
T.O. Femi-Ola
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
B.M. Olowe
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 140-146







ABSTRACT
Amylase producer Bacillus subtilis BS5 was obtained from the hindgut of wood eating termites Amitermis evuncifer Silvestri. Amylase enzyme produced by this strain when grown on basal liquid medium with cocoyam starch as substrate was characterized. The enzyme was purified through ion-exchange chromatography and gel-filtration. The purified enzyme had a molecular weight of 63 kDa and was active optimally at pH 6.0 and 50°C. The apparent Km value and Vmax of the enzyme during hydrolysis of soluble starch were 16.67 mg mL-1 and 3.8 mg/min/mL-1, respectively. The activities of the amylase were stimulated by Ca2+, Na+ and Mg2+ but inhibited by EDTA and HgCl2.
PDF Abstract XML References Citation
Received: November 18, 2009;
Accepted: July 31, 2010;
Published: September 30, 2010
How to cite this article
T.O. Femi-Ola and B.M. Olowe, 2011. Characterization of Alpha Amylase from Bacillus subtilis BS5 Isolated from Amitermes evuncifer Silvestri. Research Journal of Microbiology, 6: 140-146.
URL: https://scialert.net/abstract/?doi=jm.2011.140.146
URL: https://scialert.net/abstract/?doi=jm.2011.140.146
INTRODUCTION
Starch is an abundant carbon source in nature. α-amylase (1, 4 α D-glucanohydrolase; EC 3.2.1.1) hydrolyses α-1, 4-glucosidic linkage in starch and related molecules. It is one of several enzymes involved in starch degradation. Amylases constitute one of the most important groups of industrial enzymes being extensively used in food, textiles, paper, brewing and distilling industries. Most of the available amylases produced commercially are of microbial origin (Pandey et al., 2000). The enzyme is widespread among aerobes and anaerobes. Gram positive bacteria, particularly the genera Bacillus and Clostridia are prolific producers of amylases (Haddaoui et al., 1995; Shih and Labbe, 1995; Obineme et al., 2003; Swain and Ray, 2007). Several species of Bacillus produces a wide range of extra cellular enzymes of which amylases and proteases are of significant industrial importance. These enzymes are used in the textile and paper industries, in starch liquefaction, as a food additive and in sugar production (Pandey et al., 2000). The amylase activity has been shown to be influenced by temperature, pH and presence of some chemicals (Swain and Ray, 2007). A Bacillus subtilis strain BS5 was isolated from the hindgut of wood-eating termite, Amitermes evuncifer Silvestri (Femi-Ola and Aderibigbe, 2006; Femi-Ola et al., 2007). It was selected for further study because of its high amylolytic and cellulolytic activity which was comparable with a reference strain B. subtilis NCIB 3610. This study reports a partial purification and characterization of alpha amylase produced by the B. subtilis strain BS5.
MATERIALS AND METHODS
Organisms and culture conditions: The B. subtilis strain BS5 used in this study was isolated from the hindgut of a wood-eating termite, Amitermes evuncifer (Femi-Ola et al., 2007). This experiment was conducted between March 2008 and June 2009. The organism was grown in a basal medium containing (g L-1): K2HPO4, 1.5; KH2PO4, 0.5; MgSO4, 0.05; NaCl, 1.5; (NH4)2SO4, 1.0; CaCl2.2H2O, 0.02; FeSO4.7H2O 0.02; yeast extract, 0.5 and 1% cocoyam starch. The inocula for the experiments were prepared by growing the organism in nutrient broth (NB, Oxoid) at 35°C for 18 h on a rotary shaker (Gallenkamp) and shaked at 120 rpm. Sterilized medium (500 mL) in 1000 mL conical flasks was inoculated with 10 mL of inocula (8.6x106 cells mL-1). The flask was incubated at 35°C on a rotary shaker (120 rpm) for 48 h and then centrifuged at 5000 rpm for 20 min in cold to remove bacterial cells. The supernatant obtained was used as the crude extract for further studies.
Amylase assay: Enzyme assay was estimated by the dinitrosalicyclic acid (DNSA) method of Miller (1972). The reaction mixtures consist of 0.5 mL of substrate solution (1% soluble starch in 0.05 M phosphate buffer, pH 6.9) and 0.5 mL of the cell free extract. The reaction mixture was incubated for 3 min at 30°C. The reaction was terminated by the addition of 1 mL of dinitrosalicyclic (DNSA) reagent. The mixture was heated at 100°C for 5 min and cooled. The optical density was read at 540 nm in a spectrophotometer (Jenway, 6305). One unit of alpha amylase activity (U) was defined as the amount of enzyme that liberated reducing sugar equivalent to one micromole of D-glucose from starch under the assay condition.
Purification and characterization of alpha amylase: All chromatography procedures were carried out at 4°C except where stated otherwise.
Ion exchange chromatography: DEAE-Sephadex A-50 (Pharmacia, Uppsala,Sweden) was packed into a column (2.5x40 cm) and equilibrated with 0.015 M Na2HPO4 buffer (pH 6.9). Twenty milliliters of the crude enzyme was applied to the cation exchanger column at a flow rate of 20 mL h-1. The fractions containing alpha amylase activity unbound to the gel were pooled and bound proteins were eluted with by a 0-1 M NaCl gradient in the same buffer at a flow rate of 20 mL h-1. The pooled fractions were concentrated by dialysis against glycerol solution at 4°C overnight.
Gel filtration chromatography (using Sephadex G-150): Active fractions from DEAE Sephadex A-50 (Pharmacia) column was applied to a Sephadex G-150 (Pharmacia) column (1.5x75) which had been previously equilibrated with 0.015 M Na2HPO4 buffer, pH 4.7. The column was eluted with the same buffer at a flow rate of 25 mL h-1. A fraction of 5.0 mL were collected at interval of 30 min and the absorbance at 280 nm was read using spectrophotometer (Jenway, 6305). For determination of molecular weight by gel filtration the standards used were: lysozyme 14 kDa; lactaglobulin 20 kDa; bovine trypsinogen, 24 kDa; pepsin, 34 kDa; egg albumin, 45 kDa and bovine serum albumin, 66 kDa (Sigma, UK).
Effect of temperature on alpha amylase activity and stability: Alpha amylase activity was assayed by incubating the enzyme reaction mixture at different temperatures ranged from 20°C to 80°C for 3 min. The thermal stability at 70 and 80°C was also determined. Samples were taken at 5 min intervals and analyzed for amylolytic activity.
Effect of pH on amylase activity: Substrates (1% soluble starch) having pH ranging from 3.0 to 8.0 were prepared using 0.05 M of different buffer system (Glycine-HCl, pH 3; acetate buffer, pH 4 and 5; phosphate buffer pH 6 and 7; Tris- HCl, pH 8). Enzyme activity was determined at 35°C.
Effect of substrate concentration on alpha amylase activity: The effect of substrate concentration [S] on the rate of enzyme action was studied using [S] concentration ranged from 2.0 to 10.0 mg mL-1. The Lineweaver-Burke plot was made. Both the Vmax and Km of the enzyme were calculated.
Effect of heavy metals on enzyme activity: A stock solution of 0.01 M of HgCl2 and EDTA were prepared. The substrate/chemical mixture was incubated at room temperature for 5 min before it was used in enzyme reaction assay.
Effect of cations: A stock solution of 0.01 M of each salt was prepared. The effects of some salts/cations (NaCl, CaCl2, CuSO4 and MgCl) on enzyme activity was also determined. The substrate/salts mixture was also incubated before it was used for enzyme assay.
RESULTS
Fractionation of the enzyme concentrate on DEAE Sephadex A-50 produced a double peak (Fig. 1). From the elution profile on Sephadex G-150, it was observed that the amylase was eluted as a well resolved single peak of amylase activity (Fig. 2). These purification procedures yielded a pure amylase with specific activity of 0.99 U mg-1 proteins and a purification of approximately four fold with 74.62% yield of proteins (Table 1). The molecular weight of the alpha amylase produced was estimated to be 63 kDa. The effect of temperature on the activity of the purified amylase is shown in Fig. 3. Maximum activity was observed at 50°C. The pH value for maximal activity was 6.0 (Fig. 4). The enzyme was partially stable at 70°C as it retained 56.6% of its activity after when heated for 10 min (Fig. 5). At temperature above 70°C, the enzyme was quickly denatured, completely lost its activity after 10 min.
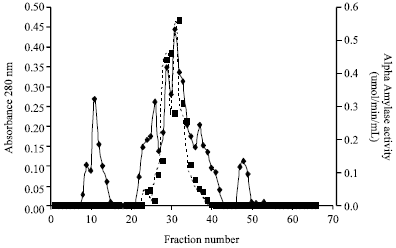 | |
| Fig. 1: | Elution profile of α-amylase produced by Bacillus subtilis from ion exchange column (2.5x40 cm) |
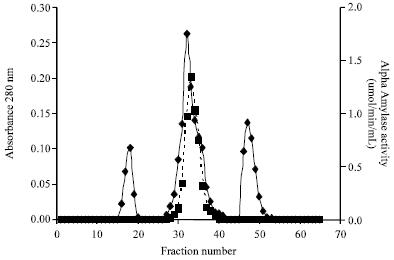 | |
| Fig. 2: | Elution profile of α-amylase produced by Bacillus subtilis from Gel filtration column (1.5x75 cm) |
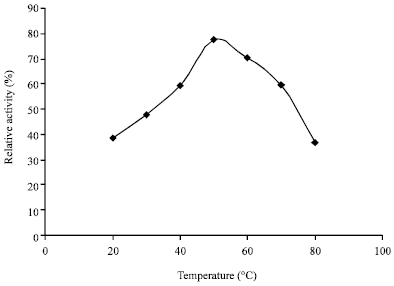 | |
| Fig. 3: | Effect of temperature on the activity of partially purified alpha amylase of Bacillus subtilis BS5 |
| Table 1: | Purification of extracellular amylase of B. subtilis BS5 |
 | |
A Lineweaver-Burke plot of the purified Alpha amylase activity of B. subtilis (Fig. 6) indicates that this enzyme has apparent Km and Vmax values for the hydrolysis of soluble starch of 16.67 mg mL-1 and 3.82 U, respectively. The activity of alpha amylase was stimulated by Ca2+, Na+ and Mg2+ while heavy metals inhibited the activity (Table 2).
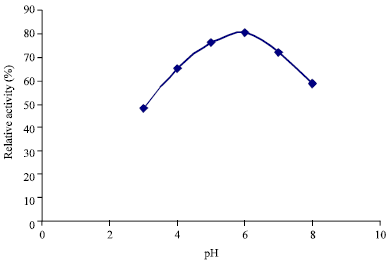 | |
| Fig. 4: | Effect of pH on alpha amylase production by B. subtilis BS5 |
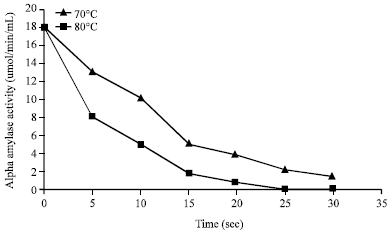 | |
| Fig. 5: | Effect of heating at 70 and 80°C on alpha amylase activity of B. subtilis BS5 |
 | |
| Fig. 6: | Lineweaver-Burke plot for the hydrolysis of starch by partially purified α-amylase from Bacillus subtilis BS5 |
| Table 2: | Effect of salts on the activity of alpha amylase of B. subtilis |
 | |
DISCUSSION
The current result of this investigation showed that temperature considerably affected the activity of the amylase enzyme. The optimum temperature for the α-amylase activity was 50°C. Other investigators working on alpha amylase have reported various temperature optima in this range. Obineme et al. (2003) and Shelby (1993) reported an optimal activity of α-amylase at 50°C for Aspergillus oryzae and Streptococcus bovis JBI respectively. The α-amylase of B. subtilis BS5 still retained 56.6 and 30.46% of its activity when heated for 10 min at 70 and 80°C, respectively. Denaturation of enzyme proteins at temperature higher than 70°C has been reported (Aderibigbe, 1998). The optimum pH for the α-amylase activity in this study was 6.0. This optimal α-amylase activity pH value was within the range of values reported for most starch degrading bacterial strain (Gupta et al., 2003). Mishra and Behera (2008) had also reported a pH of 6.0 and 7.0 for normal growth and enzyme activity for alpha amylase of Bacillus strains isolated from soil receiving kitchen waste. Similar values for bacterial amylases production by Streptococcus bovis JBI, new Clostridium isolate and Clostridium acetobutylicum SAI were reported (Chojecki and Blascheck, 1986; Madi et al., 1987; Shelby, 1993). It was also noticed that amylolytic activity increased with substrate concentration. Among the physiological parameters, optimum temperature, substrate concentration and the pH of the growth medium plays an important role in production and activity of microbial enzymes.
This study also revealed that amylolytic activity was affected by the concentration of cations at 0.04 M. Ca2+ and Na+ had stimulatory effect on the activity of B. subtilis amylase, while heavy metals, Hg2+ and EDTA inhibited the activity. The inactivation of the enzyme by heavy metals poisoning are well documented (Kim et al., 2005; Pandey et al., 2000). In their reports, it was suggested that EDTA acted by chelating Ca2+ and once the Ca2+ content of the enzyme was completely removed by EDTA, there followed a quick loss in activity of the enzyme. Heavy metals are known to react with protein sulphydryl groups thus converting them to mercaptides. The inhibition of the enzyme activity by EDTA suggests that the enzyme contains metallic ions which are removed by the chelating agent, forming an active complex with EDTA (Dixon and Webb, 1971).
The results presented in this study indicate that the α-amylase produced by B. subtilis BS5 possesses properties of an industrial enzyme; productivity and thermo stability.
REFERENCES
- Chojecki, A. and H.P. Blaschek, 1986. Effect of carbohydrate source on alpha-amylase and glucoamylase formation by Clostridium acetobutylicum SA-1. J. Ind. Microbiol., 1: 63-67.
CrossRefDirect Link - Femi-Ola, T.O., E.Y. Aderibigbe and L. Awoyemi, 2007. Microbiology of the hindgut and survival of Amitermes evuncifer (Silvestri) on some Nigerian woods. Res. J. Microbiol., 2: 910-917.
CrossRefDirect Link - Gupta, R., P. Gigras, H. Mohapatra, V.K. Goswami and B. Chauhan, 2003. Microbial α-amylases: A biotechnological perspective. Process Biochem., 38: 1599-1616.
CrossRefDirect Link - Haddaoui, E., M.F. Petit-Glatron and R. Chambert, 1995. Characterization of a new cell bound alpha amylase in Bacillus subtilis 168 Marburg that is only immunologically related to the exocellular alpha amylase. J. Bacteriol., 177: 5148-5150.
PubMed - Kim, J.Y., S.H. Hur and J.H. Hong, 2005. Purification and characterization of an alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-81. Biotechnol. Lett., 27: 313-316.
CrossRef - Madi, E., G. Antranikian, K. Ohmiya and K.G. Gottschu, 1987. Thermostable alpha amylolytic enzyme from a new Clostridium isolate. Applied Environ. Microbiol., 53: 1661-1667.
Direct Link - Mishra, S. and N. Behera, 2008. Amylase activity of a starch degrading bacteria isolated from soil receiving kitchen wastes. Afr. J. Biotechnol., 7: 3326-3331.
Direct Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Freer, S.N., 1993. Purification and characterization of the extracellular alpha amylase from Streptococcus bovis JBI. Applied Environ. Microbiol., 59: 1398-1402.
PubMedDirect Link - Shih, N. and R.G. Labbe, 1995. Purification and characterization of an extracellular amylase from Clostridium perfringes Type A. Applied Environ. Microbiol., 61: 1776-1779.
Direct Link - Swain, M.R. and R.C. Ray, 2007. Alpha amylase production by B. subtilis CM3 in solid state fermentation using cassava fibrous residue. J. Basic Microbiol., 47: 417-425.
CrossRef - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link
tanya tejeswita Reply
very clean results