
K. Apun
Department of Molecular Biology,
K.L. Kho
Department of Molecular Biology,
Y.L. Chong
Department of Zoology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia
F.H. Hashimatul
Department of Molecular Biology,
M.T. Abdullah
Department of Zoology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia
M.A. Rahman
Department of Zoology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia
M.B. Lesley
Department of Molecular Biology,
L. Samuel
Department of Molecular Biology,
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 132-139







ABSTRACT
This study was carried out to assess the occurrence of Escherichia coli and to detect the pathogenic strain Escherichia coli O157:H7 in birds, bats and rodents from disturbed habitats comprising of two urban forests and an oil palm plantation habitats located along the Rejang Basin, Sibu in the state of Sarawak, using both standard microbiological and molecular techniques. A total of 105 bird hosts, 44 of rodent hosts and 84 bat hosts represented 48 species of birds, one species of rodent and ten species of bats were screened for the presence of Escherichia coli. The representative isolates were cultured on a highly selective agar, Cefaxime-tellurite sorbitol MacConkey agar for the detection of Escherichia coli O157:H7. From the microbiological analysis, the overall occurrences of Escherichia coli in the hosts were 43% in rodents, 18% in birds and 11% in bats. The isolates were tested for the presence of pathogenic Escherichia coli O157:H7 strain by multiplex PCR method targeting the slt-I, slt-II, rfbE and fliCH7 genes. The slt-I, slt-II, rfbE genes were not detected in any of the E. coli isolates. This study indicated that bats, birds, or rodents from these habitats in Sarawak did not serve as an important reservoir of Escherichia coli O157:H7 and thus were of no risk in the epidemiologic cycle of emerging enteric bacterial zoonoses in the state of Sarawak, Malaysia.
PDF Abstract XML References Citation
Received: June 22, 2010;
Accepted: September 04, 2010;
Published: September 30, 2010
How to cite this article
K. Apun, K.L. Kho, Y.L. Chong, F.H. Hashimatul, M.T. Abdullah, M.A. Rahman, M.B. Lesley and L. Samuel, 2011. Detection of Escherichia coli O157:H7 in Wildlife from Disturbed Habitats in Sarawak, Malaysia. Research Journal of Microbiology, 6: 132-139.
URL: https://scialert.net/abstract/?doi=jm.2011.132.139
URL: https://scialert.net/abstract/?doi=jm.2011.132.139
INTRODUCTION
Wildlife is well-known to be involved in most of the zoonotic diseases (Kruse et al., 2004). The zoonotic agents, typically various bacteria, carried by wildlife, serves as the major reservoirs for microbial transmission to both human and domestic animals. Outbreaks of zoonoses have recently received increased attention worldwide due to their major impact on human health, agriculture production, wildlife-based economies and wildlife conservation (Chomel et al., 2007). Wild animals are thought to be the source of more than 70% of all emerging infections (Kuiken et al., 2005). According to World Health Organization (2008), millions of people are infected by zoonotic diseases every year.
Enterobacteriaceae are among the best-characterized group of microflora in the gastro-intestinal tract of wildlife. The Enterobacteriaceae and Escherichia coli (E. coli) are among the most common members and harmless commensal strains found in the gastro-intestinal tract of humans and animals (Boyd, 1995). However, certain E. coli strain carried the toxin gene, for instance, E. coli serotype O157:H7 which cause haemorrhagic colitis and haemolytic uremic syndrome (Griffin and Mead, 1998). E. coli O157:H7 was first recognized as a human pathogen since 1982 with the outbreaks of hemorrhagic colitis occurred by consumption of undercooked meat in Oregon and Michigan (Riley et al., 1983). E. coli O157:H7 belongs to Shiga toxin-producing E. coli (STEC) represent the only pathogenic group of the E. coli that has a definite zoonotic origin with cattle being identified as a major reservoir for this organism (Caprioli et al., 2005). However, it has been reported to be isolated from wild birds (Wallace et al., 1997; Foster et al., 2006) and rodents (Nielsen et al., 2004) which implicated wild animals also served as potentials vectors for the dissemination of E. coli O157:H7.
The surveillance of the epidemiology of pathogenic enterobacteria in wildlife is important due to the increasing activity of translocation of wild animals which increase the possibility of disseminating the pathogenic bacteria. In addition, the study of diseases spreading and epidemiology in human population are crucial for the development of prevention steps and control. The surveillance of pathogenic enterobacteria in different ecological niche is important as the development affects the diversity and distribution of microorganisms in the animal hosts (Gordan and Fitzgibbon, 1999). When the habitat is disturbed, this will influenced the animal hosts’ community structure and living behavior which indirectly affects the enteric community structure (Gordon and Cowling, 2003). Therefore, this study was conducted to compare the occurrence of E. coli and the detection of E. coli O157:H7 in wildlife from the two urban forests and an oil palm plantation sites located along the Rejang Basin, Sibu, in the state of Sarawak, Malaysia which is one of the world’s richest and most diverse ecosystem. The two urban forests and oil palm plantation represented disturbed habitats.
MATERIALS AND METHODS
Study habitats and specimens collection: Bat, rodent and bird specimens were collected from a forest park, a recreational park and an oil palm plantation in Sibu, Sarawak, Malaysia during June 2008. All three sampling habitats represented disturbed habitats located about ten km from Sibu Town. Birds and fruit bats were captured using standard mist nets; while insect bats were captured using harp traps and small rodents were captured using cage traps. Identification of mammals (bats and rodents) and birds was done following Payne et al. (1985) and Francis (2005), respectively.
Bacteria samples collection: Anal swabs of small mammals and cloacal swabs from avian specimens were collected using wet sterilized cotton buds and immediately stored in 900 μL Phosphate Buffer Saline (PBS). Samples were kept at 4°C until processed, usually within the same day. All of the samples were cultured directly on MacConkey agar (Oxoid, England), a non-selective media for isolation of Enterobacteriaceae and incubated aerobically at 37°C for 24 h, on the field. Isolates tentatively identified as E. coli were further plated on EMB agar (Oxoid, England) after taken back to the laboratory. Plates were examined for the characteristic appearance of E. coli (metallic green sheen) and colonies that exhibited these characteristics were stored in nutrient agar (Oxoid, England).
Bacteria identification: All isolates were tested through gram-staining and a series of standard biochemical tests. The biochemical tests included Kligler Iron Agar, IMVIC test and motility test.
E. coli reference cultures ATCC25922 was included as a positive control. The bacteria were identified and the results were compared to the Bergey’s Manual of Systematic Bacteriology (Krieg et al., 1984). The bacteria were further confirmed by using commercial identification kits, API 20E system (BioMerieux, France). Representative isolates were cultured on a highly selective agar Cefaxime-tellurite sorbitol MacConkey Agar (Oxoid, England) for the detection of E. coli O157:H7. Colonies which were non-sorbitol fermenter were further confirmed with E. coli O157:H7 latex agglutination kit (Oxoid, England).
DNA extraction: Bacterial total DNA of the E. coli isolate was extracted by using boiling extraction method as described by Gallian (2003) with minor modification. For the extraction of bacterial genomic DNA, E. coli isolates was cultured in 5 mL Luria-Bertani broth (BBL, United States) for overnight at 37°C with agitation at 150 rpm. Overnight broth suspensions (1 mL) were then centrifuged at 13,000 rpm for 5 min to collect the pellets. After that, the pellets were re-suspended in 100 μL of distilled water and boiled for 20 min. The lysed cells were then cooled in ice for another 20 min and centrifuged at 13,000 rpm for 3 min. The supernatant containing DNA was used as template in the multiplex PCR assay.
Multiplex PCR assay for the detection of E. coli O157:H7: Multiplex PCR assay for genes encoded Shiga toxin I and II, antigen O157 and antigen H7 were performed in a single tube reaction. Four set of primers synthesized commercially by FIRST BASE laboratory Sdn. Bhd. were used in this study. SLT-I, SLT-II primer sets were targeted for the Shiga toxin producing genes (slt-I and slt-II) and Rfb and FLICh7 primer sets were targeted for the gene involved in biosynthesis of O157 antigen and H7 antigen, respectively. E. coli O157:H7 reference strains EDL933 was included as a positive control. The primers used for the amplification of the Shiga-toxin genes were described by Meng et al. (1997) and the detection of rfbE gene and fliCh7 gene were as previously reported by Gannon et al. (1997).
The condition for the multiplex PCR assay was performed as described by Hu et al. (1999). Each PCR was performed in a total reaction volume of 25 μL containing 1 X PCR buffer, 2.5 mM Magnesium Chloride (MgCl2) (Promega Corp, USA), 0.2 mM dNTP mix (Promega Corp, USA), 0.2 μM of primers SltI-F/ SltI-R, SltII-F/ SltII-R, Rfb-F/ Rfb-R, FLICh7-F/ FLICh7-R , 1 Unit GoTaq@ Flexi DNA polymerase (Promega Corp, USA) and 10 μL of DNA template. PCR amplification was performed using iCyclerTM Thermal Cycler (Bio-Rad, USA) as follows: Initial denaturation at 94°C for 5 min, 35 cycles of denaturation at 94°C for 30 sec; annealing at 59°C for 1 min and extension at 72°C for 1 min followed by final extension at 72°C for 7 min.
Gel electrophoresis: A 10 μL aliquot of each amplification product was analyzed using electrophoresis on 2% agarose gels cast and ran in 1X TAE buffer. A 100 bp marker (Vivantis, Malaysia) was included in the gel. Gel was stained with ethidium bromide (0.5 μg mL-1) and visualized using transmitted ultraviolet illumination and photographed using gel documentation system (AlphaDigiDoc RT).
RESULTS
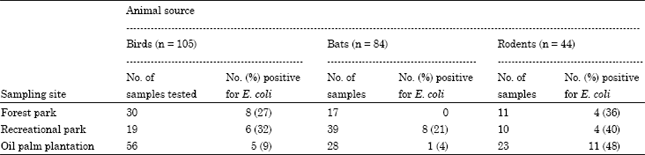
A total of 105 birds (48 species), 84 bats (10 species) and 44 rodents (one species) were captured during the fieldtrips. The results of the 105 bird hosts revealed that 19 cloacal swab samples were positive for E. coli. Nine anal swab samples from 84 bat hosts were positive for E. coli and 19 anal swab samples out of 44 rodent hosts harbored E. coli.
| Table 1: | Occurrence of E. coli in selected wildlife sampled from disturbed habitats of Sibu, Sarawak |
 | |
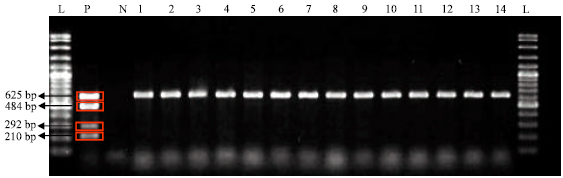 | |
| Fig. 1: | Amplicon obtained by multiplex PCR for the 14 E. coli isolates isolated from Sibu with flich7 gene with expected size of 625bp fragmented by 2% agarose gel electrophoresis. Lane L: 100bp ladder, Lane P: E. coli O157:H7 reference strains EDL933 as Positive control, Lane N: Negative control, Lane 1: Isolate (C0813) from bat sample in recreation park, Lane 2: Isolate (TK153581) from bat sample in recreation park. Lane 3: Isolate (TK153583) from bat sample in recreation park, Lane 4: Isolate (BA025) from bird sample in recreation park, Lane 5: Isolate (A2110) from bird sample in forest park, Lane 6: Isolate (TK156080) from rodent sample in oil palm plantation, Lane 7: Isolate (PL007) from rodent sample in oil palm plantation, Lane 8 : Isolate (PL010) from rodent sample in oil palm plantation, Lane 9: Isolate (PL011) from rodent sample in oil palm plantation, Lane 10: Isolate (PL0031) from rodent sample in oil palm plantation, Lane 11: Isolate (PL033) from rodent sample in oil palm plantation, Lane 12: Isolate (PL0035) from rodent sample in oil palm plantation, Lane 13: Isolate (PL0036) from rodent sample in oil palm plantation and Lane 14: Isolate (D0113) from bird sample in oil palm plantation |
The 106 isolates of E. coli represented 32, 40 and 34 isolates from the forest park, recreational park and oil palm plantation, respectively.
The occurrence of E. coli in the wildlife samples was illustrated in Table 1 the overall occurrence of E. coli in the wildlife isolated from the three habitats in Sibu was 11, 18 and 43% for bats, birds and rodents, respectively. The occurrence of E. coli was the highest in rodents from the three sampling habitats and the least in bats.
All of the positive E. coli isolates were then further cultured on Cefaxime-tellurite Sorbitol MacConkey Agar (CT-SMAC) for the detection of E. coli O157:H7. Shiga toxin-producing strains of E. coli did not ferment sorbitol and appeared as colourless colonies. There were twelve isolates that produced colourless colony (non-sorbitol fermenter) on CT-SMAC. All the isolates did not show any agglutination for E. coli O157:H7 latex agglutination kit, indicating negative reaction. The remaining E. coli isolates (sorbitol fermenter) showed pink colonies on CT-SMAC.
To confirm if any of the positive (sorbitol fermenter) E. coli isolates belong to E. coli O157:H7, 47 representative E. coli isolates from the three habitats were subjected to multiplex PCR. From 47 isolates tested, none of the isolates generated slt-I, slt-II and rfbE gene with the expected band size of 210, 292 and 484 bp, respectively. However, gene encoded for antigen H7 of expected size of 625bp was detected in 14 E. coli isolates recovered from three bats hosts, three bird hosts and eight rodent hosts (Fig. 1). This result indicated that none of the E. coli isolates were E. coli O157:H7 or STEC.
DISCUSSION
Present results showed that among the animal hosts studied, the occurrence of E. coli was always higher in rodents, followed by birds and lastly bats, in all the three sites examined. The occurrence of E. coli in birds (18%) and rodents (43%) was in general agreement with a previous study in Australia, where 23 and 43% of E. coli were found in bird and rodents, respectively (Gordon and Cowling, 2003). The occurrence of E. coli in bats (11%) was generally in accordance with previous study by Adesiyun et al. (2009), where 13% of the bats in Trinidad and Tobago were reported positive for E. coli.
Earlier study by Gordan and FitzGibbon (1999) revealed that enteric bacteria were distributed according to their animal host or geographical locality where the animal isolate was collected. Factors thought to be responsible for this distribution were the living behavior of the animal hosts and the adaptation of the bacteria in the hosts. The occurrence of E. coli in these animal hosts may be associated with the feeding habits, diet and living behavior such as contact or exposure with livestock or human. In the present study, occurrence of E. coli was consistently higher in rodents in all the three sites of the disturbed habitats. However, when the occurrence was compared between the three disturbed habitats, the occurrence of E. coli in rodents was the highest in the oil palm plantation (48%) where permanent habitation for the workers were more compared to the recreational park and forest park. In this environment, rodents may feed on food sources originated from human wastes, garbage, food waste, or sewage effluent which may serve as sources of E. coli. In addition, rodents lived on land and came into contact with soil and fecal samples, which served as reservoir for enteric pathogen (Santamaria and Toranzos, 2003). The occurrence of the Escherichia coli in birds (32%) was lower than in rodent (43%). The presence of Escherichia coli in birds was attributed to its feeding habits where the bacteria could be picked from rubbish tips, sewage outfalls and consumption of contaminated food (Wallace et al., 1997). It was possible that the recreation park provided more opportunity for the birds to feed thus acquiring a higher proportion of E. coli. Bats were found to harbor the least E. coli (11%) and this could be attributed to their living behavior as the least contact with human. Bats are known to live in roosts, in caves, under storey of forest or hollow trees. In addition, bats have varied feeding habit, for example, insect bats only feed on insects and fruit bats feed on fruits or nectar (Payne et al., 1985), which has the least association with human and contaminated environment. Present results were in agreement with previous reported by Prem-Anand and Sripahi (2004) that feeding habits could be affected by the types and distribution of bacteria in bats.
All of the E. coli isolates were screened for O157:H7 strains. The primary screening of E. coli O157:H7 was carried out by direct culture on CT-SMAC and further confirmed with E. coli O157:H7 latex agglutination kit (Oxoid, England). The results of sorbitol fermenter on CT-SMAC did not show any positive reaction. The CT-SMAC was previously reported to be less sensitive in detection of E. coli O157:H7 (Karch et al., 1996). Therefore, multiplex PCR were used for simultaneous identification of E. coli O157:H7 and its virulence factors in a single reaction tube. Multiplex PCR has been widely used for the detection of E. coli O157:H7 in previous studies through the detection of Shiga-toxin genes and gene encoded for antigen O157 and antigen H7 (Hu et al., 1999; Radu et al., 2001). Results from the multiplex PCR in our present study revealed that none of the isolates harbored genes encoding for shiga toxin 1, shiga toxin 2 and the gene involved in biosynthesis of O157 antigen. These results suggested that none of the isolates tested possessed the virulence factors. However, gene encoded for H7 antigen was detected in 14 E. coli isolates. Although, H7 gene was present in these 14 isolates, they however lacked in the gene encoding for O157 antigen as shown by the multiplex PCR assay. Thus, this E. coli could be identified as non E. coli O157:H7 serotype. It has been revealed that the H7 gene encodes adhesive properties, which contributed to colonization of mucosal surfaces (Erdem et al., 2007). The presence of the gene encoded for H7 antigen in the 14 E. coli isolates may indicated that they possessed flagella which could assist in binding to host cells. However, the absence of the Shiga toxin producing gene suggested that these isolates were not virulent strains.
The absence of STEC or E. coli O157:H7 in the wildlife examined in the present study was in agreement with previous published reports. E. coli O157:H7 was not detected in free-ranging and captive avian and mammalian wildlife of Trinidad and Tobago (Adesiyun, 1999). The absence of E. coli O157:H7 was also reported from pigeons in Colorado (Pedersen et al., 2006). In recent study by Adesiyun et al. (2009), E. coli O157 strain was also not detected in bats in Trinidad and Tobago.
To the best of our knowledge, the present study is the first in Malaysia to assess the occurrence of the pathogenic E. coli from wildlife in a disturbed environment. In the present study, the absence of E. coli O157:H7 from wildlife sampled in the disturbed habitats, which was thought to provide opportunity for higher contact between animals and human, indicated that there is low risk for the transmission of E. coli serotype O157:H7 (which may cause haemorrhagic colitis and haemolytic uremic syndrome) in the environment. Therefore, the three types of wildlife studied here may not play an important role as reservoir in the transmission of the pathogenic strain of E. coli from animal to human or vice versa in the state of Sarawak, Malaysia.
CONCLUSION
In conclusion, E. coli was distributed according to their animal host or habitats of the animal host. The occurrence of E. coli showed that it was always higher in rodent followed by bird and the least in bats regardless of the habitats. Results from this study indicated that bats, birds, or rodents from these habitats are not important reservoir in the epidemiologic cycle of emerging enteric bacterial zoonoses in the state of Sarawak, Malaysia.
ACKNOWLEDGMENTS
This project was funded by UNIMAS grant E14006/F07/06/ZRC/03/2007(03). The authors also would like to thank Mr. Wahab Marni, Mr. Besar Ketol, Mr. Azis Ajim, Mr. Dahlan Rambli and all the staff and students from Zoology Department in the Faculty of Resource Science and Technology, Universiti Malaysia Sarawak who assisted in the sampling trip. Samples collected were approved by the State Planning Unit under permit (13) UPN/S/G1/I/10.1 Vol.26, 37/2008 for Forest Park Entrance Permit and NCCD.907.4 (IV)-29 for sampling in Forest Park by Sarawak Forestry Department, TAP-OP/GM/0508/11 by Ta Ann Plywood Sdn Bhd for sampling permit at oil palm plantation and also Sibu Municipal Council for approval letter to sample at recreational park.
REFERENCES
- Adesiyun, A.A., 1999. Absence of Escherichia coli O157 in a survey of wildlife from Trinidad and Tabago. J. Wildlife Dis., 35: 115-120.
PubMed - Adesiyun, A.A., A.S. Johnson and N.N. Thompson, 2009. Isolation of enteric pathogens from bats in Trinidad. J. Wildlife Dis., 45: 952-961.
PubMed - Caprioli, A., S. Morabito, H. Brugere and E. Oswald, 2005. Enterohaemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res., 36: 289-311.
Direct Link - Chomel, B.B., A. Belotto and F.X. Meslin, 2007. Wildlife, exotic pets and emerging zoonoses. Emerg. Infect. Dis., 13: 6-11.
CrossRefPubMedDirect Link - Erdem, A.L., F. Avelino, J.X. Cortes and J.A Giron, 2007. Host protein binding and adhesive properties of H6 and H7 flagella of attaching and effacing Escherichia coli. J. Bacteriol., 189: 7426-7435.
PubMed - Foster, G., J. Evans, H.I. Knight, A.W. Smith and G.J. Gunn et al., 2006. Analysis of feces samples collected from a wild-Bird garden feeding station in Scotland for the presence of verocytotoxin-producing Escherichia coli O157. Applied Environ. Microbiol., 72: 2265-2267.
PubMed - Gannon V.P., S. D`Souza, T. Graham, R.K. King, K. Rahn and S. Read, 1997. Use of the flagellar H7 gene as target in multiplex PCR assays and improved specificity in identification of enterohaemorrhagic Escherichia coli strains. J. Clin. Microbiol., 35: 656-662.
PubMed - Gordan, D.M. and F. FitzGibbon, 1999. The distribution of enteric bacteria from Australian mammals: Host and geographical effects. Microbiology, 145: 2663-2671.
PubMedDirect Link - Gordon, D.M. and A. Cowling, 2003. The distribution and genetic structure of Escherichia coli in Australian vertebrates: Host and geographic effects. Microbiology, 149: 3575-3586.
CrossRef - Hu, Y., Q. Zhang and J.C. Meitzler, 1999. Rapid and sensitive detection of Escherichia coli O157:H7 in bovine faeces by a multiplex PCR. J. Applied Microbiol., 87: 867-876.
CrossRef - Karch, H., C. Janetzki-Mittmann, S. Aleksic and M. Datz, 1996. Isolation of enterohemorrhagic Escherichia coli O157 strains from patients with hemolytic-uremic syndrome by using immunomagnetic separation, DNA-based methods and direct culture. J. Clin. Microbiol., 34: 516-519.
PubMed - Kuiken, T., F.A. Leighton, R.A.S. Fourchier, J.W. Le-Due and J.S. Peiris et al., 2005. Public health: pathogen surveillance in animals. Science, 309: 1680-1681.
CrossRef - Meng, J., S. Zhao, M.P. Doyle, S.E. Mitchell and S. Kresovich, 1997. A multiplex PCR for identifying Shiga-like toxin-producing Escherichia coli O157:H7. Lett. Applied Microbiol., 24: 172-176.
PubMed - Nielsen, E.M., M.N. Skov, J.J. Madsen, J. Lodal, J.B. Jespersen and D.L. Baggesen, 2004. Verocytotoxin-producing Escherichia coli in wild birds and rodents in close proximity to farms. Applied Environ. Microbiol., 70: 6944-6947.
CrossRef - Pedersen, K., L. Clark, W.F. Andelt and M.D. Salman, 2006. Prevalence of Shiga toxin-producing Escherichia coli and Salmonella enterica in rock pigeons captured in Fort Collins, Colorado. J. Wildlife Dis., 42: 46-55.
Direct Link - Prem-Anand, A.A. and K. Sripahi, 2004. Digestion of cellulose and xylan by symbiotic bacteria in the intestine of the Indian flying fox (Pteropus giganteus). Comp. Biochem. Physiol. A Mol. Integr. Physiol., 139: 65-69.
PubMed - Radu, S., W.L. Ooi, G. Rusul, M.I.A. Karim and M. Nishibuchi, 2001. Detection of Escherichia coli O157:H7 by multiplex PCR and their characterization by plasmid profiling antimicrobial resistance, RAPD and PFGE analyses. J. Microbiol. Methods, 46: 131-139.
CrossRef - Riley, L.W., R.S. Remis, S.D. Helgerson, H.B. McGee and J.G. Wells et al., 1983. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med., 308: 681-685.
PubMedDirect Link - Santamaria, J. and G.A. Toranzos, 2003. Enteric pathogens and soil: A short review. Int. Microbiol., 6: 5-9.
Direct Link