiratory passages |
Stability after production: Once a BWA is produced in quantity, it must be fairly stable, either in bulk storage or once put into a weapon or a delivery system. It must, therefore, retain its viability and virulence or toxicity during production, storage, transportation and delivery. Agents that have a very short shelf life or those that are subject to rapid degradation in the environment once released would, by definition, be poor bioweapons. On the other hand, those with long shelf lives, or those that are very slowly degraded by environmental influences such as temperature extremes, dryness, or ultraviolet radiation, would be more useful as weapons, assuming other requirements are met. Anthrax spores are one example of an ideal agent from this standpoint, as they are both very hardy in the environment and stable in storage (Manchee et al., 1994).
Category A biological agents: Anthrax, botulism, plague, smallpox, tularaemia and viral hemorrhagic fevers are diseases caused by the Category A BWAs, so-called because they pose particularly serious threats as bioweapons. The key features of BWAs that pose particularly serious risks if used as weapons against civilian populations (Center of Disease Control and Prevention, 2004):
| • | High morbidity and mortality |
| • | Potential for person-to-person transmission |
| • | Low infective dose and highly infectious by aerosol dissemination, with a commensurate ability to cause large outbreaks |
| • | Effective vaccine unavailable or available only in limited supply |
| • | Potential to cause public and healthcare worker anxiety |
| • | Availability of pathogen or toxin |
| • | Feasibility of large-scale production |
| • | Environmental stability |
| • | Prior research and development as a biological weapon |
Bacillus anthracis: Robert Koch used B. anthracis to demonstrate pathogenesis as a pathogen-host relation in 1877. Bacillus anthracis is the causative agent of well known disease called anthrax which predominantly infecting domesticated and wild animals, especially herbivores (Baillie and Read, 2001). The name of Bacillus derives its name from the Greek word for coal, B. anthracis, because of its ability to cause black, coal-like cutaneous eschars. B. anthracis is a large Gram-positive, aerobic, spore bearing bacillus, 1-1.5x3-10 μm in size, is the only obligate pathogen within the genus bacillus. The spores are very resistant to harsh physical conditions and even long-term stability up to 200 years was observed; for this reason, virulent cadavers should be burned (Titball et al., 1991). It is convenient to class B. anthracis within the B. cereus group, which comprises B. cereus, B. anthracis, B. thuringiensis and B. mycoides by phenotype (Turnbull, 1999). Although, it is not possible to discriminate between species by 16S rRNA sequencing, multiple locus variable number tandem repeat analysis and amplified fragment length polymorphism provide clear evidence that B. anthracis can be reliably distinguished from others of the B. cereus groups (Keim et al., 1997). In addition, although B. anthracis is one of the most molecular monomorphic bacteria known, all known strains have been separated into five categories on the basis of variable numbers of tandem repeats in the variable region of the VrrA gene (Jackson et al., 1998). Unlike the other members of the B. cereus group, B. anthracis is non-motile and non-haemolytic on blood agar, grows at 37°C and forms typical white colonies with bee’s eye appearance that is, oval, slightly granular but not dry, about 2 mm in diameter. Spores do not form in host tissues unless the infected body fluids are exposed to air. When nutrients are exhausted, resistant spores form that can survive in soil for decades (Manchee et al., 1994). These spores then germinate when exposed to a nutrient rich environment, such as the tissues or blood of an animal or human host. In infected blood or tissues, the bacilli are frequently present in short chains, surrounded by the polypeptide capsule, which can be visualised under the microscope if stained with polychrome methylene blue. In stained smears from colonies cultivated on plates there is no capsule unless the medium contains 0.7% biocarbonate or 5% serum and the plates are incubated in 5-10% carbon dioxide (Carman et al., 1985) Provided no antibiotics have been prescribed, there is usually no difficulty in isolating B. anthracis from clinical specimens. However in environmental sampling because of the presence of other Bacillus sp., a selective medium is recommended, such as polymyxin lysozyme EDTA thallus acetate agar (Knisely, 1966). The major virulence factors of B. anthracis are encoded on two virulence plasmids pXO1 and pXO2 (Hanna, 1998). The tri-toxin bearing plasmid pXO1 is 184.5 kbp in size and codes for three toxins, which cause haemorrhage, oedema and necrosis (Leppla, 1991). They comprise the 83 kDa lethal factor, 89 kDa oedema factor and the 85 kDa protective antigen (O’Brien et al., 1985). The smaller capsule bearing plasmid pXO2 is 95.3 kbp in size and encodes three genes (cap B, cap C and cap A) involved in the synthesis of the poly-D-glutamyl capsule that inhibits host phagocytosis of the vegetative form of B. anthracis. Both plasmids are necessary for full virulence; loss of either results in an attenuated strain. Sterne, such as attenuated strain, carries pXO1 and therefore, can synthesise exotoxin, but does not have a capsule (Hanna, 1998).
Three forms of anthrax are known. The most common one is the cutaneous form where spores of B. anthracis pass through minor skin breaks, resulting in the formation of dermal ulcers. The second form is gastrointestinal and it occurs most commonly after ingestion of poorly cooked meat contaminated with spores. The intestinal ulcers are formed initially and later spread into the lymphatic system, finally causing septicaemia. The last form is pulmonary anthrax (woolsorter’s disease) following after breathing of 8000-40000 spores. The first symptoms are similar to influenza. After 2-3 days, high fever with haemorrhage continues, resulting from systematic infection. Gastrointestinal and inhalation anthrax are fatal without treatment and high mortality is reported even if anthrax was not diagnosed early and antibiotics were not dosed. B. anthracis is susceptible to penicillin (LaForce, 1994).
Weaponizatio n of B. anthracis: Though B. anthracis was described as BWA, attempts were also made to misuse it for terrorist attacks. B. anthracis could be considered to be the perfect agent for bioterrorism (Christopher et al., 1997). One kilogram of aerosolized B. anthracis could produce as many as 3 million deaths. The Soviet Union produced weoaponized anthrax in ton quantities during the cold war era. An outbreak of inhalation anthrax occurred near a Soviet bioweapons facility at Sverdlovsk in 1979, resulting in 77 infections and 66 deaths (Guillemin, 1999). In 2001, October, terrorists used US postal system to distribute B. anthracis lethal spores (Canter et al., 2005). Five letters were sent. The first two letters were sent to the NBC television in New York and to the New York Post on 18th September 2001. Nobody died but some people became ill. These letters were not very high-levelled considering purity of the agent; they contained 10% spores (Josefson, 2001). The one of the most virulent strains of natural B. anthracis is the Ames strain (Higgins et al., 2002). The following three letters, one sent to the Florida’s tabloid newspaper The Sun and two to Washington DC to the office of Senator Leahy and Daschle on 9th October 2001. The most shocking information was about the included particles (Bartlett et al., 2002).
The obtained B. anthracis samples were pure spores prepared as homogeneous 10 μm particles chemically stabilized. The particle size is one of the most important characteristic for bioweapons and 10 μm is considered to be optimal for penetration into lungs. Bacillus anthracis spores were detected in powder form by the fluorescence-based fibre-optic biosensor (Tims and Lim, 2004). Primary antibody was immobilized on the waveguide surface and cross-linked through the biotin-avidin (bound on antibody Fc part) connection. The secondary antibody was labelled by Cy-5. The limit of detection was evaluated as 3.2x105 spores in 1 mg of powder matrix. Spores were detected in powder form by the fluorescence-based fiber-optic biosensor (Tims and Lim, 2004). Bacillus anthracis specific PCR for chromosomal and pXO1/pXO2 plasmid sequences has been described (Beyer et al., 1999). The PCR for B. anthracis is a very specific method, making possible the species identification (Kiel et al., 2008).
BIODEFENSE AGAINST ANTHRAX
Anthrax vaccine protects against anthrax disease. The first human vaccine for anthrax was developed from non-encapsulated strains of B. anthracis in 1943 by the former USSR (Pile et al., 1998). This vaccine, which is currently in use in Russia, is a human live-attenuated strain, but it has been considered unsuitable for use in US because of safety concerns (Lane et al., 2001).The vaccine used in the US (AVA), licensed in 1970, does not contain B. anthracis cells and it does not cause anthrax. However, it is an aluminium hydroxide-precipitated cell-free filtrate (Contains PA) of attenuated, non-encapsulated B. anthracis of the Sterne strain (Dixon et al., 1999). Anthrax vaccine for livestock, based on a live non-encapsulated avirulent variant of B. anthracis, was developed (Sterne, 1939). Since then, this vaccine has served as the principal veterinary vaccine in the Western Hemisphere. The use of livestock vaccines was associated with occasional animal casualties and live vaccines were considered unsuitable for humans (Salsbery, 1926).
Anthrax vaccine is recommended for certain people 18 through 65 years of age who might be exposed to large amounts of B. anthracis bacteria on the job, including: some people handling animals or animal products some military personnel, as determined by the Department of Defence. These people should get 5 doses of vaccine (in the muscle): the first dose when risk of a potential exposure is identified and the remaining doses at 4 weeks and 6, 12 and 18 months after the first dose. Annual booster doses are recommended for ongoing protection (Center of Disease Control and Prevention, 2002a, b).
The vaccine is recommended for use in the U.S. military and for post-exposure prophylaxis in public health officials. Vaccination is also recommended for laboratory personnel involved in the production of B. anthracis cultures and those performing confirmatory testing for B. anthracis. The anthrax vaccine has been administered to members of the U.S. armed forces since 1998 and was given to victims of the 2001 anthrax attacks with an IND protocol as post-exposure prophylaxis (Inglesby et al., 2002). It is approved for individuals 18 to 65 years of age and though not officially approved for children, it is felt to be safe and efficacious based on experience with other inactivated vaccines (Inglesby et al., 2002). AVA has not been shown to have an effect on birth rates and pregnancy or result in adverse birth outcomes. Despite its use for the prevention of anthrax, several studies have underscored the relatively high incidence of toxicity attributable to AVA.
Yersinia pestis: The genus Yersinia consists of 11 species, of which 3 are human pathogens (Y. pestis, Y. pseudotuberculosis and Y. enterocolitica). The type species, Y. pestis, is a Gram-negative, nonmotile, non-sporing, slowly growing, coccobacillus. Y. psetis belongs to the family Enterobacteriaceae (formerly classified in the family Pasteurellaceae); it is the causative agent of the well known disease plague. Three cases of plague are known namely bubonic-, septicaemia-and pneumonic plague. Most of the cases of disease in humans that occur nowadays are bubonic plague, which is usually the consequence of the bite from a flea that has previously fed on a rodent infected with Y. pestis. It is this form of the disease that gives rise to the classical symptom of plague: the swelling of the local draining lymph node (or bubo), usually in the groin or armpit. Occasionally, the infection spreads beyond this focus of infection to the bloodstream. Such cases of septicaemia plague are difficult to treat and often result in the colonization of the lungs. A subsequent secondary pneumonia can result in the dissemination of bacteria by the airborne route, as a consequence of coughing. It is these airborne bacteria that pose the greatest risk to human health, because their inhalation can result in primary pneumonic plague. Pneumonic plague is notoriously difficult to treat because of the speed with which the disease develops (typically the incubation period is 1-3 days) and also because, by the time individuals are symptomatic, they are often close to death (Titball et al., 2003). Treatment as well as prophylactics of plague employs many antibiotics. Streptomycin was one of the choices but tetracyclines are commonly considered as more potent, now. Yersinia pestis is sensitive to penicillin in vitro but it was found to be ineffective against extended human disease (Crook and Tempest, 1992).
A laboratory diagnosis of plague is similar to other infections. Blood is recommended as specimen for analysis; bubo aspirates and sputum are other options. Yersinia pestis can be cultivated on routine laboratory culture media, such as sheep blood agar. MacConkey and eosin-methylene-blue agars are chosen as selective media. Cultivation at 37°C for 2 days is optimal to produce visible colonies. Some of the cultivation techniques are convenient for challenging specific marker synthesis; the recent experience of Fedorova et al. (2005) should be mentioned: they were able to induce biosynthesis of YopE (plasmid coding effector protein qualifying pathogenicity of Y. pestis). Biosynthesis was proved at cultivation in the presence of substances from phagolysosome and/or a typical condition in them (Chanteau et al., 1998) and in zoonotic vectors (Kilonzo et al., 2006) is detectable by ELISA. F1 is specific for Y. pestis; moreover, it is not only a good marker approachable for identification but also for immunization of protecting against wild variants of plague (Wang et al., 2008). Another promising antigen is ATP-binding cassette transporter proteins consisting of OppA, PstS, YrbD and PiuA (Tanabe et al., 2006). The recent effort pointed at a plausible application of antigens being used for vaccine construction as good markers for identification of Y. pestis during an immunoassay (Smither et al., 2007). PCR assays frequently employ pla and a structural gene for the F1 antigen caf1 (Rahalison et al., 2000). This gene was also found to be long-term stable and in this way suitable for an interesting study of plague victims from the 16th-18th century (Bianucci et al., 2008).
Plague has a remarkable place in history. It has played a role in both natural disasters and war and has been used as a weapon since at least medieval times (Ligon, 2006). The relationship between humans and plague has been known from ancient history. About 200 million has been suggested as a credible number for palgue death toll throughout recovered history. Three pandemics of plague were recorded. The Justinian plaque epidemic (First pandemic) spread from Egypt into Mediterranean Europe after 541 A.D (Russell, 1968). From 14th to 18th centuries, Europe seems to have escaped most epidemic disease. Second epidemic of plague (Black Death epidemic 1347-1351) influenced the European population resulting in up to 40 % casualties.
| Table 2: | Human plague cases between 1994 to 2003 in the countries that reported >100 confirmed or suspected cases (WHO, 2004) |
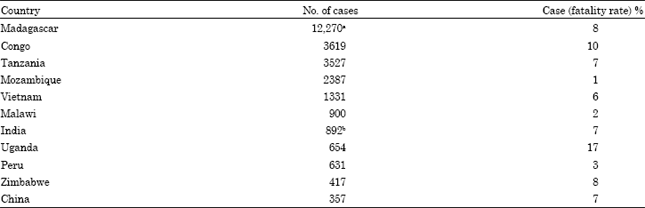 | |
| aClinical forms from Madagascar in 1996-1998 reported as 97% bubonic and 3% pneumonic cases (Chanteau et al., 2000). Other countries also reported predominantly bubonic plague. Septicemic plague was rarely reported, but in 2006, the United States reported that 5 (38%) of its 13 cases were septicemic (Centers for Disease Control and Prevention, 2006). bCases in India were predominantly pneumonic plague | |
This plaque pandemy was spread in consequence of tragic events. Probably at the beginning, Tatar forces in 1346 during the battle for the seaport city Kaffa placed in today’s Ukraine catapulted plague victims into the city in order to spread this disease (Christopher et al., 1997). The Genoese merchants who escaped from Kaffa and docked in Genoa in October 1347 probably transferred to Mediterranean ports infected rats and bubonic plague was consequently spread in Europe (Derbes, 1966). The third pandemic started in the 19th century, plague was spread into Hong Kong from China and consequently to the remaining parts of Asia, America and Africa (Perry and Fetherston, 1997). Plague caused deadly epidemics in medieval Europe and in the 20th century, it caused extensive mortality in India and Vietnam. Crossing into the 21st century, it has attracted particular attention as a potential bioweapons, for which a new vaccine needs to be developed. Considerable strides have been made in understanding the causative organism’s virulence, although plague has persisted as a killer disease in Africa, Asia and the Americas (Butler, 2008).
For the decade 1994-2003, the WHO reports that the number of confirmed and suspected human cases of plague in all countries was 28, 530, with 2015 deaths, for a case fatality rate of 7.1% (Table 2).
Weaponization of Y. pestis: Attempts to use plague as a bioweapon dated back to the ancient practice of flinging plague-infected corpses over the walls of cities under siege. The Japanese attempted to use plague as a bioweapon by releasing infected fleas over cities in Manchuria during the Second World War, but dissemination attempts met with limited success. US did not develop plague as a potential weapon because of its persistence in the environment and the possibility of noncombatant and friendly casualties after an attack. The Soviet Union reportedly developed dry, antibiotic-resistant, environmentally stable forms of Y. pestis that could be disseminated as an aerosol (Holty et al., 2006). Though Y. pestis is unstable in aerosol for longer times, which makes its use as BWA difficult; CDC enlisted it into category A due to the high mortality and high virulence. Unlike anthrax, Y. pestis does not form spores and does not survive well outside the bodies of persons or animals. For this reason, no one has succeeded in developing an effective bioweapon using aerosolized bacteria. Efforts are under way to develop new subunit vaccines that will protect persons against plague. Earlier vaccines to prevent flea-borne plague have been used for more than half a century for persons in areas of endemicity, including>1 million US military personnel deployed to Vietnam in the 1960 and 1970s, but the formalin killed whole cell plague vaccine, which did not protect persons against pneumonic disease, was discontinued by its US manufacturer in 1998. The LAV EV76 has been used for a long time in Europe and other countries but is not commercially available. New subunit vaccines that contain F1 and V antigens of Y. pestis are being tested for safety and immunogenicity. Although, this is an active area of current research, doubts have been raised about whether subunit vaccines that engender antibody responses will protect against pneumonic plague, which has an intracellular phase and may require a cell-mediated immune response for protection (Parent et al., 2005).
Biodefense against plague: Since, the 1950s, the former USSR and the US have developed methods to aerosolize Y. pestis that could be used to induce primary pneumonic plague. There is also concern that multi-drug resistant strains of Y. pestis could be used as bioweapons (Inglesby et al., 2000). In 1970, WHO estimated that, in a worst-case scenario, 50 kg of aerosolized Y. pestis released over a city of five million could result in 150, 000 cases of pneumonic plague and 36, 000 deaths (Inglesby et al., 2000). Therefore, the fear of illegitimate use of Y. pestis as a bioweapon has renewed interest in developing a vaccine to prevent the devastating diseases it causes.
Both antibiotics and vaccines have been used to prevent Y. pestis infections. The tetracyclines are the popular antibiotics for plague prophylaxis. Plague vaccines have been used since the late 19th century, but their effectiveness has never been measured precisely. There are two types of plague vaccine currently used in various parts of the world. The LAV is derived from a Pgm- (pigmentation derived mutant) strain usually related to EV76, while killed vaccine uses a formalin-fixed virulent strain of Y. pestis (Titball and Williamson, 2001). Field experience indicates that vaccination with plague vaccine reduces the incidence and severity of disease resulting from the bite of infected fleas. The degree of protection afforded against primary pneumonic infection is not known. The plague vaccine licensed for use in US is prepared from Y. pestis organisms grown in artificial media, inactivated with formaldehyde and preserved in 0.5% phenol (Center of Disease Control and Prevention, 1982). Both killed vaccine and LAV for plague have been used in humans. A formaldehyde-killed whole cell vaccine was the only licensed vaccine for plague in the US, but the manufacture of this vaccine was discontinued in 1999. There is indirect evidence that the vaccine may be effective in humans, in light of the reduced incidence of bubonic plague in vaccinated U.S. servicemen who served in Vietnam (Titball and Williamson, 2001). In contrast, there is no conclusive evidence that the killed whole cell vaccine is protective against pneumonic plague, which could develop from an aerosolized weapon (Perry and Fetherston, 1997). Currently, new generation subunit vaccines for plague, including some that utilize mucosal delivery systems, are under investigation. The importance of a safe and effective vaccine for plague is underscored by the isolation of multi-drug resistant Y. pestis, which illustrates the risk of relying upon antibiotics for post-exposure prophylaxis for agents of bioterror (Inglesby et al., 2000).
Persons exposed to plague patients who have pneumonia or to Y. pestis aerosols in the laboratory should be given a 7- to 10-day course of antimicrobial therapy regardless of vaccination history. Recommended antimicrobials include tetracyclines, chloramphenicol, or streptomycin.
Francisella tularensis: F. tularensis is the causative agent of tularaemia; it is a small, intracellular and non-motile Gram negative coccobacillus requiring aerobic conditions. Tularaemia was first notified in Californian’s Tulare Country in 1911 (McCoy and Chapin, 1912). The human disease was recognized and described by Edward Francis in 1922. Lately, the taxonomic position was changed to Pasteurella and even Brucella; the final taxonomic position was proposed in 1947 and the genus was called Francisella (Dorofe’ev, 1947). Francisella tularensis remains viable for weeks in water, soil, carcasses and hides and for years in frozen meat. There are several subspecies of F. tularensis formerly, it was divided into subtypes A and B; at present, four subspecies are described. The most virulent is the subspecies tularensis (subtype A; also named as nearctica by investigators in the former Soviet Union) occurring in North America. Recently, F. tularensis subsp. tularensis was reported in Europe (Gurycova, 1998). This subspecies is able to ferment glycerol as well as L-citrulline (Olsufjev et al., 1959). Analysis of 16S rRNA is more important for distinguishing this subspecies rather than metabolic activity studies. The case-fatality rate for this subspecies reached 1.4% in the US during 1985-1992. Subspecies holartica (subtype B; can be referred to as palaeartica) is found in North America as well as in Eurasia. This subspecies was present in nearly 5% of rodents in China. The holarctica subspecies can be divided into three biovars: the erythromycin-sensitive biovar I, erythromycin-resistant biovar II and biovar japonica. The subspecies holartica exerts a 10% lower case-fatality rate in comparison with the tularensis subspecies. The subspecies mediaasiatica is spread mainly in central Asia, being less virulent than tularensis; on the other hand, it metabolizes L-citrulline and glycerol and exhibits high genomic similarity to the tularensis subtype. Broekhuijsen et al. (2003) using the microarray technique confirmed genetic similarity between the F. tularensis subspecies mediaasiatica and tularensis and a close genetic connection between mediaasiatica and the tularensis strain Schu S4. The last subtype of F. tularensis is novicida, being isolated from water supplies in Utah and first described as a separate species. After genetic distinction, this species was proposed to be a subspecies of F. tularensis (Hollis et al., 1989). A better distinguishing of F. tularensis subspecies will be simpler after characterization of diagnostic markers obtained by proteome analysis. Tens of specific proteins were described for every subspecies. An important factor about virulence should be obtained after complete genome analysis, which is now starting (Prior et al., 2001).
Francisella tularensis causes zoonotic disease tularaemia (Rabbit or deer fly fever) predominantly localized in the Northern hemisphere. Naturally, this disease spreads among mammals, mainly rodents, rabbits and hares. It is occurred after contact with infected animals or from the bites of infected vectors such as deer flies, mosquitoes, or ticks. It can also caused by the ingestion of contaminated food and water and the inhalation of contaminated air (Hubalek et al., 1998). The most frequent disease presentations are ulceroglandular, glandular, oculo glandular, oropharyngeal, pneumonic, typhoid and septic (Pullen and Stuart, 1945). The onset of tularaemia is quite fast. Symptoms such as high fever of 38-40°C (Plourde et al., 1992), body pain and dry cough can be observed. The infection process was also intensively investigated in multiple vectors (Bandouchova et al., 2009). For disease treatment, several antibiotics are convenient. Streptomycin and gentamicin are widely recommended but tetracycline and chloramphenicol are acceptable alternatives (Enderlin et al., 1994). In history, vaccination against tularaemia was realized using the Live Attenuated Vaccine (LAV) in the former Soviet Union in the tularaemia endemic areas and in the US Army research facility (Tamilselvam and Daefler, 2008).
Several methods are suitable for the assay of F. tularensis. When cultivation tests are used, the best growth is observed in cysteine-enriched broths and blood or chocolate-supplemented agars. The characteristically opalescent colonies are formed after 1-2 day of incubation at 37°C in wet atmosphere. A wide range of immunoassays were described. Serological diagnosis of patient sera was possible by microagglutination and ELISA (Ozcurumez et al., 2004; Schmitt et al., 2005). The PCR is typically targeted to the tul4 and fopA genes encoding 17 and 43 kDa outer membrane proteins, respectively. This technique was employed, e.g., for tissue samples from infected mice (Emanuel et al., 2000) for tissue specimens from brown hares (Lepus europaeus) infected during the epizootic period of tularaemia in Austria in 1997 (Grunow et al., 2000) and for confirmation after microagglutination tests when the infection broke out in Turkey in February 2004 (Celebi et al., 2006). DNA extraction from soil spiked with F. tularensis SHU-4 was optimized for PCR assay purposes; the lowest limit of detection reached only 20 cfu g-1 of soil (Whitehouse and Hottel, 2006). Sellek et al. (2008) performed real-time PCR based on SYBR Green I and tul4 gene for F. tularensis LVS assay.
Weaponization of F. tularensis: F. tularensis was weaponized by US in the 1950 and 1960s during the offensive biowarfare program and other countries are also suspected to have weaponized the organism. Francisella tularensis could potentially be stabilized for weaponization and produced in either a wet or dried form for delivery in a terrorist attack (Dennis et al., 2001). As few as 10 to 50 organisms may cause disease if inhaled or injected intradermally; however, approximately 108 organisms are required to cause infection after oral ingestion (Moran et al., 2008). Due to the low infection dose, 10-50 organisms in aerosol (Dennis et al., 2001) and simple cultivation F. tularensis was enrolled by CDC into category A. During the Second World War, the Japanese bioweapons operation Unit 731 expressed interest in tularaemia. Ken Alibek was secretly immigrated to US in 1992, revealed information about Soviet experimentation with tularaemia as a BWA prior to the Second World War. In his 1999 book Biohazard, Alibek reported the possible use of tularaemia against German troops shortly before the Battle of Stalingrad (Alibek and Handelman, 1999). Alibek based his claims on evidence of highly infectious tularaemia outbreaks among the soldiers during that time. Some scientists argued that an outbreak from natural causes was the more likely explanation. The former deputy director of the Soviet biological weapons program, Biopreparat, Alibek himself may have been accidentally infected by tularaemia bacteria due to a leaky fermentor. In 1982, it was reported that Biopreparat developed a vaccine-resistant strain of tularaemia. Fort Detrick in Maryland, the centre of the US biological weapons program, was built in 1942. In the 1950 and 1960s, US biological weapons program also developed the potential of using F. tularensis bacteria as a biological agent. Volunteers were infected with the bacteria through direct an aerosol delivery system and in the aerosol chamber known at Fort Detrick as the 8 ball, the largest aerosol testing facility in the world. Most of the volunteers were Seventh Day Adventists, army recruits whose religious beliefs did not allow them to participate in combat. The program developed a vaccine that partially protected against the virulent Schu S-4 strain of the tularaemia bacteria. The Schu S-4 strain was labelled Agent UL and standardized for use in E120 bursting spherical bomblet. By the 1960s, the US program was able to freeze droplets of tularaemia bacteria in a concentrated liquid culture that could be stored up to 3 years at -18°C. In 1965, near Hawaii, the US biological weapons program tested aerosolized tularaemia bacteria on barges populated with monkeys in the deck and in the hold. A military aircraft sprayed a 32 mile line of Agent UL. Over the Pacific, the bacteria were infectious for 60 miles. After a few days of incubation, half the monkeys developed symptoms and most of the sick monkeys died. Since, the end of the US bioweapons program, the US Army Research Institute continued to conduct defensive medical research on the F. tularensis agent. WHO expert committee in 1969 estimated that 50 kg of F. tularensis sprayed in aerosol form over a city of 5 million inhabitants would result in 250,000 incapacitating casualties and 19,000 fatalities. Such an aerosol attack would result in acute symptoms of pleuropneumonic tularaemia 3-5 days following the exposure (Pappas et al., 2006). Proper laboratory isolation of the F. tularensis and diagnosis could take weeks. The victims would be incapacitated for a few weeks and relapses would be expected months after the attack. The CDC estimated that a tularaemia attack would cost society $5.4 million in damages for every 100,000 persons exposed. Vaccinated individuals would only be partially protected. Inhalation tularaemia would be less virulent than inhalation anthrax or plague (Moran et al., 2008).
Biodefense against tularaemia: F. tularensis is sometimes considered a lethal BWA, since high-dose aerosol dissemination would result in a disproportionate number of cases of the pneumonic form of tularemia. Based on its potential for aerosol transmission and very low infective dose, F. tularensis is classified as a Category A bioweapon. The first LAV for tularaemia was developed from the attenuated strain Moscow in the former USSR and given to humans in 1942. The efficacy of this vaccine was reportedly demonstrated in several thousand individuals, but the strain was lost. Another live-attenuated strain (strain 15) was subsequently developed and an estimated 60 million individuals received this vaccine in the former USSR until 1960 (Sjostedt et al., 1996). In 1956, a version of strain 15 and another strain (strain 155) developed in the USSR were brought to the United States. A Live Vaccine Strain (LVS) derived from these strains protected mice and guinea pigs against inhalation of virulent F. tularensis (Cohen and Stockard, 1967) and was subsequently found to protect against inhalation of virulent F. tularensis in a small cohort of human volunteers (Saslaw et al., 1961). LVS had greater efficacy after oral and respiratory than intradermal administration (Hornick and Eigelsbach, 1966). In the 1960s, the FDA approved the LVS vaccine for use in clinical trials as an IND based on evidence it induced immunity to tularaemia. However, questions regarding its attenuation, virulence, efficacy in animal models and mechanism of efficacy have made formal licensing difficult. Importantly, the LVS vaccine is not fully protective against pneumonic tularaemia, making it unsuitable for post exposure prophylaxis.
Antibiotics (Doxycycline, 100 mg orally twice daily; Ciprofloxacin, 500 mg orally twice daily (Dennis et al., 2001) can be used for post-exposure prophylaxis in the event of mass exposure to F. tularensis, but drug-resistant strains could render antibiotics ineffective. At present, the LVS vaccine is the only effective vaccine for tularaemia. It is not currently available in US, but efforts to license it are under way. Regarding new vaccine development, there has been substantial progress in identifying LVS antigens that induce protective immune responses. Evidence that the LPS from LVS was immunogenic and protective in mice suggests that antibody could mediate vaccine efficacy and that LPS may be a suitable subunit vaccine antigen (Ellis et al., 2002). The LPS and other defined microbial determinants that can elicit surrogates for vaccine efficacy, such as specific antibody, hold promise as antigens for new vaccine development.
Clostridium botulinum toxin (BoNT): Clostridium botulinum is Gram-positive, anaerobic spore-forming bacteria and causes botulism. Botulism is a potentially fatal neuroparalytic disease, is caused by highly potent neurotoxins. C. botulinum can produce up to seven antigenically distinct exotoxins (BoNT), which are differentiated serologically by specific neutralization with antitoxin (Hatheway, 1992). BoNT have been designated as serotypes A, B, C1, D, E, F and G. Four of the seven neurotoxins (A, B, E and F) cause naturally occurring human botulism, but all seven are known to cause inhalational botulism in primates (Middlebrook and Franz, 1997). Types C and D cause botulism in primates when ingested and type C toxin was shown to be effective when used in human clinical trials treating patients for blepharospasm and cervical dystonia. Thus, it is reasonable to assume that all seven toxins can cause botulism in humans if the exposure level is high enough (Eleopra et al., 2002).
Portals of entry for BoNT include the pulmonary tract (inhalation botulism), the gastrointestinal tract (food-borne and infant botulism) and mucus membranes of wounds (wound botulism). Regardless of exposure route, toxin can enter the circulatory system from different peripheral sites and then be transported to the neuromuscular junctions of motor neurons where its destructive action occurs. The clinical hallmark of botulism is an acute flaccid paralysis, which begins with bilateral cranial nerve impairment involving muscles of the eyes, face, head and pharynx and then descends symmetrically to involve muscles of the thorax and extremities. Signs and symptoms characterizing the onset of the disease include blurred vision, ambiopia, ptosis and photophobia, followed by signs of bulbar nerve dysfunction such as dysarthria, dysphonia and dysphagia. However, early symptoms may be non-specific and difficult to associate with botulinum intoxication. If botulism is a consequence of ingesting improperly preserved foodstuffs contaminated with bacteria and their pre-formed toxins, nausea, abdominal pain, vomiting and diarrhoea may often precede or accompany the neurologic indications. As the disease progresses, death may result from respiratory failure caused by paralysis of the tongue or muscles of the pharynx causing upper airway occlusion or from paralysis of the diaphragm and intercostal muscles (Dembek et al., 2007). The rapidity of the onset of symptoms, severity and duration of the disease are dependent upon the specific serotype associated with the exposure and the level of the intoxication (Foran et al., 2003). Botulinal toxins are among the most potent toxins in existence with LD50 of roughly 0.005-0.05 μg kg-1. A single gram of crystalline toxin, evenly dispersed and inhaled, would kill more than 1 million people, although technical factors would make such dissemination difficult (Kukreja and Singh, 2009).
Weaponization of BoNT: Botulism is a rare, naturally occurring disease that can also be caused by accidental or intentional exposure to BoNT. Development and use of BoNT as a possible bioweapon began at least 60 years ago (Smart, 1997). The head of the Japanese BW group (Unit 731) admitted to feeding cultures of C. botulinum to prisoners with lethal effect during that country's occupation of Manchuria, which began in the 1930s. The US biological weapons program first produced BoNT during the Second World War, because of concerns that Germany had weaponized BoNT, more than 1 million doses of botulinum toxoid vaccine were made for Allied troops preparing to invade Normandy on D-Day (Bryden, 1989). The US biological weapons program was ended in 1969-1970 by executive orders of Richard M. Nixon, then president. Research pertaining to BW use of BoNT took place in other countries as well (Holzer, 1962).
Although, the 1972 Biological and Toxin Weapons Convention prohibited offensive research and production of bioweapons, signatories Iraq and the Soviet Union subsequently produced BoNT for use as a weapon (Bozheyeva et al., 1999). Four of the countries (Iran, Iraq, North Korea and Syria) listed by the US government have developed, or are believed to be developing, BoNT as a weapon (Bermudez, 2001). After the 1991 Persian Gulf War, Iraq admitted to the UN inspection team to having produced 19000 L of concentrated BoNT, of which approximately 10000 Litre were loaded into military weapons. These 19000 L of concentrated BoNT are not fully accounted for and constitute approximately 3 times the amount needed to kill the entire current human population by inhalation. In 1990, Iraq deployed specially designed missiles with a 600 km range; 13 of these were filled with BoNT, 10 with aflatoxin and 2 with anthrax spores. Iraq also deployed special 180 kg bombs for immediate use; 100 bombs contained BoNT, 50 contained anthrax spores and 7 contained aflatoxin. It is noteworthy that Iraq chose to weaponize more BoNT than any other of its known BA.
Despite efforts to produce an effective BoNT weapon, botulism is unlikely to ever be effectively deployed as a weapon of mass destruction. Aerosol delivery would require large quantities of toxin at the optimal time, because BoNT quickly degrades in the environment and is rendered non-lethal within minutes after release. Municipal water reservoirs are most likely safe from contamination by terrorists, because ton quantities of toxin would be necessary due to the effects of dilution. BoNT is not stable for extended periods in water and chlorination provides an effective means of destruction (Moran et al., 2008). An aerosolized or food-borne BoNT weapon would cause acute symmetric, descending flaccid paralysis with prominent bulbar palsies such as diplopia, dysarthria, dysphonia and dysphagia that would typically present 12 to 72 h after exposure (Arnon et al., 2001).
Biodefense against botulism: Vaccination is an effective strategy of providing specific protection against exotoxins such as BoNT by eliciting neutralizing antibodies that would prevent the binding of the toxin to an appropriate receptor and promote clearance and degradation by phagocytes. Early development of diphtheria and tetanus toxoid vaccines was done by Ehrlich. Efforts to develop vaccines against BoNT intensified during the Second World War because of concerns the toxin might be used as a bioweapon against allied forces and methods for preparing alum-precipitated type A and B toxoids for use in animals and humans were developed in US (Hottle et al., 1947).
Natural protection against botulism does not involve the generation of antitoxin antibodies, most likely because the action of toxins is too rapid and the toxin is present in too small an amount to elicit an antibody response (Casadevall and Pirofski, 2003). However, specific antibody can neutralize BoNT, making vaccination a reasonable approach for prevention of botulism. A licensed vaccine for botulism is not available, but pentavalent (ABCDE) botulinum toxoid (PBT) is available in the US as an IND (Cohen and Marshall, 2001). PBT has been used for more than 30 years in over 3000 laboratory workers in various countries. In the US, the CDC has distributed it for laboratory workers at high risk for botulism and the military has used it to protect troops against a potential bioterrorism attack. Despite its efficacy in preventing sporadic disease, PBT has shortfalls that limit its potential utility in a bioterror attack. For example, it does not include all serotypes, requires multiple administrations and a prolonged period of time to develop protective titres, making it unsuitable for post-exposure prophylaxis. Furthermore, it is highly reactogenic and expensive. The latter is partly related to relatively low yields of BoNT and the need for a distinct facility to safely handle and manufacture large quantities of such a potent and dangerous toxin. The reactogenicity is related to the use of thimerosal (0.01%) and formaldehyde for inactivation of toxin and prevention of reactivation (Smith, 2006). A partially purified monovalent toxoid vaccine against BoNT type F has also been evaluated in phase I and phase II trials and is available as an IND (Smith, 2006). Efforts to reduce the labor and risk involved in toxoid production have led to the development of recombinant vaccines. Recombinant vaccines use defined, reproducible and predictable antigens. Initially, genetically engineered yeast were used to produce botulinum toxins and a candidate vaccine with serotypes A, B, C and F has been produced (Smith, 2006).
Smallpox: Smallpox is an infectious disease unique to humans, caused by either of two virus variants, Variola major and Variola minor. Smallpox (variola) is a DNA orthopoxvirus that has been a scourge to humans throughout recorded history. No nonhuman reservoirs or human carriers exist for smallpox; the disease survives through continual person to person transmission. Smallpox localizes in small blood vessels of the skin and in the mouth and throat. In the skin, this results in a characteristic maculopapular rash and later, raised fluid-filled blisters. V. major produces a more serious disease and has an overall mortality rate of 30-35%. Transmission of smallpox occurs through inhalation of airborne variola virus, usually droplets expressed from the oral, nasal, or pharyngeal mucosa of an infected person. It is transmitted from one person to another primarily through prolonged face-to-face contact with an infected person, usually within a distance of 6 feet, but can also be spread through direct contact with infected bodily fluids or contaminated objects (fomites) such as bedding or clothing (Ryan and Ray, 2004).
The first documented epidemic of smallpox was during the Egyptian-Hittite war in 1350 BC. The mummy of Ramses V has lesions that suggest he died of smallpox at the age of 35 years in 1143 BC. Smallpox was used inadvertently as a bioweapon when Cortez introduced it to the new world in 1520, devastating much of the native population. The English used smallpox intentionally during the French and Indian war in 1754 when tainted blankets were distributed to Native Americans, with up to 50% mortality in many tribes. The last case of wild smallpox occurred in Somalia in 1977, although a few small outbreaks have occurred related to laboratory exposure. The disease was declared eradicated by WHO in 1980 and routine vaccination was stopped soon after (Moran et al., 2008).
Weaponization of smallpox: Smallpox was first used as a biological weapon during the French and Indian Wars (Henderson et al., 1999). Based on its devastating morbidity and fatality rate and that it can be transmitted from person to person by an aerosol route and the assumption that the US public is highly susceptible since vaccination was discontinued over three decades ago, smallpox has been classified as a Category A bioweapon. Reports that remaining smallpox stocks could be used as agents of bioterror and that they might have been weaponized (Henderson et al., 1999) have led to interest in the reintroduction of smallpox vaccine and vaccination. Because vaccination is no longer given, most persons today are susceptible to infection. Even those who were vaccinated as children are likely to be susceptible, because immunity wanes over time. Stocks of variola virus are supposedly stored at only two WHO-approved storage facilities: the CDC in Atlanta and the NPO (Scientific and Production Association) in the Novosibirsk region of Russia. The Soviet Union may have developed stockpiles of weaponized smallpox and experimented with genetic manipulation of the virus (Alibek and Handelman, 1999). Many believe that some virus samples may be in the hands of potential terrorists. Because the virus is difficult to obtain, an intentional smallpox exposure would require extensive resources that might be out of reach for small groups (Moran et al., 2008). If a biological attack is not known to have occurred, some early smallpox cases are likely to be mistaken for chickenpox or other diseases. Chickenpox differs from smallpox in that the prodrome is milder, the vesicles are superficial (i.e., easily collapse on puncture) and predominate on the trunk as opposed to the distal extremities and active and healing lesions occur simultaneously. Mortality is reported as approximately 30% overall among unvaccinated persons, but this reflects historical data in populations without modern medical care. Mortality is higher in infants and elderly individuals and would likely be much lower among healthy adults and older children. Death occurs late in the first week or during the second week of the illness and is caused by the toxemia induced by the overwhelming viremia. A rare hemorrhagic form occurs with extensive bleeding into the skin and gastrointestinal tract followed almost universally by death within a few days (Moran et al., 2008).
Biodefense against smallpox: The original smallpox vaccine was an inoculum of cowpox used by Edward Jenner in the late 18th century, which was subsequently supplanted by vaccinia virus (Bartlett et al., 2003). The current smallpox vaccine (Dryvax; Wyeth Laboratories, Madison, NJ) is a lyophilized preparation of live un-attenuated vaccinia virus that has been available in the US since the 1970s. Despite the fact that it is derived from another viral agent, the vaccinia vaccine is protective against smallpox, presumably because of antigenic mimicry in which a cross-reactive antigen induces protection (Casadevall and Pirofski, 2003). Vaccination causes a localized pustule at the site of inoculation (a Jennerian pustule), which is indicative of successful primary vaccination (Breman and Henderson, 2002). Primary vaccination confers protection in over 95% of persons for approximately 5 to 10 years (Bartlett et al., 2003). Although, immunization practices varied, primary vaccination was generally performed in the first 2 years of life and revaccination was recommended when children began and left primary school. In 1964, the WHO Expert Committee on Smallpox recommended that people in endemic areas receive primary vaccination as neonates, a booster at 12 months, followed by another every 3 years (5 to 10 years for those in nonendemic areas) (Fenner et al., 1988). The smallpox vaccine is effective and licensed for use in children. Similar to other live virus vaccines, it should not be administered to pregnant or breast-feeding women. The vaccine has not been routinely used in the US since 1972, but this policy was reassessed in the wake of increasing concerns about bioterrorism. Currently, the US has a big enough stockpile of smallpox vaccine to vaccinate everyone in the US in the event of a smallpox emergency. In December 2002, US President announced the initiation of a smallpox vaccination program (The White House Web site answers frequently asked questions about Smallpox vaccination:
http://www.whitehouse.gov/news/releases/2002/12/20021213-3.html). Members of the U.S. Armed Forces were given the vaccine and a program to vaccinate medical and health care personnel was begun in January 2003 (Bartlett et al., 2003). Although, there are only 15 million doses of smallpox vaccine available at this time, recent studies by the National Institute of Allergy and Infectious Diseases Vaccine Unit demonstrated that it can be diluted 5 to 10 times and still result in local viral replication and vesicle formation in over 97% of recipients (Breman and Henderson, 2002). The vaccine can also be used for post-exposure prophylaxis and may prevent or attenuate illness if given in the first few days after a smallpox exposure (Henderson et al., 1999).
Category B potential biological agents: Potential category B BAs is listed in Table 3. Category B is an enormous group of pathogenic microorganisms and toxins, with varying clinical characteristics, ranging from difficult to develop as weapons (e.g., viruses) with significant mortality to easily to develop as weapons (e.g., bacteria) that induce clinical syndromes with minimal mortality, to unclearly studied pathogens with unproved risk as weapons.
Further stratification of this category is warranted, one that may bring certain agents as ricin, Shigella and West Nile virus closer to category A agents, while simultaneously administering category C status to agents as Kyasanur Forest virus and epsilon toxin of Clostridium perfringens (ETX). Further stratification of the existing classifications will result in more appropriate definitions of priorities.
| Table 3: | List of category B biological warfare agents (Moran et al., 2008) |
 | |
Bacterial biological warfare agents: There are many bacterial pathogens included in category B BAs. In this review, Brucella species (Brucellosis), Chlamydia psittaci (causing parrot fever), Burholedria mallei (causing glanders), Rickettsia prowazekii (Epidemic typhus), Coxiella burnetii (Q fever) and bacterial toxin of Clostridium perfringenes.
Brucella: Brucella is slow growing Gram-negative coccobacilli, small (0.5 to 0.7 by 0.6 to 1.5 μm), non-motile, non-encapsulated and facultative intracellular parasites. Brucella species can survive for many weeks in soil and water. Brucella is the causative agent of brucellosis, which is a zoonosis. Brucellosis transmitted by ingesting infected food, consumption of un-pasteurized milk products, direct contact with an infected animal (e.g., exposure to cattle, sheep, pigs), or inhalation of aerosols. Transmission from human to human is exceedingly rare, but possible (Memish and Balkhy, 2004). Brucellosis is considered the commonest zoonosis worldwide, with an increasing number of new disease loci emerging in recent years in various development countries and associate of the developed world with the disease as a result of travel-related infections and through international food (Glynn and Lynn, 2008). Minimum infectious exposure is between 10-100 organisms.
There are a few different species of Brucella, each with slightly different host specificity. B. melitensis which infects goats and sheep, B. abortus which infects cattle, B. suis infects pigs and B. ovis infects sheep. Most disease is called by B. melitensis, whereas other species, such as B. suis, B. abortus and B. canis, are considered less virulent. The pathogenesis of the disease is unique, as exhibited by the complex immune response elicited by Brucella and the fact that the bacteria practically hide inside the macrophages, in unique compartments of acidic environment, replicating without affecting cellular viability (Gorvel and Moreno, 2002). Brucellosis in humans causes a febrile disease with nonspecific manifestations. Spondylitis, central nervous system involvement of various presentations and endocarditis are the most troublesome. Diagnosis requires isolation of the organism from blood or bone marrow specimens, but the reported sensitivity varies significantly in the literature, ranging from 10% to more than 70%. ELISA and PCR assays have been used for diagnosis (Navarro et al., 2004) and real-time PCR assays allow for rapid diagnosis (Queipo-Ortuno et al., 2005). The traditional treatment options include a combination of doxycycline and rifampin for 6 weeks, or doxycycline for 6 weeks and streptomycin for 2 to 3 weeks. Alternative regimens use other aminoglycosides, co-trimoxazole and quinolones. Relapse rates with accepted regimens are at the level of 10% and relapse usually presents in the first year post-treatment (Pappas et al., 2005a, b).
Weaponization of Brucella: Brucella was one of the first agents to be used in the development of bioweapons, in particular B. suis, which had been weaponized in the shape of particle-filled bombs, allegedly by US and possibly by other countries more than 50 years ago (Christopher et al., 2005). The attractiveness of the pathogen lies in its exquisite ability to be transmitted as an inhalational pathogen; laboratory-acquired brucellosis is a common event in non-specialized laboratories dealing with the agent. Brucella is highly infectious when aerosolized; consequently, inhalation will be the most likely rout of infection during a terrorist attack. The organism could be spread as slurry in bomblets or as a dry aerosol (Franz et al., 1997). Reports on attack rates vary and the mortality of the disease is minimal. The relatively protracted incubation period (varying from 10 days to 3 months) further compromises its potential as a bioweapon to disrupt social structures. Existence of adequate antibiotic options further compromises this pathogen’s potential, although development of resistant strains could not be excluded. Moreover, in a situation of deliberate exposure, the use of prophylactic antibiotic administration has not been clarified and universally acceptable human vaccines are not at present available (Bossi et al., 2004). A final important implication of brucellosis if used as a bioweapon is the effect on animal population of the targeted area, which might be significant for the overall economy of the region, if largely based on animal husbandry. Yet minimal mortality, availability of treatment options, protracted inoculation period and the emergence of new, more virulent potential weapons means that its inclusion among agents of bioterrorism is nowadays mainly of historical significance (Pappas et al., 2006).
Biodefense against brucellosis: Veterinary vaccines that have significant efficacy against brucellosis have been studied and employed. The vaccination of livestock to reduce enzootic disease load, in combination with the slaughter of infected animals, is largely responsible for the declining incidence of human brucellosis. In the United States, the decline of human brucellosis cases reported to the CDC has paralleled the control of infections due to B. abortus in cattle (Young, 1995). Most veterinary vaccines in use today derive from B. abortus strain 19, an attenuated organism with stable virulence, or from Rev 1, a live, virulence-stable B. melitensis strain. A review of the role of brucellosis vaccination in veterinary medicine has been published (Nicoletti, 1989).
No licensed human vaccine against brucellosis is available in most of the Western world, including US, although live Brucella vaccines have been employed at various times in many developing nations (Roux, 1989). Nonetheless, administration of either live preparation to humans is hampered by a modest but notable incidence of clinical brucellosis cases, as well as by significant hypersensitivity reactions. Such problems were noted in the former Soviet Union, where human vaccination is still widely employed and in a US trial of strain 19 and Rev 1 vaccines conducted 35 years ago (Spink et al., 1962).
Chlamydia psittaci: Chlamydial organisms belong to one of the most ubiquitous and diverse groups of bacterial pathogens, causing a broad spectrum of disease in a wide host range (In man C. trachomatis is the leading cause of preventable blindness (trachoma) world-wide and is the most common cause of sexually transmitted disease. Chlamydia pneumoniae causes acute respiratory disease and has also been associated with chronic obstructive pulmonary disease, asthma, reactive airway disease, Reiter's syndrome, sarcoidosis and atherosclerosis. The principal animal pathogens are C. psittaci (formerly called Chlamydophila psittaci), C. abortus, C. pecorum, C. suis and C. felis. C. psittaci infects a wide variety of both wild and domesticated birds, where the disease is known as psittacosis and game, pigeons and poultry, where the disease is known as ornithosis.
Chlamydia psittaci is a lethal intracellular bacterial species that causes endemic avian chlamydiosis, epizootic outbreaks in mammals and respiratory psittacosis in humans. Chlamydia psittaci is transmitted by inhalation, contact or ingestion among birds and to mammals. Psittacosis is occupational disease transmitted from birds to human. Psittacosis (psittacosis is derived from the Greek word for parrot, psittakos) in birds and in humans often starts with flu-like symptoms and becomes a life-threatening pneumonia. The disease is transmitted through the inhalational route, after exposure to contaminated bird droppings or dust generated in environments contaminated by infected bird droppings. C. psittaci is an environmentally stable pathogen. Incubation period is 1 to 2 weeks and the disease produced ranges in severity from asymptomatic to severe pneumonia. Case fatality rate is 15 to 20% in untreated patients, but 1% post-treatment. Prominent headache is reported as characteristic in the clinical presentation. During convalescence, cases of thrombophlebitis and subsequent pulmonary embolism have been reported. Diagnosis is based on serology (complement-fixing antibody). PCR assays have been developed, whereas culture is laborious and hazardous. Treatment with doxycycline results in prompt response (Gregory and Schaffner, 1997).
Weaponization of Chlamydia psittaci: Psittacosis is included in the CDC list. Undoubtedly, as an inhalational disease, it is far less potent than other pathogens of this group in creating major public health consequences. Moreover, psittacosis is a largely forgotten disease, which implies that knowledge of its pathogenetic properties is limited (a fact that may prove problematic both for weaponization and improvement of response policies). The one intriguing characteristic of the disease in terms of bioterrorism is the ability to be transferred wide distances through infected free-ranging birds. Limited data exist, however, about the ecology of the disease outside captive settings (Pappas et al., 2006).
Burkholderia mallei: Burkholderia mallei (previously known as Pseudomonas mallei) is a non-motile, Gram-negative bacillus. B. mallei is the causative agent of glanders. Glanders is a disease of horses, mules and donkeys. The infection can also occur in humans and other animals. Human infection is rare but can be severe. The route of naturally occurring infection is unclear, but infection is believed to occur through broken skin or nasal mucosa contaminated with infected material. Infection also seems to occur through an aerosol route, as evidenced by infections in laboratory workers from routine handling of cultures (Centers for Disease Control and Prevention, 2000).
Weponization of B. mallei: Its ability to cause serious illness and infect through aerosol indicate that B. mallei may have potential use in bioterrorism. In fact, this organism has been used as a bioweapon; animals were deliberately infected with glanders during the First World War (Mobley, 1995).
Melioidosis is a human illness caused by B. pseudomallei, which is clinically similar to glanders but does not seem to be particularly infectious through aerosol (Moran et al., 2008).
Coxiella burnetii: Coxiella burnetii is a Gram-negative coccobacillus, usually described in parallel with Rickettsiae, although their phylogenetic differences have been largely outlined in recent years (Maurin and Raoult, 1999). The disease is a global zoonosis that often runs undetected because of its low mortality. It is an intracellular pathogen, which after infection can survive for prolonged periods in the autophagosomes, leading to chronic infections (Raoult et al., 2005). It is usually transmitted by inhalation and the inoculum needed to induce human infection is extremely low, often mentioned as even a sole organism. The disease produced, Q fever, is largely an occupational disease in people in close contact with animal hosts of the pathogen, such as sheep. Coxiella burnetii exhibits a remarkable environmental stability and generation of aerosols (dust-borne disease) from previously infected areas does not require current presence or contact with infected animal hosts. Person-to-person transmission does not occur. The clinical syndrome induced 10 to 21 days after exposure is usually mild, in the form of a flu-like illness or an atypical pneumonia, with varying radiologic appearances. Mild elevations of serum aminotransferases, indicating hepatitis, are also common. Serious complications are rare and mortality is minimal, but the disease exhibits a tendency for chronicity, which can manifest in various forms, the most important of which is chronic endocarditis (Fenollar et al., 2001). A chronic fatigue-like syndrome has also been recognized. The diagnosis is usually based on serologic tests, such as ELISA and indirect immunofluorescence assays. PCR detection has gradually become the diagnostic gold standard (Scola, 2002), although its use is still limited to reference laboratories. Q fever is often asymptomatic and self-limited. In detected clinical cases, a short course of doxycycline (7-10 days) is the choice regimen, although alternatives, such as quinolones, erythromycin and in special populations co-trimoxazole and rifampin, exist (Choi, 2002). Chronic endocarditis requires protracted administration of doxycycline in combination with hydroxychloroquine, although valve replacement may not be avoided (Marrie and Raoult, 2002).
Weaponization of Coxiella burnetti: C. burnetii is classified as a Category B bioweapon and was one of the 10 agents in the U.S. biological weapons arsenal destroyed by 1970 after the renunciation of offensive BW. It is resistant to heat and desiccation and is infectious by the aerosol route (Cieslak et al., 2000). The historical military significance of Q fever is characteristically outlined in the large number of outbreaks in military personnel stationed in Mediterranean countries during World War II (Kelly et al., 2002), although none of these outbreaks was attributed to deliberate release. Q fever exhibits most characteristics of a potential biological weapon, excluding mortality. It is extremely stable in the environment, easily found and aerosolized in large quantities and has further significance for the animal population and the economy and subsequent living in a targeted area. Various countries have experimented with weaponizing the pathogen in the past, including the United States (Kagawa et al., 2003). In a recent attack scenario targeting a 100,000 person city, massive disruption of societal and health structure was predicted despite an extremely low mortality rate, even without further interventions (Pappas et al., 2005a, b).
Biodefense against Q fever: There are currently three vaccines for Q fever in use or under evaluation (Bratu and Lutwick, 2002), all of which are prepared from phase I organisms. A formalin-inactivated whole-cell preparation (purified Henzerling strain) is available in the US as an IND. A similar vaccine licensed in Australia (Q-Vax, Commonwealth Serum Laboratories) was found to prevent Q fever and induce long lasting immunity in humans exposed to high concentrations of aerosolized C. burnetii (Marmion et al., 1984). The whole cell vaccines can cause severe local reactions in individuals with immunity to C. burnetii due to prior infection. A skin test can identify such individuals but increases the cost and complexity of immunization. A more recently developed vaccine, the Chloroform-Methanol Residue Vaccine (CMRV), is less reactogenic than Q-Vax and may eliminate the need for skin testing (Waag et al., 1997). CMRV was developed as a potentially safer alternative. CMRV is safe and sufficiently immunogenic in nonimmune human volunteers (Waag et al., 2008).
Rickettsia prowazekii: Rickettsia prowazekii is a species of Gram-negative bacillus, obligate intracellular parasitic, aerobic bacteria that is the etiologic agent of epidemic typhus, transmitted in the faces of lice. R. prowazekii is the only Rickettsia listed as a potential bioweapon, although appeals for the inclusion of other rickettsial species, most notably R. rickettsi, the causative agent of Rocky Mountain spotted fever, have emerged (Azad and Radulovic, 2003). Epidemic typhus, a major determinant of significant historical turning points, is transmitted to humans by the human body louse and is related to conditions of poor hygiene and social disruption, as outlined by the characteristics of recent epidemics in Burundi and Russia (Raoult et al., 1998). Head lice and flying squirrels have been also recently implicated as R. prowazekii reservoirs (Duma et al., 1981). Human-to-human transmission does not occur. Self-inoculation by scratching the bite-site is the usual mode of transmission, although exposure to aerosol particles containing infected lice faeces also leads to disease. The typical disease induced by R. prowazekii 8 to 12 days after exposure is characterized by fever; severe headache and a subsequent generalized maculopapular, sometimes finally purpuric, rash spreading centrifugally. Mortality is 20% when untreated, significantly higher in older adults and related to gangrene, central nervous system complications, diffuse intravascular coagulation, or severe hypovolemia. Adequate treatment leads to a decline in mortality rates to about 2 to 4%. Establishment of chronic infection can lead to recrudescence (Brill-Zinsser disease) in 15% of the patients, which is usually mild and may appear decades after the typhus episode, often related to external or internal stress. Diagnosis is based on serology (IFA and ELISA), although cross-reactions with other rickettsiae from the typhus group are extensive and their avoidance subject to newer techniques (La Scola and Raoult, 1997). Moreover, the need for convalescent samples allows for a retrospective-only diagnosis. When epidemiologic situations related to the disease have been already recognized, the diagnosis is anticipated and can be achieved clinically (Raoult et al., 1998). Specific PCR techniques for this pathogen have been developed (Carl et al., 1990), including real time PCR (Jiang et al., 2003). Treatment, apart from supportive measures in critically ill patients, relies on the administration of doxycycline, a single dose of which can be life saving (Perine et al., 1992).
Weaponization of R. prowazekii: An older attack scenario developed by WHO outlined the grave sequences of an attack with an aerosolized form of R. prowazekii, with an estimated number of 19,000 deaths. The process of weaponization with R. prowazekii is not as easy, however, as with C. burnetii and one can suppose that such a task could be abandoned in favour of weaponizing a category A pathogen. An alternative approach would use extended spread of infected lice, but the emerging epidemiologic situation would lead to anticipation of epidemic typhus and rapid response (Pappas et al., 2005a). There are currently no vaccines available for epidemic typhus. The whole-cell vaccine that was successful during the Second World War in minimizing epidemic typhus among allied troops is not being produced anymore, but projects for new vaccine products are currently under way (Coker et al., 2003).
| Table 4: | Toxins produced by strains of C. perfringens (Ghabriel et al., 2000) |
 | |
Epsilon toxin of Clostridium perfringens (ETX): Clostridium perfringens is a Gram-positive, rod-shaped, anaerobic, spore-forming bacterium of the genus Clostridium. There are five strains of C. perfringens (Table 4), designated A through E. Each strain produces a unique spectrum of toxins. The epsilon toxin is made by types B and D. This toxin is a pore-forming protein; it causes potassium and fluid leakage from cells. In addition to the epsilon toxin, C. perfringens type D strains produce alpha toxin and type B strains produce alpha and beta toxins. C. perfringens type B causes severe enteritis in young calves, foals, lambs and piglets. Type D causes enterotoxemia in sheep and goats and, on rare occasions, in cattle. All five strains can infect wounds in any species. Epsilon toxin (ETX) is a major toxin produced by an anaerobic bacterium C. perfringens (type B and D) strains and the other types of which cause various human diseases as gas gangrene and necrotizing enteritis. There is a paucity of knowledge over the effect of ETX on humans, because the types of C. perfringens carrying it are not human pathogens and data are largely based on experience from animal disease and mice models (Ghabriel et al., 2000). It is known that ETX exhibits an exquisite neurotropism, indicated by the severe neurologic sequelae of certain C. perfringens type D infections in sheep and the rapid accumulation of ETX in mice brain, resulting in death, after intravenous administration of minimal quantities (Finnie, 2003). This neurotropism may be related to recognition of specific receptors on brain cells. ETX seems to act through alteration of the permeability of the cellular membrane, with formation of a heptameric pore (Nagahama and Sakurai, 1992). Increasing knowledge about protective humoral immunity in infected animals may allow further understanding of the pathophysiology of ETX and further extrapolation on human risk.
Weaponization of EXT: ETX is an aberration in category B pathogen list, because practically nothing is known about its possible consequences in humans. To deliver a large-scale biological attack, it has to be used in the form of an aerosol, or through poisoning of water or food supplies. Yet, because nothing is known about ETX’s interaction with human gastrointestinal and respiratory mucosa, no safe projections could be made over its lethality and its consequences (Nagahama et al., 1998). For example, it is known that goats infected by C. perfringens type D develop enterocolitis and a wasting syndrome that is compatible with the mechanism of action of ETX, yet apart from expecting that the cleavage of the protoxin secreted by C. perfringens by trypsin and chymotrypsin definitely ensues in human gastrointestinal tract as well, no other projections can be made.
Biodefense against EXT: Attempts to develop a recombinant vaccine against ETX are currently under way (Oyston et al., 1998), although a formalin-inactivated vaccine exists for susceptible animals. Treatment would most likely be supportive. Hyperimmune serum might be helpful if given soon after exposure. In mice, a variety of drugs, including reserpine, diazepam, apomorphine, gamma-butyrolactone and phenothiazine and butyrophenone derivatives can prevent death or delay the effects of epsilon toxin, but only if they are given before exposure. A genetically modified toxin can protect mice against lethal challenge. In natural infections with type B strains, hyperimmune serum and antibiotics may be helpful. Toxoid vaccines can prevent type B and D enterotoxemia.
Staphylococcal enterotoxin B (SEB): SEB is an exotoxin produced by S. aureus. It is one of the toxins responsible for staphylococcal food poisoning in humans and has been produced by some countries as a biological weapon. SEB is a superantigen; it acts by stimulating cytokine release and inflammation (Krakauer, 2005). SEB is toxic by inhalation and ingestion. In a biological attack, it could be administered in food, water or as an aerosol. SEB is a prevalent cause of acute diarrheal illness when ingested, causing a self-limited syndrome of undetermined pathogenesis that starts hours after exposure and gradually wanes during the next 72 h. Diagnosis is based on sophisticated toxin assays performed at reference laboratories.
Weaponization of SEB: Its inclusion in category B pathogen list is warranted because of its possible effects when administered in aerosol form, based on limited human data and various experimental animal models. Sixteen cases of inhalational exposure have been documented, resulting rapidly (<24 h) in evolution of a clinical syndrome characterized by fever, malaise, cough, dyspnoea, nausea and vomiting. Ocular exposure in humans can result in purulent conjunctivitis (Rusnak et al., 2004). In animal models exposed to aerosolized form of SEB initial gastrointestinal symptoms were followed by death caused by pulmonary oedema by Day 3 (Mattix et al., 1995). The estimated minimum dose for evoking an emetic response in human volunteers is 5 mg, whereas in aerosolized exposure, the estimated LD50 is 0.02 mg kg-1. Certain aspects of a potential deliberate release of SEB in inhalational form should be addressed, one of the most important being the difficulties posed in differential diagnosis, especially in the context of the absence of widely available diagnostic procedures. A wide variety of both formalin-inactivated and recombinant vaccines have been tested, but none has been approved for human use, although new candidates emerge (Mantis, 2005).
Biodefense against SEB: A wide variety of both formalin inactivated and recombinant vaccines have been tested, but none has been approved for human use, although new candidates emerge (Mantis, 2005). Immune protection through anti-SEB antibodies has been outlined (Boles et al., 2003). Treatment is supportive, although experimental therapeutic procedures are currently under investigation (Krakauer, 2005).
Viruses as biological warfare agents: Most of category B and category C biological warfare agents are viruses. The most important viruses used in weaponization are listed in Table 5.
Hemorrhagic Fever Viruses (HFV): HFVs are caused by viruses of four distinct families: arenaviruses, filoviruses, bunyaviruses and flaviviruses (Table 6). Each of these families shares a number of features:
| Table 5: | Most important viruses used as biological agents (Moran et al., 2008) |
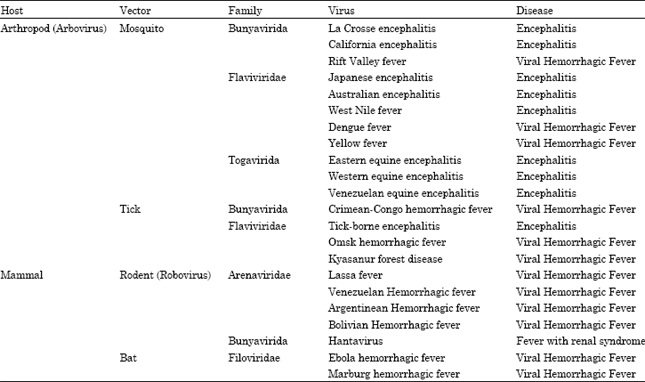 | |
| Table 6: | Hemorrhagic fever viruses (Borio et al., 2002) |
 | |
| • | They are RNA viruses and all are covered, or enveloped, in a fatty coating |
| • | Their survival is dependent on an animal or insect host (natural reservoir) |
| • | The viruses are geographically restricted to the areas where their host species live |
| • | Humans are not the natural reservoir for any of these viruses. Humans are infected when they come into contact with infected hosts. However, with some viruses, after the accidental transmission from the host, humans can transmit the virus to one another |
| • | Human cases or outbreaks of hemorrhagic fevers caused by these viruses occur sporadically and irregularly. The occurrence of outbreaks cannot be easily predicted |
| • | With a few noteworthy exceptions, there is no cure or established drug treatment for VHFs |
The term HFV has referred to a clinical illness associated with fever and a bleeding diathesis caused by a virus belonging to 1 of 4 distinct families: Filoviridae, Arenaviridae, Bunyaviridae and Flaviviridae (Table 6). The HFVs are transmitted to humans via contact with infected animal reservoirs or arthropod vectors. The mode of transmission, clinical course and mortality of these illnesses vary with the specific virus, but each is capable of causing a hemorrhagic fever syndrome. Some HFVs exhibit a significant number of the key features that characterize and pose serious risk as biological weapons, including Ebola and Marburg viruses (Filoviridae), Lassa fever and New World arenaviruses (Arenaviridae), Rift Valley fever (Bunyaviridae) and yellow fever, Omsk hemorrhagic fever and Kyasanur Forest disease (Flaviviridae) (Borio et al., 2002).
Weaponization of VHF: The VHF viruses are spread in a variety of ways. Some may be transmitted to humans through a respiratory route. Although, evidence for a history of Weaponization does not exist for many of these viruses, all are considered by military medical planners to have a potential for aerosol dissemination, weaponization, or likelihood for confusion with similar agents that might be weaponized (Casadevall and Pirofski, 2004).
VHFs have been weaponized by several nations (Borio et al., 2002). HFVs have been weaponized by the former Soviet Union and the US (Alibek and Handelman, 1999). Aerosolized Ebola, Marburg, Lassa, yellow fever viruses and New World arenaviruses can cause disease in nonhuman primates (Borio et al., 2002); however, aerosolized transmission of VHF from person to person has never been documented (Weber and Rutala. 2001). There are reports that yellow fever may have been weaponized by North Korea. The former Soviet Union and Russia produced large quantities of Marburg, Ebola, Lassa and NewWorld arenaviruses (specifically, Junin and Machupo) until 1992 (Miller et al., 2002). Soviet Union researchers quantified the aerosol infectivity of Marburg virus for monkeys, determining that no more than a few virions are required to cause infection (Bazhutin et al., 1992). Yellow fever and Rift Valley fever viruses were developed as weapons by the US offensive biological weapons program prior to its termination in 1969. The Japanese terrorist cult Aum Shinrikyo unsuccessfully attempted to obtain Ebola virus as part of an effort to create biological weapons. Several studies have demonstrated successful infection of nonhuman primates by aerosol preparations of Ebola (Johnson et al., 1995) Marburg (Lub et al., 1995), Lassa (Stephenson et al., 1984) and New World arenaviruses (Kenyon et al., 1992). Arguments asserting that the absence of effective antiviral therapy and vaccines would make these viruses too dangerous to develop as weapons are not supported by the historical record. In 1999, CDC classified the HFVs as category A bioweapon agents, based on the potential to cause widespread illness and death, ease of dissemination or person-to-person transmission, potential for major public health impact and requirement of special action for public health preparedness (Center of Disease Control and Prevention, 2002a, b).
Biodefense against VHF: As a group, VHFs are classified as Category A (Center of Disease Control and Prevention, 2002a, b) agents because of their propensity to cause epidemic disease and high mortality, capacity to be transmitted from person to person and their requirement for specialized preparedness on the part of the public. Vaccines for VHFs such as Ebola and Lassa fever (Fisher-Hoch et al., 2000) and Dengue fever (Holen, 2010) are under development. A promising investigational vaccine for Ebola was recently shown to be highly protective in nonhuman primates (Sullivan et al., 2003). The only licensed vaccine for any of the VHFs is the live-attenuated 17D yellow fever vaccine (Center of Disease Control and Prevention, 2002a, b). The availability of this vaccine makes the potential use of the yellow fever virus as a bioweapon less likely and as such, it has been classified as a Category C bioweapon (Table 8). The yellow fever vaccine is a historically important vaccine that was instrumental in the containment of yellow fever for almost a century. It is recommended for residents of the United States who travel to areas of endemicity in tropical South America, Africa and other countries in the yellow fever endemic zone and for laboratory personnel that are at risk for yellow fever via aerosolization (Center of Disease Control and Prevention, 2002a, b; Monath, 2001). Protective antibody levels arise in 90% of vaccines in 10 days and 99% in 30 days (Weber and Rutala, 2001). A single dose of yellow fever vaccine probably provides lifelong protection, but international health regulations require revaccination 10 years after the initial dose (Monath, 2001). Given the inadequate supply of yellow fever vaccine (Nathan et al., 2003), the lack of vaccines for other VHFs and the potential use of these agents as bioweapons, passive vaccination has been strongly considered to treat and prevent disease (Casadevall, 2002). Passive vaccination with specific antibody is a logical choice for post-exposure prophylaxis because of the short incubation period of many VHF agents, including yellow fever, which makes active vaccination a poor strategy for preventing disease. Passively administered antibody has been used for Ebola, Argentine and Lassa hemorrhagic fevers (Casadevall, 2002).
Arboviruses causing encephalitis: Arboviral (arthropod-borne virus) encephalitis is a group of viral illnesses that cause inflammation of the brain and are transmitted to humans by biting insects. Types of arboviruses include West Nile Virus (WNV), Venezuelan Equine Encephalitis (VEE), Western Equine Encephalitis (WEE) and Eastern Equine Encephalitis (EEE). VEE, WEE and EEE is an alphavirus (Table 7) belongs to the group IV Togaviridae. Alphaviruses have a positive sense single stranded RNA genome. WNV is an in the family Flaviviridae and is the leading cause of arboviral disease in US.
| Table 7: | Medically important category B alphaviruses (Stienlauf et al., 2002) |
 | |
| Table 8: | Approximate assessment of vaccines for human use against CDC categories A, B and C agents (Hilleman, 2002) |
 | |
| IND: Investigative New Drug | |
An estimated 80% of WNV infections are asymptomatic. Most symptomatic persons develop an acute systemic febrile illness that often includes headache, myalgia, arthralgia, rash, or gastrointestinal symptoms. Less than 1% of infected persons develop neuroinvasive disease, which typically presents as encephalitis, meningitis, or acute flaccid paralysis (Centre of Disease Control and Prevetion, 2010). WNV is rarely fatal and most people infected recover. VEE, WEE and EEE are fatal in most cases.
VEE, WEE, EEE and WNV may be used as BWAs (Stienlauf et al., 2002). VEE is the commonest and the most extensively studied pathogen, with a geographic distribution encompassing Latin America and in certain outbreaks reaching US. Human disease is rare, usually in the form of a self-remitting flulike syndrome. Progression to encephalitis occurs in 1% of the adults and 4% of the paediatrics population, with 20% mortality. EEE is principally located in US, east of the Mississippi River, but principally animal disease has also been reported from the Caribbean and Latin America. Incubation period is 5 to 7 days. The ratio of self-remitting or nonapparent to severe infections is 40:1 for adults and 17:1 for the paediatric population. The clinical syndrome is similar to VEE, but mortality is much higher, with an additional number of survivors exhibiting severe neurologic sequelae (Calisher, 1994). WEE is distributed in various western territories of US and Canada, but also in regions of Latin America. The ratio of self-remitting or nonapparent to severe infections is 1150:1 for adults, but only 1:1 for infants. As with EEE, the disease has a seasonal distribution, probably related to the increased outdoor activities during the summer period. The clinical syndrome induced, after an incubation period of 5 to 10 days, is particularly severe in infants, with common neurologic sequelae present in survivors of this age group (Moran et al., 2008).
Weaponization of viral encephalitis: The attractiveness of VEE as a bioweapon is based on its potential for widespread infection through aerosolization or release of infected mosquitoes, its relatively low infective dose for humans, its ease of production and its implications for animal populations of the targeted area (Bronze et al., 2002). Studies on weaponization of VEE have allegedly taken place in the past. A particular disturbing scenario regarding VEE’s low attack rates (at least in the form of severe disease) might include genetic manipulation and evolution of highly neurotropic strains. Prophylaxis through vaccination is not available for civilians: an inactivated vaccine is available as an investigational new drug and live attenuated vaccines are in development (Lee et al., 2005). The weapon potential of EEE is roughly equal to that of VEE, its higher infective dose being compensated by the relative severity of the clinical syndrome. Pre-exposure prophylaxis through vaccination is not available for the public, but only through the US Army. On the contrary, the pathogenetic characteristics of WEE make it a paediatrics bioweapon (Schoepp et al., 2002). CDC and ATCC were sent WNV and many BWAs to Iraq up until 1989. These BWAs were used for Iraq's bioweapons research program.
Biodefense against viral encephalitis: There is no approved therapy for the agents of viral encephalitis and care remains supportive. LAV for VEE, TC-83, has been used in horses since 1961 (Cieslak et al., 2000). This vaccine has not been approved for humans because it is highly reactogenic in up to 20% of vaccines, potentially teratogenic, diabetogenic and immunogenic in only 60% of recipients. Efforts to reduce the side effects of the vaccine through genetic engineering have resulted in the development of recombinant VEE vaccines in vaccinia vectors (Phillpotts et al., 2000). A new vaccine, V3526, is a live-attenuated virus derived by site-directed mutagenesis from a virulent clone of VEV, intended for human use in protection against Venezuelan equine encephalitis at dose as low as 10 (2) pfu, were safe (Fine et al., 2007). Mucosal immunity may have critical importance for protection against airborne VEE (Elvin et al., 2002) since increased protection was achieved with aerosolized administration of a live-attenuated VEE vaccine candidate (Hart et al., 2000). The development of EEE and WEE vaccine candidates is being pursued (Schoepp et al., 2002). Passive vaccination approaches have been bolstered by the development of murine monoclonal antibodies to VEE that were protective against aerosolized VEE in mice (Phillpotts et al., 2002).
Foodborne and waterborne pathogens
The bacteria: Most of the known bacterial causes of infectious diarrhoea are included in category B pathogen list. Most of them are universally widespread and responsible for millions of annual cases worldwide.
Salmonella species can induce acute infectious diarrhoea that manifests usually within 1 day after exposure, demand a low inoculum of almost 100 colony forming units and can be treated only with supportive measures. More severe cases may demand the use of co-trimoxazole, quinolones, or ceftriaxone. Salmonella typhi is the causative agent of typhoid fever, a protracted systematic illness that, at least in experimental studies, demands high inoculums (Glynn et al., 1995). The incubation period ranges from 7 to 14 days. The disease causes complications in the gastrointestinal tract or the central nervous system in 10% of patients, but the overall case-fatality rate is less than 2%. Blood and bone marrow cultures are the preferential diagnostic methods, the latter reaching a sensitivity of 95% (Parry et al., 2002).
Shigella species may be the most formidable food-borne and water-borne pathogen, because it can induce bloody diarrhoea with a very low inoculums; can further be transmitted from person to person through the faecal-oral route and is related to the late development of haemolytic uremic syndrome. Its incubation period is 1 to 7 days. The importance of Shiga’s toxin as a prototype toxin cannot be overemphasized (Niyogi, 2005).
Enteropathogenic E. coli strains can be implicated in acute watery diarrhoea, but also in a low-inoculum-induced, often afebrile (50%), bloody (in 90% of the patients) diarrhoea by Shiga’s toxin-producing strains (Qadri et al., 2005), the importance of which is paramount, because the disease is also related to the development of haemolytic uremic syndrome in a significant percentage of children. Rapid diagnosis is imperative, because antibiotics may further predispose to haemolytic uremic syndrome development, although other reports suggest the opposite (Boyce et al., 1995). Attack rates for Shiga’s toxin-producing strains are estimated at 20%.
Campylobacter jejuni is the most prevalent bacterial cause of infectious diarrhoea in adults, with an incubation period of 1 to 7 days and a very high ratio of subclinical to clinical cases, possibly because of its exquisite susceptibility to hydrochloric acid. It is also related to certain sequelae, as Guillain-Barre syndrome (Nachamkin et al., 1998). Macrolides are first-line therapeutic choices, especially because quinolone resistance to the pathogen has been continuously on the rise (Butzler, 2004).
Vibrio cholerae has been historically responsible for a huge number of epidemics worldwide (Kaper et al., 1995), inducing through an exotoxin a severe watery diarrhoea that leads to hypovolemia and death in certain clinical settings. Achlorhydria is also a risk factor for symptomatic disease. The attack rates are high, however and treatment, beyond aggressive fluid and electrolyte resuscitation, may be achieved with a single dose of doxycycline or ciprofloxacin. V. parahemolyticus is the commonest pathogen implicated in vibrio-related diarrhoea in the Western world, causing in less than 24 h after exposure a usually benign gastroenteritis, although septicaemia might develop in immunocompromised patients and patients with underlying liver disease. It requires a high inoculum. Treatment is largely supportive and in severe cases doxycycline and quinolones can be used (Butt et al., 2004).
Y. enterocolitica is a zoonotic pathogen largely localized to Northern Europe, which usually causes acute enteritis, especially in young children. The severity of infection (septicaemia associated with 50% mortality) is related to host factors, namely iron overload as in thalassemia, underlying liver disease, diabetes mellitus and old age; the disease may be more common in African Americans and infants in the United States. Isolation of the organism suggests the diagnosis, although serology might be helpful. Treatment is usually suggested for immunocompromised patients and severe infections, using combinations of doxycycline, co-trimoxazole, aminoglycosides, or quinolones (Ray et al., 2004).
The viruses: Norovirus causes approximately 90% of epidemic non-bacterial outbreaks of gastroenteritis around the world and may be responsible for 52% of all foodborne outbreaks of gastroenteritis in the US. The inoculum is low (<100 viral particles) and attack rates exquisitely high, reaching 100%. Further human-to-human transmission is feasible, especially in the presence of severe symptoms. Incubation period is hours to 2 days and supportive measures are usually the only indicated treatment. One important aspect of Noroviruses is their ability to exist in aerosolized form and their relative environmental stability, because they are resistant to common disinfectants (Musher and Musher, 2004).
Hepatitis A virus has been related to massive epidemics because of consumption of infected seafood, surviving from common hygiene measures of potable water supplies. The inoculum for induction of infection is low (<100 viral particles) and the incubation period is 2 to 6 weeks. The resulting acute hepatitis is benign (0.2% mortality in patients who developed jaundice) and does not exhibit chronicity (Cuthbert, 2001).
The protozoa: Cryptosporidium parvum is an intracellular waterborne pathogen that has caused few significant epidemics (Mac Kenzie et al., 1994), but is of main importance in the pathology of AIDS. Its cysts are environmentally stable and a small number of cysts (even one to two) are required to induce infection. Incubation period is 1 to 14 days. Immunocompetent patients are usually symptom-free or exhibit limited symptoms, but the disease is more severe in immunocompromised patients. Human-to human secondary transmission is feasible. Treatment is advocated for severe cases and paromomycin is the agent of choice, although new treatment options are emerging (Smith and Corcoran, 2004).
Toxoplasma gondii is an intracellular food-borne pathogen, which poses significant risks for patients with AIDS, while also adversely affecting the outcome of the fetus when non-immune pregnant mothers are infected. Rare cases of chorioretinitis are the most significant presentations in immunocompetent adults. The evolution of PCR has augmented accurate diagnosis. Treatment, when needed, is based on combinations of pyrimethamine and sulfadiazine or clindamycin (Montoya and Liesenfeld, 2004).
Food-borne and waterborne bacteria can be implicated in potential bioterrorism events because, besides air, food and water supplies are the other options for accessing massive numbers of candidate patients (Elad, 2005). In 1984, a religious group used S. typhimurium to contaminate salad bars to influence the outcome of a regional election (Torok et al.,1997) and an attempt of poisoning using Shiga’s toxin was later reported (Kolavic et al., 1997). Category A pathogens can also be implicated in poisoning of the food and water chain (Wein and Liu, 2005). Certain agents are not that well known, so it is difficult for extremist groups to develop them for dispersion: category B includes both Shigella, which might be a primary extremist choice. Vaccine availability for foodborne and waterborne pathogens is limited at present. Commercially available vaccines exist for typhoid, cholera and hepatitis A virus (Lucas et al., 2005; Parry et al., 2002).
Plant as biological agents: Ricin, the toxic glycoprotein of the castor oil plant's seed (Ricinus communis) recently has earned a great significance, as a potential biological weapon. It is regarded as an ideal BA for biological warfare and bioterrorism (Franz and Jaax, 1997) because of its notoriety arising from the high profile assassination of a leading communist dissident in London (Schep et al., 2009). Furthermore, it is readily accessible and its relative ease of extraction from the castor bean plant, as well as its stability in both hot and cold conditions (Center of Disease Control and Prevention, 2004), seem to make it a weapon of choice. It has been regarded as one of the most potent poisons (LD50 is 3 μg kg-1) in the plant kingdom and has been described as a toxin that can cause death within minutes of exposure (Lee and Wang, 2005). Ricin (64 KDa) consists of a neutral A-chain (32 kDa) bound by a disulfide bond to an acidic B-chain (32 kDa). The B-subunit binds to glycoproteins on the surface of epithelial cells, enabling the A-subunit to enter the cell via receptor-mediated endocytosis. This subunit inactivates ribosomal RNA by depurinating a specific ribosomal residue, thereby inhibiting protein synthesis. One ricin molecule can inactivate 2000 ribosomes per minute, which ultimately leads to the death of the cell (Lord et al., 1994). Ricin has no selectivity for specific cells. Since, all types of cells are affected, symptoms depend on the locus of entry of the toxin into the body. Depending on the dose, first symptoms occur after a latent period of 2-24 h. After inhaling significant amounts of ricin, the victim may suffer from breathing difficulties, fever, cough, nausea and tightness in the chest. Heavy sweating may follow as well as pulmonary oedema. This makes breathing even more difficult and the victim becomes cyanotic. Finally, low blood pressure and respiratory failure may occur, leading to death. After oral ingestion, victims develop vomiting and diarrhoea that may become bloody. Severe dehydration and low blood pressure are the consequence. In severe cases, victims suffer from hallucinations and seizures. Within a few days, the victim's liver, spleen and kidneys may stop working. Fatalities from ricin poisoning may ensue within 36-72 h after exposure, depending on the dose and route of administration (Bigalke and Rummel, 2005).
Weaponization of ricin toxin: In recent years there has been an increased concern regarding the potential use of biological weapons for mass urban terror. In particular, there are concerns that ricin could be employed as such an agent. This has been reinforced by recent high profile cases involving ricin and its use during the cold war to assassinate a high profile communist dissident. Ricin is a suitable weapon for assassination (Schep et al., 2009).
The Weir Wood reservoir, which supplies water to approximately 60,000 residents in Sussex, England, has a capacity of 1237 million L. To achieve the required lethal concentration, approximately 7422 kg of pure ricin powder would need to be introduced to the reservoir (Mackinnon and Alderton, 2000). Terrorists may, however, seek to contaminate water to strategic targets such as houses of parliament or military facilities.
Dermal application of ricin has been considered an alternate route of ricin toxicity. Members of the Minnesota Patriots Council mixed ricin with DMSO and planed to smear doorknobs or items of clothing to assassinate unspecified individuals (Tucker, 1999). Parenteral delivery of ricin can be associated with a greater mortality rate, as indicated by the limited number of case reports in humans (De Paepe et al., 2005). By injection, ricin is a suitable weapon for assassination however, a scenario involving parenteral administration to a large urban population is clearly not reasonable (Schep et al., 2009).
Category C biological warfare agents: The agents classified by the CDC as category C are emerging pathogens that could someday be engineered for mass exposure because of availability, ease of production and dissemination and potential for high morbidity and mortality. Preparedness for category C agents requires ongoing research to improve disease detection, diagnosis, treatment and prevention. Which newly emergent pathogens terrorists might use is impossible to know in advance. Category C including hantaviruses, multidrug-resistant tuberculosis, Nipah virus, the tick-borne encephalitis viruses, the tick-borne hemorrhagic fever viruses and yellow fever (Moran et al., 2008).
Nipah virus: Nipah virus is negative strand RNA virus and belongs to Paramyxoviridae. Nipah virus causes severe illness characterized by inflammation of the brain (encephalitis) or respiratory diseases. Nipah virus can be transmitted to humans from animals and can also be transmitted directly from human-to-human (not proved); in Bangladesh, half of reported cases between 2001 and 2008 were due to human-to-human transmission. Nipah virus can cause severe disease in domestic animals such as pigs. There is no treatment or vaccine available for either people or animals. Fruit bats of the Pteropodidae family are the natural host of Nipah virus. In April 1999, an outbreak of 257 cases of encephalitis (100 fatal) was reported in Malaysia (Centers for Disease Control and Prevention, 1999). A previously unrecognized paramyxovirus called Nipah was identified as the cause. Pigs appeared to be the primary source of human infection in this outbreak. Patients in the reported outbreak presented with fever, headache and myalgias and eventually developed signs of meningitis or encephalitis. A few patients had respiratory symptoms. Identification of Nipah virus requires specialized testing in a reference laboratory, such as the CDC or USAMRIID. IgM antibodies can be detected in blood and CSF. Better diagnostic tests for this recently discovered agent are under development (Daniels et al., 2001). Infection control precautions person-to-person spread of Nipah virus has not been identified. However, virus has been isolated from respiratory secretions and urine of patients infected with Nipah virus (Chua et al., 2001). Pending further study of the potential for person-to-person spread, strict isolation would be prudent for patients suspected of being infected with this virus.
Hantaviruses: Hantaviruses are found in many rodent species worldwide. These rodent-borne viruses are members of the family Bunyaviridae and are transmitted to humans in aerosols of rodent excreta. There are more than 20 recognized Hantaviruses, some of which are associated with one of two serious human diseases: hemorrhagic fever with renal syndrome (HFRS) or Hantavirus Pulmonary Syndrome (HPS) (Schmaljohn, 2009).
The existence of Hantaviruses in the US was known since 1983, when Prospect Hill Virus (PHV) was isolated from a meadow vole in Frederick, MD (Lee et al., 1985). The first appearance of an acute Hantavirus associated disease in the New World occurred in 1993 when a cluster of cases of adult respiratory distress were observed in the South-Western US. Within days of obtaining clinical specimens, the disease was shown to be caused by a previously unrecognized Hantavirus, now known as Sin Nombre Virus (SNV), which was carried by deer mice (Nichol et al., 1993). After the discovery of SNV, several other pathogenic and presumably non-pathogenic Hantaviruses were identified in various rodent species throughout the New World. None of the North American Hantaviruses have been shown to be transmitted person-to-person; however, there is evidence that the South American virus andes virus, can occasionally be contagious among humans (Toro et al., 1998). The disease caused by these hantav-ruses, HPS, is sometimes referred to as Hantavirus cardiopulmonary syndrome to recognize the cardiogenic shock often associated with the disease (Hallin et al., 1996).
Weaponization of hantavirus: Hantaviruses are assigned to categories A and C on the National Institute of Allergy and Infectious Diseases priority pathogens list and to category C on CDC emergency preparedness and response list of bioterrorism agents/diseases (Schmaljohn, 2009). In spite of the infections by Hantavirus are rare, Hantavirus is the most pathogenic virus. Aerosols of virus-contaminated rodent urine or feces seemed to be the mechanism of transmission in these cases. Unavailability of vaccine and due to aerosol transmission is possible makes the virus is believed to have potential for weaponization. Hantaviruses and diseases that they cause have a long association with war and military operations. It is suspected that the 7000 cases of war nephritis reported among British soldiers stationed in Flanders during World War I were caused by Hantavirus infections. In the early 1930s, Russian troops encountered a disease with nephritis, bleeding and shock along the Amur River, which forms the border between far-eastern Russia and Manchuria. Around the same time, the Japanese Army suffered more than 12,000 cases of a similar disease when they invaded Manchuria (Johnson, 2001). Epidemics have also occurred; for example, The Republic of Bashkortostan reported more than 10,000 cases of HFRS, with an incidence rate of 287 per 100,000 in 1997 (Nurgaleeva et al., 1999).
Biodefense against hantavirus: Hantaviruses are distributed throughout the world and cause serious human diseases, which pose particular threats to military personnel who might come in contacts with the rodents in the field operation. According to this situation urgent needs to vaccines protect Hantavirus infections. To date, Hantavirus vaccines have not been tested in humans. A phase I clinical study of Hantavirus vaccines is in progress. It is possible to reduce Hantavirus infections through rodent control and through exercise of prequations when cleaning rodent infested area (Schmaljohn, 2009).
Other agents: Several arthropod-borne viruses might have potential for use as biological weapons, including the flaviviruses that cause yellow fever and tick-borne encephalitis. Person-to-person transmission of flaviviruses does not appear to occur, except through the arthropod vectors.
Yellow fever is a mosquito-borne virus of historical interest because of large outbreaks that played a role in development of the Americas. The disease has been greatly diminished through mosquito control and vaccination, although sporadic outbreaks still occur. The severity of illness can range from a mild self-limited viral syndrome to a fatal hemorrhagic fever (Monath, 2001). After an incubation period of several days, symptoms begin as fever, headache and myalgias. Conjunctivitis, relative bradycardia and leukopenia may be present. Jaundice occurs secondary to hepatitis and gastrointestinal bleeding may also occur. Death can occur 7 to 10 days after onset.
| Table 9: | WHO estimates of casualties produced by attacks with biological agents (Hilleman, 2002) |
 | |
Treatment of yellow fever is supportive. The illness is preventable with the attenuated 17D vaccine, which produces immunity in approximately 95% of those vaccinated (Moran et al., 2008).
Tick borne encephalitis is caused by Tick-Borne Encephalitis Virus (TBEV), a member of the family Flaviviridae and was initially isolated in 1937. Three virus sub-types are described: European or Western TBEV, SiberianTBEV and Far eastern TBEV. Tick-borne encephalitis occurs in many areas of Europe and Asia. Infection can also occur from consumption of unpasteurized milk products. Most infections are asymptomatic or only mildly symptomatic, but a small fraction of infected individuals can develop encephalitis. Only approximately 1% of encephalitis cases are fatal, mostly in elderly individuals (Tsai, 2000). No specific therapy exists for flavivirus encephalitis.
Multidrug-resistant tuberculosis has become a significant problem in many areas of the world over the past several decades. Although, illness progression and person-to-person transmission occur slowly, the ability to disseminate through aerosol and difficulty treating multidrug-resistant strains could make the organism attractive as a bioweapon. Treatment options for highly resistant strains are severely limited (Small and Fujiwara, 2001).
Rank order of killing capability of biological agents: Thirty years ago, a committee of WHO (Table 9), prepared a rank order for the killing capability of 50 kg of each of 7 agents not including smallpox, delivered upwind from a city of 500,000 populations. Brucellosis, Q fever, tularaemia and anthrax greatly outranked rift valley fever, tick-borne encephalitis and typhus with respect to numbers of persons incapacitated. The three most lethal agents were anthrax, tularaemia and typhus.
Delivery of biological warfare agent: BWAs can be disseminated in various ways (1). Through the air by aerosol sprays: To be an effective biological weapon, airborne germs must be dispersed as fine particles. To be infected, a person must breathe a sufficient quantity of particles into the lungs to cause illness. (2) Used in explosives (artillery, missiles, detonated bombs): The use of an explosive device to deliver and spread BAs is not as effective as the delivery by aerosol. This is because BWAs tend to be destroyed by the blast, typically leaving less than 5% of the agent capable of causing disease. (3) Put into food or water: Contamination of water supplies requires an unrealistically large amount of an agent as well as introduction into the water after it passes through a regional treatment facility. (4) Absorbed through or injected into the skin. This method might be ideal for assassination, but is not likely to be used to cause mass casualties (Moran et al., 2008).
BWAs may be delivered in either wet or dry form. Dry powders composed of very small particles tend to have better dissemination characteristics and have advantages in storage. Dried agents require an increased level of technological sophistication to produce, although freeze drying or spray drying technology has been available in industry for a number of years. BWAs might be released by an aggressor against our forces or against civilian populations by means of sprays, explosive devices and contamination of food and water.
Most commonly, delivery methods use aerosolized agent. The agent can be dispersed by attaching a spray device to a moving conveyance. An industrial insecticide sprayer designed to be mounted on an aircraft is an example. A line of release would then occur while the sprayer is operating. This is known as a line source and is sprayed perpendicular to the direction of the wind, upwind of the intended target area. Up to a certain range, anyone downwind of such a line source would theoretically be at risk. The range that the infectious or toxic agent would reach depends on a number of factors, including wind speed and direction, atmospheric stability and the presence of inversion conditions and on characteristics of the agent itself (e.g., stability to desiccation or ultraviolet light). Studies using computer models have shown that clouds of hardy organisms such as anthrax spores can be infectious more than 200 km from the source of the aerosol. A second type of aerosol source is a point source, which is a stationary device for aerosolization of the agent, such as a stationary sprayer. A modified point source would be a group of spray devices, such as specially designed bomblets dispersed in a pattern on the ground by a missile or artillery shell designed to release such bomblets. Many such devices have been designed; an example is the Flettner rotor. Several types of spherical bomblets have also been widely tested and most biological bomblets are spherical (Fig. 2). Bomblets may be designed to disseminate on imimpact or at a predetermined altitude above the ground. They may be released from missiles or aircraft and may have special designs to improve their aerodynamics or pattern in the target area. Other types of delivery systems for BAs have been designed by various countries. These include bombs or bomblets that release the agent by exploding (generally very inefficient delivery systems), land and sea mines, pipe bombs and many other special devices. Clandestine means of delivering BWAs are potentially available to terrorists or Special Forces units; these include devices that penetrate and carry the agent into the body via the percutaneous route, such as pellets or flechettes, or mean to contaminate food or water supplies so that the agent would be ingested.
Infective aerosol dose of biological agents: US military prepared a list of 10 likely BWA (Table 10) for which prophylactic immunization or rapid diagnosis and treatment can have an impact on outcome (Franz et al., 1997).
| Table 10: | US military list of 10 likely biological agents for which rapid diagnosis and treatment can reduce impact (Hilleman, 2002) |
 | |
| aRemoved from market. bUsed under Investigative New Drug (IND) Law | |
 | |
| Fig. 2: | E120 biological bomlet, developed before the US signed the biological and toxic weapons convesion |
As shown, there were wide differences in the required infective dose of microorganisms needed to kill. For seven of these agents, there are credible licensed vaccines or vaccines that may be worthy of use under IND application. For the five bacterial agents, there are antibiotics. For two viral agents, there are antiviral drugs of possible value. Polyvalent immune globulin exists for treating botulinus poisoning. Possibly, specific immune sera will be effective in treating against HFVs.
Role of biotechnology in biological warfare: Rapid developments in biotechnology, genetics and genomics are undoubtedly creating a variety of environmental, ethical, political and social challenges for advanced societies. But they also have severe implications for international peace and security because they open up tremendous avenues for the creation of new biological weapons. By using genetic engineering, biological researchers have already developed new weapons that are much more effective than their natural counterparts. Such weapons, designed for new types of conflicts and warfare scenarios, secret operations or sabotage activities, are not mere science fiction, but are increasingly becoming a reality that we have to face. In this review provides a systematic overview of the possible impact of biotechnology on the development of biological weapons.
Military experts are perfectly aware of the danger of genetically engineered bioweapons, as their traditional defence measures, e.g. detection methods or vaccines, are easily sidestepped by the artificial microbes. The speedy development of genetic engineering is one driving force to strengthen the Bioweapons Convention and establish a verification system (Borzenkov et al., 1993).
Some of applications of biotechnology in biological weapons
Example 1: Bacteria causing unusual symptoms: Researchers from Obolensk near Moscow inserted a gene into F. tularensis. The gene made the bacteria produce beta-endorphin, an endogenous human drug, which caused changes in the behaviour of mice when infected with the transgenic bacteria (Borzenkov et al., 1993). According to the published results, the endorphin gene was not introduced into a fully virulent strain, but only into a vaccine strain. If inserted into virulent F. tularensis, the victims would not show the usual symptoms of tularaemia, but instead unusual symptoms that would obscure the diagnosis and delay therapy. Development of symptom-altered BWAs has been identified as one possible application of genetic engineering for BW purposes by the US Department of Defence.
Example 2: Transferring a lethal factor to harmless human gut bacteria: Genetic engineering could make previously harmless bacteria lethal biological weapons by introducing deadly genes from a highly pathogenic organism. This was done by US researchers as early as 1986. They isolated the gene for the lethal factor of B. anthracis and introduced into E. coli (normal flora). The US team reported that the lethal factor protein was active in E. coli and displayed the same deadly effects as it did when in its native B. anthracis (Robertson and Leppla, 1986).
Example 3: Antibiotic resistant anthrax and tularaemia: Antibiotic resistance is often used as a marker gene in genetic engineering experiments. However, the very same genes could render bioweapons more dangerous by making agents less treatable. Any experiment with BWAs using antibiotic resistance genes has a strong offensive potential, even if in the context of defensive research. German military researchers at the Santitaetsakademie der Bundeswehr in Munich, the main BW research facility of the German army, cultured genetically engineered F. tularensis subsp. holarctica bacteria, a close relative of the causative agent of tularaemia. An antibiotic resistance marker gene (tetracycline) was been inserted into these bacteria (DaSilva, 1999).
Recently, researchers from Porton Down in the UK used genes conferring resistance to antibiotics for genetic studies in fully virulent strains of anthrax (Bowen and Quinn, 1999). In the late 1980s, a researcher at the University of Massachusetts in Amherst also introduced antibiotic resistance genes into anthrax, making it less treatable with antibiotics (Nass, 1991).
Researchers from the Institute Pasteur in Paris (Pezard et al., 1993) and from a Russian laboratory in Obolensk (near Moscow) (Pomerantsev and Staritsyn, 1996) introduced antibiotic resistance genes into anthrax bacteria. All these studies are allegedly basic research, where antibiotic resistance is used as a marker gene. But it is obvious that the very same genetically engineered bacteria can be used to design more effective bioweapons compared to the natural anthrax strains.
Example 4: Invisible anthrax: In December 1997, the same Russian research group from Obolensk published a paper in a British scientific journal on another effort to genetically engineer anthrax (Pomerantsev et al., 1997). By putting new genes into fully pathogenic strains of anthrax, the scientists altered anthrax�s immunopathogenic properties, making existing anthrax vaccines ineffective against the new genetically-engineered types. In most cases, detection of bioweapons relies on molecular recognition of the microbe using antibodies similar to the human immune system. Altering the immunogenicity not only overcomes vaccinations; but also the detection systems (DaSilva, 1999). Western military experts were alarmed by this work. The chief of the bacteriology division at the US Army Medical Research Institute of Infectious Diseases (USAMRIID) in Fort Detrick, Md, Col. Arthur Friedlander, commented: This is the first indication we're aware of in which genes are being put into a fully virulent strain. They genetically engineered a strain that's resistant to their own vaccine and one has to question why that was done. The Russian researchers also constructed a new vaccine against the new strain. This is of particular importance, as it could enable an army to use such a bioweapon by vaccinating their soldiers against a specific strain, while the enemy remains vulnerable. The case is an example of the frightening potential of genetic engineering applied to biological weapons research (Aken and Hammond, 2003).
Concluding remarks and summary: BW is one of the mass destruction weapons and considered the weapon of poor nations because BWA is easily obtained and produced in large quantities with relatively low cost than other mass destruction weapons such as nuclear and chemical. BWA is mainly living organism or toxins produced by those organisms. When BWA is used by nations is called BW while used by non-national groups is called bioterrorism. The important tool of BW is BWA and Human discovered the BWAs from ancient times and may use those BWAs in the biological weapons over the past ages. Human developed everything in the life and therefore it was developing the use of those BWA in BW. In ancient history he used the BWAs in crude forms such as use of snake venom to pollute arrow to infect the enemy. In the last century, human use sophisticated preparation developed for use in BW. BA may be bacteria, viruses and toxins. BWAs categorized into three categories and the most virulent is category A. Multiple BWAs have been classified by the CDC as potential weapons of mass destruction or agents for biologic terrorism. Bacterial agents such as B. anthracis, Y. pestis and F. tularensis are belonging to category A. However, Brucella, B. mallei, B. pseudomallei, C. psittaci, R. prowazekii and C. burnetii are categorized in category B. Some of the bacterial agents such as multi-drug resistant TB is example of category C. Toxins also used in BW or bioterrorism. Some of toxins obtained from microorganism, such as BoNT (category A), EXT (category B) and SEB (category B) and toxin obtained from plant such as ricin (category B). Multiple viral agents have been classified by the CDC as potential weapons of mass destruction or agents for biologic terrorism. Agents such as smallpox and viral hemorrhagic fever viruses are belonging to category A. Category B viral agents include viral encephalitis. Hantaviruses, nipah viruses, TBHF, TBEV and yellow fever virus are examples of category C and others are of concern because they are highly infectious and relatively easy to produce. Biotechnology is considered as a tool of BW and applications of this science in the production, modification and manufactured of BWAs is introduced in the last four decades and the applications of biotechnology and genetic engineering in the field of BW and bioterrorism make BWAs more serious. For example, B. anthracis is susceptible to penicillin due to lack of penicillinase. However, transformation of blagene into B. anthracis makes this strain resistant to penicillin. E. coli is a normal flora in human and animals, transferring the lethal factor from B. anthracis into E. coli make it virulent as B. anthracis. Therefore, misuse of this technology in BW leads to production of new generation of BWAs which are more virulent. Another tool of BW is delivery of BWAs. Dissemination and delivery of BWAs may occur by aerosol sprays, explosives (artillery, missiles, detonated bombs), or food or water contamination. Variables that can alter the effectiveness of a delivery system include particle size of the agent, stability of the agent under desiccating conditions, UV light, wind speed, wind direction and atmospheric stability. The use of an explosive device to deliver and disseminate BWAs is not very effective, since such agents tend to be inactivated by the blast. Contamination of municipal water supplies requires an unrealistically large amount of agent and introduction into the water after it passes through a regional treatment facility. To be an effective bioweapon, airborne pathogens must be dispersed as fine particles less than 5 ì m in size. Infection with an aerosolized agent usually requires deep inspiration of an infectious dose. Advanced weapons systems (e.g., warheads, missiles) are not required for the aerosolized delivery of BWAs. Low-technology aerosolization methods including agricultural crop-dusters; aerosol generators on small boats, trucks, or cars; backpack sprayers; and even purse-size perfume atomizers suffice. Aerosolized dispersal of BWAs is the mode most likely to be used by terrorists and military groups.
Biodefnce against BWAs is another tool in BW, achieved by vaccination, prophylaxis and treatment. Protective measures should be implemented early (if warning is received) or later (once suspicion of BWA use is made). Currently, available masks such as the military gas mask or High-Efficiency Particulate Air (HEPA) filter masks used for TB exposure filter out most BW particles delivered by aerosol. Multilayered HEPA masks can filter 99.9% of 1- to 5 μm particles, but face-seal leaks may reduce the efficacy by as much as 10-20%. Individual face-fit testing is required to correct seal leak problems. Most aerosolized BWAs do not penetrate unbroken skin and few organisms adhere to skin or clothing. After an aerosol attack, simple removal of clothing eliminates a great majority of surface contamination. Thorough showering with soap and water removes 99.99% of the few organisms left on the victim's skin after disrobing. Of the potential BWAs, only plague, smallpox and VHF are spread readily person to person by aerosol and require more than standard infection control precautions (gown, mask with eye shield, gloves). Regardless, place all potential victims of BWAs in isolation. Medical personnel caring for these patients should wear a HEPA mask in addition to standard precautions pending the results of a more complete evaluation. Broad-spectrum intravenous antibiotic coverage is recommended initially for victims when a BWA is suspected. Institute this even prior to the identification of the specific BW agent. Vaccinations currently are available for anthrax, BoNT, tularaemia, plague, Q fever and smallpox. The widespread immunization of non-military personnel has not been recommended by any governmental agency. Immunization against many of BWAs are not available until now or vaccine under IND. Unavailability of immunization against these BWAs make these BWAs are very dangerous such as VHF viruses, VEE, EEE, WEE, SEB, BoNT, tularaemia and Q fever. On the other hand, certain vaccines, such as anthrax vaccine, are accepted component of post exposure prophylaxis against potential BWAs. Some vaccines have drawbacks; (1) vaccines are not accepted component of post exposure prophylaxis against potential BWAs. For example C. perfringens vaccine (IND). (2) vaccines are affective against certain type of infection and ineffective against other type of infections. For example plague vaccine have ability to protect soldiers against endemic plague and it may not protect adequately against acquisition of disease by the aerosol route.
REFERENCES
- Aken, J.V. and E. Hammond, 2003. Genetic engineering and biological weapons. EMBO Rep., 4: S57-S60.
CrossRef - Arnon, S.S., R. Schechter, T.V. Inglesby, A. Donald and M.D. Henderson et al., 2001. Botulinum toxin as a biological weapon: Medical and public health management. JAMA, 285: 1059-1070.
CrossRefPubMedDirect Link - Azad, A. and S. Radulovic, 2003. Pathogenic rickettsiae as bioterrorism agents. Ann. N. Y. Acad. Sci., 990: 734-738.
PubMed - Baillie, L. and T.D. Read, 2001. Bacillus anthracis: A bug with attitude. Curr. Opin. Microbiol., 4: 78-81.
PubMed - Bandouchova, H., J. Sedlackova, M. Hubalek, M. Pohanka and L. Peckova et al., 2009. Susceptibility of selected murine and microtine species to infection by a wild strain of Francisella tularensis subsp. holoarctica. Vet. Med., 54: 64-74.
Direct Link - Bartlett, J., L. Borio, L. Radonovich, J.S. Mair and T. O'Toole et al., 2003. Smallpox vaccination in 2003: Key information for clinicians. Clin. Infect. Dis., 36: 883-902.
PubMed - Bartlett, J.G., Jr. T.V. Inglesby and L. Borio, 2002. Management of Anthrax. Clin. Infec. Dis., 35: 851-858.
CrossRefDirect Link - Bazhutin, N.B., E.F. Belanov, V.A. Spiridonov, A.V. Voitenko and N.A. Krivenchuk et al., 1992. The effect of the meth-ods for producing an experimental Marburg virus infection on the characteristics of the course of the disease in green monkeys. Vopr. Virusol., 37: 153-156.
PubMed - Beyer, W., S. Pocivalsek and R. Bohm, 1999. Polymerase chain reaction-ELISA to detect Bacillus anthracis from soil samples: Limitations of present published primers. J. Applied Microbiol., 87: 229-236.
PubMed - Bhalla, D.K. and D.B. Warheit, 2004. Biological agents with potential for misuse: A historical perspective and defensive meseaures. Toxicol. Applied Pharmacol., 199: 71-84.
CrossRef - Bianucci, R., L. Rahalison, E.R. Massa, A. Peluso, E. Ferroglio and M. Signoli, 2008. Technical note: A rapid diagnostic test detects plague in ancient human remains: An example of the interaction between archeological and biological approaches (southeastern France, 16th-18th centuries). Am. J. Phys. Anthropol., 136: 361-367.
PubMed - Boles, J.W., M.L. Pitt, R.D. LeClaire, P.H. Gibbsa and E. Torres et al., 2003. Generation of protective immunity by inactivated recombinant staphylococcal enterotoxin B vaccine in nonhuman primates and identification of correlates of immunity. Clin. Immunol., 108: 51-59.
CrossRef - Borio L., T. Inglesby, C.J. Peters, A.L. Schmaljohn and J.M. Hughes et al., 2002. Medical and public health management: Hemorrhagic fever viruses as biological weapons. JAMA, 287: 2391-2405.
Direct Link - Borzenkov, V.M., A.P. Pomerantsev and I.P. Ashmarin, 1993. The additive synthesis of a regulatory peptide in vivo: The administration of a vaccinal Francisella tularensis strain that produces beta-endorphin. Biull. Eksp. Biol. Med., 116: 151-153.
PubMed - Bossi, P., A. Tegnell, A. Baka, F. Van Loock and J. Hendriks et al., 2004. Bichat guidelines for the clinical management of brucellosis and bioterrorism-related brucellosis. Euro. Surveill, 9: E15-16.
PubMed - Bowen, J.E. and C.P. Quinn, 1999. The native virulence plasmid combination affects the segregational stability of a theta-replicating shuttle vector in Bacillus anthracis var. New Hampshire. J. Applied Microbiol., 87: 270-278.
PubMed - Boyce, T.G., D.L. Swerdlow and P.M. Griffin, 1995. Escherichia coli O157: H7 and the hemolytic-uremic syndrome. N. Engl. J. Med., 333: 364-368.
PubMed - Bratu, S. and L.I. Lutwick, 2002. Active immunisation against human tickborne diseases. Expert Opin. Biol. Ther., 2: 187-195.
CrossRef - Breman, J.G. and D.A. Henderson, 2002. Diagnosis and management of smallpox. N. Engl. J. Med., 17: 1300-1308.
Direct Link - Broekhuijsen, M., P. Larsson, A. Johansson, M. Bystrom and U. Eriksson et al., 2003. Genome-wide DNA microarray analysis of Francisella tularensis strains demonstrates extensive ge-netic conservation within the species but identifies regions that are unique to the highly virulent F. tularensis subsp. tularensis. J. Clin. Microbiol., 41: 2924-2931.
CrossRef - Bronze, M.S., M. M.Huycke, L.J. Machado, G.W. Voskuhl and R.A. Greenfield, 2002. Viral agents as biological weapons and agents of bioterrorism. Am. J. Med. Sci., 323: 316-325.
PubMed - Broussard, L.A., 2001. Biological agents: Weapons of warfare and bioterrorism. Mol. Diagn, 6: 323-333.
PubMed - Butt, A.A., K.E. Aldridge and C.V. Sanders, 2004. Infections related to the ingestion of sea-food Part I: Viral and bacterial infections. Lancet Infect. Dis., 4: 201-212.
PubMed - Butzler, J.P., 2004. Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect., 10: 868-876.
CrossRefPubMedDirect Link - Calisher, C.H., 1994. Medically important arboviruses of the United States and Canada. Clin. Microbiol. Rev., 7: 89-116.
PubMed - Canter, D.A., D. Gunning, P. Rodgers L. O'connor, C. Traunero and C.J. Kempter, 2005. Remediation of B. anthracis contamination in the U.S. Department of Justice mail facility. Biosecur. Bioterror., 3: 119-127.
PubMed - Carl, M., C.W. Tibbs, M.E. Dobson, S. Paparello and G.A. Dasch, 1990. Diagnosis of acute typhus infec-tion using the polymerase chain reaction. J. Infect. Dis., 161: 791-793.
PubMed - Casadevall, A., 2002. Passive antibody administration (immediate immunity) as a specific defense against biological weapons, Emerg. Infect. Dis., 8: 833-841.
PubMed - Casadevall, A. and L. Pirofski, 2003. Exploiting the redundancy in the immune system: Vac-cines can mediate protection by eliciting unnatural immunity. J. Exp. Med., 197: 1401-1404.
PubMed - Casadevall, A. and L.A. Pirofski, 2004. The weapon potential of a microbe. Trends Microbiol., 12: 259-263.
PubMed - Celebi, G., F. Baruonu, F. Ayoglu, F. Cinar, A. Karadenizli, M.B. Ugur and S. Gedikoglu, 2006. Tularemia, a reemerging disease in northwest Turkey: Epidemiological investigation and evaluation of treatment responses. Japan J. Infect. Dis., 59: 229-234.
Direct Link - Centre of Disease Control and Prevetion, 2010. Surveillance for human west nile virus dis-ease-United States, 1999-2008. MMWR, 59: 1-17.
Direct Link - Center of Disease Control and Prevention, 2002. Yellow fever vaccine recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR, 51: 1-10.
Direct Link - Center of Disease Control and Prevention, 2002. Use of anthrax vaccine in response to ter-rorism: Supplemental recommendations of the Advisory Committee on Immunization Practices(ACIP). JAMA, 288: 2681-2682.
Direct Link - Center of Disease Control and Prevention, 2004. Chemical Emergencies, Facts About Ricin. CDC, Atlanta, GA.
Direct Link - Centers for Disease Control and Prevention, 1999. Update: Outbreak of Nipah virus, Malyasia and Singapore, 1999. MMWR, 48: 335-337.
PubMedDirect Link - Centers for Disease Control and Prevention, 2006. Human plague-four states, 2006. MMWR, 55: 940-943.
Direct Link - CDC, 2000. Laboratory-acquired human glanders Maryland, May 2000. Morb. Mortal. Wkly. Rep., 49: 532-535.
PubMedDirect Link - Chanteau, S., L. Rabarijaona, T. O`Brien, L. Rahalison and J. Hager et al., 1998. F1 antigenaemia in bubonic plague patients, a marker of gravity and efficacy of therapy. Trans. R. Soc. Trop. Med. Hyg., 92: 572-573.
PubMed - Chanteau, S., M. Ratsitorahina, L. Rahalison, B. Rasoamanana and F. Chan et al., 2000. Current epidemiology of human plague in Madagascar. Microbes Infect., 2: 25-31.
PubMedDirect Link - Christopher, G.W., M.B. Agan, T.J. Cieslak and P.E. Olson, 2005. History of US military contributions to the study of bacterial zoonoses. Military Med., 170: 39-48.
PubMed - Christopher, G.W., T.J. Cieslak, J.A. Pavlin and Jr. E.M. Eitzen, 1997. Biological warfare: A his-torical perspective. J. Am. Med. Assoc., 278: 412-417.
PubMed - Chua, K.B., S.K. Lam, K.J. Goh, P.S. Hooi and T.G. Ksiazek et al., 2001. The presence of Nipah virus in respiratory secretions and urine of patients during an outbreak of Nipah virus encephalitis in Malay-sia. J. Infect., 42: 40-43.
PubMed - Cieslak, T.J., G.W. Christopher, M.G. Kortepeter, J.R. Rowe, J.A. Pavlin, R.C. Culpepper and Jr. E.M. Eitzen, 2000. Immunization against potential biological warfare agents. Clin. Infect. Dis., 30: 843-850.
PubMed - Cohen, R.J. and E. Marshall, 2001. BIOTERRORISM: Vaccines for biodefense: A system in distress. Science, 294: 498-501.
CrossRefDirect Link - Cohen, R.J. and J.L. Stockard, 1967. Pneumonic plague in an untreated plague-vaccinated individual. JAMA, 4: 365-366.
PubMed - Coker, C., M. Majid and S. Radulovic, 2003. Development of Rickettsia prowazekii DNAvac-cine: Cloning strategies. Ann. N. Y. Acad. Sci., 990: 757-764.
PubMed - Crook, L.D. and B. Tempest, 1992. Plague: A clinical review of 27 cases. Arch. Intern. Med., 152: 1253-1256.
PubMed - Cuthbert, J.A., 2001. Hepatitis A: Old and new. Clin. Microbiol. Rev., 14: 38-58.
CrossRefDirect Link - Daniels, P., T. Ksiazek and B.T. Eaton, 2001. Laboratory diagnosis of Nipahand Hendra virus infections. Microbes Infect., 3: 289-295.
CrossRefPubMedDirect Link - DaSilva, E.J., 1999. Biological warfare, bioterrorism, bidefence and the biological and toxin weapons conention. Elect. J. Biotechnol., 2: 99-120.
Direct Link - Dennis, D.T., T.V. Inglesby and D.A. Henderson, J.G. Bartlett and M.S. Ascher et al., 2001. Tularemia as a biological weapon: Medical and public health management. JAMA, 285: 2763-2773.
Direct Link - Derbes, V.J., 1966. De mussis and the great plaque of 1348: A forgotten episode in bacterio-logical war. J. Am. Med. Assoc., 196: 59-62.
Direct Link - Dixon, T.C., M. Meselson, J. Guillemin and P.C. Hanna, 1999. Anthrax. N. Engl. J. Med., 341: 815-826.
Direct Link - Duma, R.J., D.E. Sonenshine, F.M. Bozeman, J.M. Veazey and Jr. B.L. Elisberg et al., 1981. Epidemic typhus in the United States associated with flying squirrels. JAMA, 245: 2318-2323.
PubMed - Elad, D., 2005. Risk assessment of malicious biocontamination of food. J. Food Prot., 68: 1302-1305.
Direct Link - Elvin, S.J., A.M. Bennett and R.J. Phillpotts, 2002. Role for mucosal immune responses and cell-mediated immune functions in protection from airborne challenge with Venezuelan equine encephalitis virus. J. Med. Virol., 67: 384-393.
PubMed - Emanuel, P.A., J. Dang, J.S. Gebhardt, J. Aldrich and E.A.E. Garber et al., 2000. Recombinant antibodies: A new reagent for biological agent detection. Biosens. Bioelectron., 14: 761-770.
CrossRef - Enderlin, G.L., Morales, R.F. Jacobs and J.T. Cross, 1994. Streptomycin and alternative agents for the treatment of tularaemia: Review of literature. Clin. Infect. Dis., 19: 42-47.
PubMed - Fedorova, V.A., A.V. Petrova and Z.L. Devdariani, 2005. Influence of cultivation conditions on the expression of Yersinia pestis YopE. Zh. Mikrobiol. Epidemiol. Imunobiol., 4: 3-7.
PubMed - Fenn, E.A., 2000. Biological warfare in eighteenth-century North America: Beyond jeffery amherst. J. Am. Hist., 86: 1552-1580.
Direct Link - Fenollar, F., P.E. Fournier, M.P. Carrieri, G. Habib, T. Messana and D. Raoult et al., 2001. Risks factors and prevention of Q fever endocarditis. Clin. Infect. Dis., 33: 312-316.
PubMed - Fine, D.L., B.A. Roberts, M.L. Teehee, S.J. Terpening and C.L. Kelly et al., 2007. Venezuelan equine encephalitis virus vac-cine candidate (V3526) safety, immunogenicity and efficacy in horses. Vaccine, 25: 1868-1876.
PubMed - Finnie, J.W., 2003. Pathogenesis of brain damage produced in sheep by Clostridium perfrin-gens type D epsilon toxin: A review. Aust. Vet. J., 81: 219-221.
CrossRef - Fisher-Hoch, S.P., L. Hutwanger, B. Brown and J.B. McCormick, 2000. Effective vaccine`s for Lassa fever. J. Virol., 74: 6777-6783.
PubMed - Foran, P., N. Mohammed, G. Lisk, S. Nagwaney and G.W. Lawrence et al., 2003. Evaluation of the therapeutic usefulness of botulinum neurotoxin B, C1, E, and F compared to the long-lasting type A: Basis for distinct durations of inhibition of exocytosis in central neurons. J. Biol. Chem., 278: 363-1371.
CrossRef - Franz, D.R., P.B. Jahrling, A.M. Friedlander, D.J. McClain and D.L. Hoover et al., 1997. Clinical recognition and man-agement of patients exposed to biological warfare agents. JAMA, 278: 399-411.
PubMed - Ghabriel, M.N., C. Zhu, P.L. Reilly, P.C. Blumbergs, J. Manavis and J.W. Finnie, 2000. Toxin-induced vasogenic cerebral oe-dema in a rat model. Acta Neurochir. Suppl., 76: 231-236.
PubMed - Glynn, J.R., R.B. Hornick, M.M. Levine and D.J. Bradley, 1995. Infecting dose and severity of ty-phoid: analysis of volunteer data and examination of the influence of the definition of ill-ness used. Epidemiol. Infect., 115: 23-30.
Direct Link - Gorvel, J.P. and E. Moreno, 2002. Brucella intracellular life: From invasion to intracellular replication. Vet. Microbiol., 90: 281-297.
CrossRef - Grunow, R., W. Splettstoesser, S. Mcdonald, C. Otterbein and T. O'Brien et al., 2000. Detection of Francisella tu-larensis in biological specimens using a capture enzyme-linked immunosorbent assay, an immunochromatographic handheld assay, and a PCR. Clin. Diagn. Lab. Immunol., 7: 86-90.
PubMed - Guarner, J. and S.R. Zaki, 2006. Histopathology and Immunohistochemistry in the Diagnosis of Bioterrorism Agents. J. Histochem. Cytochem., 54: 3-11.
Direct Link - Gurycova, D., 1998. First isolation of Francisella tularensis subsp. tularensis in Europe. Eur. J. Epidemiol., 14: 797-802.
CrossRef - Hallin, G.W., S.Q. Simpson, R.E. Crowell, D.S. James, F.T. Koster, G.J. Mertz and H. Levy, 1996. Cardiopulmonary manifestations of hantavirus pulmonary syndrome. Crit. Care Med., 24: 252-258.
PubMed - Hanna, P., 1998. Anthrax pathogenesis and host response. Curr. Top. Microbiol. Immunol., 225: 13-35.
PubMed - Hart, M.K., K. Caswell-Stephan, R. Bakken, R. Tammariello and W. Pratt et al., 2000. Improved mucosal protection against Venezuelan equine encephalitis virus is induced by the molecularly defined, live-attenuated V3526 vaccine candidate. Vaccine, 18: 3067-3075.
PubMed - Henderson, D.A., T.V. Inglesby, J.G. Bartlett, M.S. Ascher and E. Eitzen et al., 1999. Smallpox as a biological weapon: Medical and public health management. Working group on civilian biodefense. JAMA, 281: 2127-2137.
PubMed - Higgins, J.A., M. Cooper, L. Schroeder-Trucker, S. Black and D. Miller et al., 2002. A field investigation of Ba-cillus anthracis contamination of U.S. Department of Agriculture and other Washington, D.C., buildings during the anthrax attack of October 2001. Applied Environ. Microbiol., 69: 593-599.
PubMed - Hilleman, M.R., 2002. Overview: Cause and prevention in biowarfare and bioterrorism. Vaccine, 20: 3055-3067.
CrossRef - Hollis, D.G., R.E. Weaver, A.G. Steigerwalt, J.D. Wenger, C.W. Moss and D.J. Brenner, 1989. Francisella philomiragia comb.nov. (formerly Yersinia philomiragia) and Francisella tularensis biogroup novicida (formerly Francisella novicida) associated with human disease. J. Clin. Microbiol., 27: 1601-1608.
PubMed - Holty, J.E., D.M. Bravata, H. Liu, R.A. Olshen, K.M. McDonald and D.K. Owens, 2006. Systematic review: A century of inhalational anthrax cases from 1900 to 2005. Ann. Internal Med., 144: 270-280.
PubMedDirect Link - Hornick, R.B. and H.T. Eigelsbach, 1966. Eigelsbach, Aerogenic immunization of man with live Tulare-mia vaccine. Bacteriol. Rev., 30: 532-538.
Direct Link - Hottle, G.A., C. Nigg and J.A. Lichty, 1947. Studies on botulinum toxoids, types A and B. II. Methods for determining antigenicity in animals. J. Immunol., 55: 255-262.
Direct Link - Hubalek, Z., W. Sixl and J. Halouzka, 1998. Francisella tularensis in Dermacentor reticula-tus ticks from the Czech Republic and Austria. Wien. Klin. Wochenschr., 110: 909-910.
PubMed - Inglesby, T.V., D.T. Dennis, D.A. Henderson, J.G. Bartlett and M.S. Ascher et al., 2000. Plague as a biological weapon: Medical and public health management. Working group on civilian biodefense. JAMA, 283: 2281-2290.
PubMedDirect Link - Inglesby, T.V., T. O`Toole, D.A. Henderson, J.G. Bartlett and M.S. Ascher et al., 2002. Anthrax as a biological weapon, 2002: Updated recommendations for management. JAMA, 287: 2236-2252.
Direct Link - Jackson, P.J., M.E. Hugh-Jones, D.M. Adair, G. Green and K.K. Hill et al., 1998. PCR analysis of tissue samples from the 1979 Sverdlovsk anthrax victims: The presence of multiple Bacillus anthracis strains in different victims. Proc. Natl. Acad. Sci., 95: 1224-1229.
Direct Link - Jiang, J., J.J.Temenak, A.L. Richards, 2003. Real-time PCR duplex assay for Rickettsia prowazekii and Borrelia recurrentis. Ann. N. Y. Acad. Sci., 990: 302-310.
PubMed - Johnson, E., N. Jaax, J. White, P. Jahrling, 1995. Lethal experimental infections of rhesus monkeys by aerosolized Ebola virus. Int. J. Exp. Pathol., 76: 227-236.
PubMed - Josefson, D., 2001. US fear of bioterrorism spreads as anthrax cases increase. Br. Med. J., 323: 877-878.
PubMed - Kagawa, F.T., J.H. Wehner and V. Mohindra, 2003. Q fever as a biological weapon. Semin. Respir. Infect., 18: 183-195.
PubMed - Keim P., A. Kalif, J. Schupp, K. Hill and S.E. Travis et al., 1997. Molecular evolution and diversity in Bacillus anthracis as detected by amplified fragment length polymorphism markers. J. Bacteriol., 179: 818-824.
Direct Link - Kelly, D.J., A.L. Richards, J. Temenak, D. Strickman and G.A. Dasch, 2002. The past and present threat of rickettsial diseases to military medicine and international public health. Clin. Infect. Dis., 34: S145-169.
PubMedDirect Link - Kenyon, R.H., Jr. K.T. McKee, P.M. Zack, M.K. Rippy and A.P. Vogel et al., 1992. Aerosol infection of rhesus ma-caques with Junin virus. Inter. Virol., 33: 23-31.
PubMed - Kiel, J.L., J.E. Parker, E.A. Holwitt, R.P. McCreary and C.J. Andrews et al., 2008. Geographical distribution of genotypic and phenotypic markers among Bacillus anthracis isolates and related species by his-torical movement and horizontal transfer. Folia. Microbiol., 53: 472-478.
PubMed - Kilonzo, B.S., T.J. Mbise, D.C. Mwalimu and L. Kindamba, 2006. Observations on the en-demicity of plague in Karatu and Ngorongoro, northern Tanzania. Tanzania Health Res. Bull., 8: 1-6.
Direct Link - Knisely, R.F., 1966. Selective medium for Bacillus anthracis. J. Bacteriol., 92: 784-786.
Direct Link - Kolavic, S.A., A. Kimura, S.L. Simons, L. Slutsker, S. Barth and C.E. Haley, 1997. An outbreak of Shigella dysenteriae type 2 among laboratory workers due to intentional food contamination. JAMA, 278: 396-398.
PubMed - Krakauer, T., 2005. Chemotherapeutics targeting immune activation by staphylococcal su-perantigens. Med. Sci. Monit., 11: RA290-295.
PubMed - La Scola, B. and D. Raoult, 1997. Laboratory diagnosis of rickettsioses: Current approaches to diagnosis of old and new rickettsial diseases. J. Clin. Microbiol., 35: 2715-2727.
Direct Link - Lane, H.C., J. La Montagne and A.S. Fauci, 2001. Bioterrorism: A clear and present danger. Nat. Med., 7: 1271-1273.
CrossRef - Lee, J.S., A.G. Hadjipanayis and M.D. Parker, 2005. Viral vectors for use in the development of biodefense vaccines. Adv. Drug Deliv. Rev., 57: 1293-1314.
CrossRef - Lee, P.W., H.L. Amyx, R. Yanagihara, D.C. Gajdusek, D. Goldgaber and Jr. C.J. Gibbs, 1985. Partial characterization of Prospect Hill virus isolated frommeadowvoles in the United States. J. Infect. Dis., 152: 826-829.
PubMed - Leitenberg, M., 2001. Biological weapons in twentieth century: A review and analysis. Crit. Rev. Microbiol., 27: 267-320.
PubMed - Ligon, B.L., 2006. Plague: A review of its history and potential as a biological weapon. Semin. Pediatr. Infect. Dis., 17: 161-170.
PubMed - Lub, M., A.N. Sergeev, O.V. P`Iankov, O.G. P'iankova, V.A. Petrishchenko and L.A. Kotliarov, 1995. Certain pathogenetic characteristics of a disease in monkeys in infected with the Marburg virus by an airborne route. Vopr. Virusol., 40: 158-161.
PubMed - Lucas, M.E., J.L. Deen, L. Von Seidlein, X.Y. Wang and J. Ampuero et al. 2005. Effectiveness of mass oral cholera vaccination in Beira, Mozambique. N. Engl. J. Med., 352: 757-767.
PubMedDirect Link - Mac Kenzie, W.R., N.J. Hoxie, M.E. Proctor, M.S. Gradus and K.A. Blair et al., 1994. A massive outbreak in Milwau-kee of cryptosporidium infection transmitted through the public water supply. N. Engl. J. Med., 331: 161-167.
CrossRefDirect Link - Mackinnon, P.J. and M.R. Alderton, 2000. n investigation of the degradation of the plant toxin, ricin, by sodium hypochlorite. Toxicon, 38: 287-291.
CrossRef - Manchee, R.J., M.G. Brosler, A.J. Stagg and S.E. Hibbs, 1994. Formaldehyde solution effectively inactivates spores of Bacillus anthracis on the Scottish Island of Gruinard. Applied Environ. Microbiol., 60: 4167-4171.
PubMed - Mantis, N.J., 2005. Vaccines against the category B toxins: staphylococcal enterotoxin B, epsilon toxin and ricin. Adv. Drug Deliv. Rev., 57: 1424-1439.
PubMed - Marmion, B.P., R.A. Ormsbee, M. Kyrkou, J. Wright and D.A. Worswick et al., 1984. Vaccine prophylaxis of abattoir-associated Q fever. Lancet, 2: 1411-1414.
PubMed - Mattix, M.E., R.E. Hunt, C.L. Wilhelmsen and A.J. Johnson and W.B. Baze, 1995. Aerosolized staphylococcal enterotoxin B-induced pulmonary lesions in rhesus monkeys (Macaca mulatta). Toxicol. Pathol., 23: 262-268.
PubMed - McCoy, G.W. and C.W. Chapin, 1912. Further observations on a plaque-like disease of rodents with a preliminary note on the causative agent, Bacterium tularense. J. Infect. Dis., 10: 61-72.
Direct Link - Memish, Z.A. and H.H. Balkhy, 2004. Brucellosis and international travel. J. Travel. Med., 11: 49-55.
PubMed - Middlebrook, J.L. and D.R. Franz, 1997. Botulinum Toxins. In: Textbook of Military Medicine. Part I. Warfare, Weaponry and then Casualty: Medical Aspects of Chemical and Biological Warfare 33, Slidell, F.R., E.T. Takafuji and D.R. Franz (Eds.). Borden Institute, Walter Reed Army Medical Center, Washington, DC., pp: 643-654.
- Mobley, J.A., 1995. Biological warfare in the twentieth century: lessons from the past, challenges for the future. Mil. Med., 160: 547-553.
PubMed - Musher, D.M. and B.L. Musher, 2004. Contagious acute gastrointestinal infections. N. Engl. J. Med., 351: 2417-2427.
PubMed - Nachamkin, I., A.B. Mishu and T. Ho, 1998. Campylobacter species and Guillain-Barre syndrome. Clin. Microbiol. Rev., 11: 555-567.
Direct Link - Nagahama, M. and J. Sakurai, 1992. High-affinity binding of Clostridium perfringens epsilon-toxin to rat brain. Infect. Immun., 60: 1237-1240.
PubMed - Nagahama, M., S. Ochi and J. Sakurai, 1998. Assembly of Clostridium perfringens epsilon-toxin on MDCK cell membrane. J. Nat. Toxins, 7: 291-302.
PubMed - Nathan, N., M. Barry, M. van Herp and H. Zeller, 2003. Shortage of vaccines during a yellow fever outbreak in Guinea. Lancet, 358: 2129-2130.
PubMed - Navarro, E., M.A. Casao and J. Solera, 2004. Diagnosis of human brucellosis using PCR. Exp. Rev. Mol. Diagn., 40: 115-123.
PubMedDirect Link - Nichol, S.T., C.F. Spiropoulou, S. Morzunov, P.E. Rollin and T.G. Ksiazek, 1993. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science, 262: 914-917.
PubMed - Noah, D.L., K.D. Huebner, R.G. Darling and J.F. Waeckerle, 2002. The history and threat of biological warfare and terrorism. Emerg. Med. Clin. North. Am., 20: 255-271.
PubMed - Nurgaleeva, R.G., E.A. Tkachenko, A.G. Stepanenko, I.M. Mustafin and and S.G. Kireev et al., 1999. An epidemiological analysis of hemorrhagic fever with renal syndrome morbidity in the republic of Bashkortostan in 1997. Zh. Mikrobiol. Epidemiol. Immunobiol., 6: 45-49.
PubMed - O'Brien, J., A. Friedlander, T. Drier, J. Ezzell and S. Leppla, 1985. Effects of anthrax toxin compounds on human neutrophils. Infect. Immun., 47: 306-310.
Direct Link - Olsufjev, N.G., O.S. Emelyanova and T.N. Dunaeva, 1959. Comparative study of strains of B. tularense in the old and new world and their taxonomy. J. Hyg. Epidemiol. Microbiol. Immunol., 3: 138-149.
PubMed - Oyston, P.C., D.W. Payne, H.L. Havard, E.D. Williamson and R.W. Titball, 1998. Production of a non-toxic site-directed mutant of Clostridium perfringens epsilon-toxin which induces protective immunity in mice. Microbiology, 144: 333-341.
PubMed - Ozcurumez, M.P., N. Kischel, H. Priebe, W. Splettstosser, E.J. Finke and R. Grunow, 2004. Comparison of enzyme-linked immunosorbent assay, Western blotting, microagglutination, indirect immunofluorescence assay and flow cytometry for serological diagnosis of tularemia. Clin. Diagn. Lab. Immunol., 11: 1008-1015.
CrossRef - Pappas, G., N. Akritidis, M. Bosilkovski and E. Tsianos, 2005. Brucellosis. N. Engl. J. Med., 352: 2325-2336.
PubMedDirect Link - Pappas, G., P. Papadimitriou, N. Akritidis, L. Christou and E.V. Tsianos, 2006. The new global map of human brucellosis. Lancet Infect. Dis., 6: 91-99.
CrossRefPubMedDirect Link - Pappas, G., N. Akritidis and E.V. Tsianos, 2005. Attack scenarios with rickettsial species: Implications for response and management. Ann. N. Y. Acad. Sci., 163: 451-458.
PubMed - Parent, M.A., K.N. Berggren, LW. Kummer, L.B. Wilhelm, F.M. Szaba, I.K. Mullarky and S.T. Smiley, 2005. Cell mediated protection against pulmonary Yersinia pestis infection. Infect. Immun., 73: 7304-7310.
CrossRef - Perine, P.L., B.P. Chandler, D.K. Krause, P. McCardle and S. Awoke et al., 1992. A clinico-epidemiological study of epidemic typhus in Africa. Clin. Infect. Dis., 14: 1149-1158.
PubMed - Perry, R.D. and J.D. Fetherston, 1997. Yersinia pestis-etiologic agent of plague. Clin. Microbiol. Rev., 10: 35-66.
Direct Link - Pezard, C., E. Duflot and M. Mock, 1993. Constructing of Bacillus anthracis mutant strains producing a single toxin component. J. Gen. Microbiol., 139: 2459-2463.
PubMed - Phillpotts, R.J., L.D. Jones and S.C. Howard, 2002. Monoclonal antibody protects mice against infection and disease when given either before or up to 24 h after airborne challenge with virulent Venezuelan equine encephalitis virus. Vaccine, 20: 1497-1504.
PubMed - Phillpotts, R.J., T.L. Lescott and S.C. Jacobs, 2000. Vaccinia virus recombinants encoding the truncated structural gene region of Venezuelan equine encephalitis virus (VEEV) give solid protection against peripheral challenge but only partial protection against airborne challenge with virulent VEEV. Acta. Virol., 44: 233-239.
PubMed - Pile, J.C., J.D. Malone, E.M. Eitzen and A. Friedlander, 1998. Anthrax as a potential biological warfare agent. Arch. Intern. Med., 158: 429-434.
PubMed - Plourde, P.J., J. Embree, F. Friesen and G. Lindsay, 1992. Glandular tularemia with typhoidal features in a Manitoba child. Can. Med. Assoc. J., 146: 1953-1955.
Direct Link - Pohanka, M. and B. Skladal, 2009. Bacillus anthracis, Francisella tularensis and Y. pestis. The most important bacterial warfare agents. Rev. Folia Microbiol., 54: 263-272.
CrossRef - Pomerantsev, A.P. and N.A. Staritsyn, 1996. Behaviour of heterologous recombinant plasmid pCET in cells of Bacillus anthracis. Genetika, 32: 500-509.
PubMed - Pomerantsev, A.P., N.A. Staritsin, Y.V. Mockov and L.I. Marinin, 1997. Expression of cereolysine ab genes in Bacillus anthracis vaccine strain ensures protection against experimental haemolytic anthrax infection. Vaccine, 15: 1846-1850.
CrossRef - Prior, R.G., L. Klasson, P. Larsson, K. Williams and L. Lindler et al., 2001. Preliminary analysis and annotation of the partial genome sequence of Francisella tularensis strain Schu 4. J. Applied Microbiol., 91: 571-623.
Direct Link - Pullen, R.L. and B.M. Stuart, 1945. Tularemia: Analysis of 225 cases. JAMA, 129: 495-500.
Direct Link - Qadri, F., A.M. Svennerholm, A.S.G. Faruque and R.B. Sack, 2005. Enterotoxigenic Escherichia coli in developing countries: Epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev., 18: 465-483.
CrossRef - Queipo-Ortuno, M.I., J.D. Colmenero, G. Baeza and P. Morata, 2005. Comparison between LightCycler realtime polymerase chain reaction (PCR) assay with serum and PCR-enzyme-linked immunosorbent assay with whole blood samples for the diagnosis of human brucellosis. Clin. Infect. Dis., 40: 260-264.
PubMed - Rahalison, L., E. Vololonirina, M. Ratsitorahina and S. Chanteau, 2000. Diagnosis of bubonic plague by PCR in Madagascar under field conditions. J. Clin. Microbiol., 38: 260-263.
Direct Link - Raoult, D., J.B. Ndihokubwayo, H. Tissot-Dupont, V. Roux, B. Faugere, R. Abegbinni and R.J. Birtles, 1998. Outbreak of epidemic typhus associated with trench fever in Burundi. Lancet, 352: 353-358.
PubMed - Ray, S.M., S.D. Ahuja, P.A. Blake, M.M. Farley and M. Samuel et al., 2004. Population-based surveillance for Yersinia enterocolitica infections in FoodNet sites, 1996-1999: Higher risk of disease in infants and minority populations. Clin. Infect. Dis., 38: S181-189.
PubMed - Riedels, S., 2005. Plague: From natural disease to bioterrorism. Proc. Bayl. Univ. Med. Center, 18: 116-124.
CrossRefPubMedDirect Link - Robertson, D.L. and S.H. Leppla, 1986. Molecular cloning and expression in Escherichia coli of the lethal factor gene of Bacillus anthracis. Genetic, 44: 71-78.
PubMed - Roffey, R., A. Tegnell and F. Elgh, 2002. Biological warfare in a historical perspective. Clin. Microbiol. Infect., 8: 450-454.
CrossRef - Rusnak, J.M., M. Kortepeter, R. Ulrich, M. Poli and E. Boudreau, 2004. Laboratory exposures to staphylococcal enterotoxin B. Emerg. Infect. Dis., 10: 1544-1549.
PubMed - Ryan, K.J. and C.G. Ray, 2004. Sherris Medical Microbiology. 4th Edn., McGraw Hill, New York, USA, "pp: 1-997".
Direct Link - Saslaw, S., H.T. Eigelsbach, J.A. Prior, J.E. Wilson and S. Carhardt, 1961. Tularemia vaccine study. II. Respiratory challenge. Arch. Inter. Med., 107: 702-714.
PubMed - Schep, L.J., W.A, Temple, G.A. Butt and M.D. Beasley, 2009. Ricin as a weapon of mass terror: Separating fact from fiction. Environ. Int., 35: 1267-1271.
CrossRef - Schmitt, P., W. Splettstosser, M.P. Ozcurumez, E.J. Finke and R. Grunow, 2005. A novel screening ELISA and a confirmatory Western blot useful for diagnosis and epidemiological studies of tularemia. Epidemiol. Infect., 133: 759-766.
PubMed - Schoepp, R.J., J.F. Smith and M.D. Parker, 2002. Recombinant chimeric western and eastern equine encephalitis viruses as potential vaccine candidates. Virology, 302: 299-309.
PubMed - Scola, B.L., 2002. Current laboratory diagnosis of Q fever. Semin. Pediatr. Infect. Dis., 13: 257-262.
Direct Link - Sellek, R., O. Jimenez, C. Aizpurua, B. Fernandez-Frutos and P. De Leon et al., 2008. Recovery of Francisella tularensis from soil samples by filtration and detection by real-time PCR and ELISA. J. Environ. Monit., 10: 362-369.
PubMed - Sjostedt, A., A. Tarnvik and G. Sandstrom, 1996. Francisella tularensis: Hostparasite inter-action. FEMS Immunol. Med. Microbiol., 13: 181-184.
Direct Link - Small, P.M. and P.I. Fujiwara, 2001. Management of tuberculosis in the United States. N. Engl. J. Med., 345: 189-210.
Direct Link - Smith, H.V. and G.D. Corcoran, 2004. New drugs and treatment for cryptosporidiosis. Curr. Opin. Infect. Dis., 17: 557-564.
PubMed - Smither, S.J., J. Hill, B.L. Van Baar, A.G. Hulstb, A.L. de Jongb and R.W. Titball, 2007. Identification of outer membrane proteins of Yersinia pestis through biotinylation. J. Microbiol. Meth., 68: 26-31.
CrossRef - Stephenson, E.H., E.W. Larson and J.W. Dominik, 1984. Effect of environmental factors on aerosol-induced Lassa virus infection. J. Med. Virol., 14: 295-303.
PubMed - Stienlauf, S., A. Eisenkraft, E. Robenshtok and A. Hourvitz, 2002. Viral encephalitis caused by biowarfare agents. Harefuah, 141: 121-121.
PubMed - Sullivan, N.J., T.W. Geisbert, J.B. Geisbert, L. Xu and Z.Y. Yang et al., 2003. Accelerated vaccination for Ebola virushaemorrhagic fever in non-human primates. Nature, 424: 681-684.
PubMed - Tamilselvam, B. and S. Daefler, 2008. Francisella targets cholesterol-rich host cell membrane domains for entry into macrophages. J. Immunol., 180: 8262-8271.
PubMedDirect Link - Tanabe, M., H.S. Atkins, D.N. Harland, S.J. Elvin and A.J. Stagg et al., 2006. The ABC transporter protein OppA provides protection against experimental Yersinia pestis infection. Infect. Immun., 74: 3687-3691.
Direct Link - Tims, T.B. and D.V. Lim, 2004. Rapid detection of Bacillus anthracis spores directly from powders with an evanescent wave fiber-optic biosensor. J. Microbiol. Meth., 59: 127-130.
CrossRef - Titball, R.W. and E.D. Williamson, 2001. Vaccination against bubonic and pneumonic plague. Vaccine, 19: 4175-4184.
PubMed - Titball, R.W., J. Hill, D.G. Lawton and K.A. Brown, 2003. Yersinia pestis and plague. Biochem. Soc. Trans., 31: 104-107.
Direct Link - Titball, R.W., P.C.B. Turnbull and R.A. Huston, 1991. The monitoring and detection of Bacillus anthracis in the environment. J. Applied Bacteriol. Symp., 20: 9-18.
PubMed - Toro, J., J.D. Vega, A.S. Khan, J.N. Mills and P. Padula et al., 1998. An outbreak of hantavirus pulmonary syndrome. Chile, 1997. Emerg. Infect. Dis., 4: 687-694.
Direct Link - Torok, T.J., R.V. Tauxe, R.P. Wise, J.R. Livengood and R. Sokolow et al., 1997. A large community outbreak of salmonellosiscaused by intentional contamination of restaurant salad bars. JAMA, 278: 389-395.
Direct Link - Tucker, J.B., 1999. Historical trends related to bioterrorism: An empirical analysis. Emerg. Infect. Dis., 5: 498-504.
Direct Link - Turnbull, P.C.B., 1999. Definitive identification of Bacillus anthracis: A review. J. Applied Microbiol., 2: 237-240.
PubMed - Waag, D.M., M.J. England and M.L. Pitt, 1997. Comparative efficacy of a Coxiella bur-netii chloroform: Methanol residue (CMR) vaccine and a licensed cellular vaccine (Q-Vax) in rodents challenged by aerosol. Vaccine, 15: 1779-1783.
PubMed - Waag, D.M., M. J. England, C.R. Bolt and J.C. Williams, 2008. Low-dose priming before vacci-nation with the phase I chloroform-methanol residue vaccine against Q fever enhances humoral and cellular Immune responses to Coxiella burnetii. Clin. Vac. Immunol., 15: 1505-1512.
CrossRef - Wang, T., Z. Qi, B. Wu, Z. Zhu and Y. Yang et al., 2008. A new purification strategy for fraction 1 capsular antigen and its efficacy against Yersinia pestis virulent strain challenge. Protein Expr. Purif, 61: 7-12.
PubMed - Weber, D.J. and W.A. Rutala, 2001. Risks and prevention of transmission of rare zoonotic diseases. Clin. Infect. Dis., 32: 446-446.
PubMed - Wein, L.M. and Y. Liu, 2005. Analyzing a bioterror attack on the food supply: The case of botulinum toxin in milk. Proc. Natl. Acad. Sci., 102: 9984-9999.
Direct Link - Whitehouse, C.A. and H.E. Hottel, 2006. Comparison of five commercial DNA extraction kits for the recovery of Francisella tularensis DNA from spiked soil samples. Mol. Cell. Probes, 21: 92-96.
PubMed - Young, E.J., 1995. An overview of human brucellosis. Clin. Infect. Dis., 21: 283-289.
CrossRefPubMedDirect Link - Raoult, D., T. Marrie and J. Mege, 2005. Natural history and pathophysiology of Q fever. Lancet Infect. Dis., 5: 219-226.
PubMedDirect Link