
W.I.A. Saber
Department of Microbiology, Soils, Water and Environment Research Institute, Agricultural Research Center, Giza, Egypt
K.M. Abd El-Hai
Department of Leguminous and Forage Disease Research,Agricultural Research Center, Giza, Egypt
K.M. Ghoneem
Department of Seed Pathology Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Research Journal of Microbiology
Year: 2009 | Volume: 4 | Issue: 8 | Page No.: 286-300







ABSTRACT
Experiments were carried out to correlate the biochemical features of Trichoderma species and Rhizobium leguminosarum to both biocontrol of Botrytis fabae and improving the productivity of faba bean. Of several Trichoderma species, isolated from phyllosphere of faba bean, six isolates, which grew considerably faster than B. fabae and have moderate to very good antagonism against this pathogen, were selected. Trichoderma’s growth inhibiting properties of B. fabae were due to the combined action of non-volatile and volatile metabolites (with antibiotic nature) and the secretion of cell-wall degrading enzymes. Trichoderma viride (tag3 and tag4) and T. harzianum tag7 have shown to be efficient mycoparasites on B. fabae (in which the mycelium appeared to be fragmented hyphae, vacuolated and disrupted as a result of Trichoderma parasitism). These three Trichoderma isolates were further applied in field of faba bean combined with R. leguminosarum which, the chromatographical analysis of its supernatant showed activity in growth promoter substances. The dual inoculation of seeds with a mixture of R. leguminosarum and T. viride tag4 then foliar spraying of the developed plants with the spore suspension of the same T. viride tag4 at the 35th and 55th day from sowing reduced chocolate spot disease and enhanced nodulation, nitrogenase activity and nitrogen fixing bacterial population in the rhizosphere. In addition to the improvements in the physiological activities (photosynthetic pigments, total phenol and polyphenol oxidase), plant growth and yield. On average, this treatment recorded about 57% reduction in chocolate spot disease and 23% increase in faba bean yield, compared to control plants. Therefore, a commercial production of an inoculum based on a mixture of Rhizobium and Trichoderma is very encouraged.
PDF Abstract XML References Citation
How to cite this article
W.I.A. Saber, K.M. Abd El-Hai and K.M. Ghoneem, 2009. Synergistic effect of Trichoderma and Rhizobium on Both Biocontrol of Chocolate Spot Disease and Induction of Nodulation, Physiological Activities and Productivity of Vicia faba. Research Journal of Microbiology, 4: 286-300.
URL: https://scialert.net/abstract/?doi=jm.2009.286.300
URL: https://scialert.net/abstract/?doi=jm.2009.286.300
INTRODUCTION
Trichoderma species are free-living imperfect fungi, which are highly interactive in root, soil and foliar environments. Some of Trichoderma members possess mycoparasitic abilities against pathogenic fungi, which allow for the development of biocontrol strategies based on Trichoderma in order to restore the beneficial balance of natural ecosystems, which is often lost in the crop situation (Harman et al., 2004; Kredics et al., 2004). Trichoderma work against fungal phytopathogens either indirectly by competing for nutrients and space, modifying environmental conditions, promoting plant growth and plant defense mechanisms and antibiosis; or directly through mechanisms such as mycoparasitism (Shakeri and Foster, 2007; Reino et al., 2008).
Recent evidences indicating induced localized and systemic resistance by Trichoderma species in a range of plants to a variety of phytopathogens, in addition to their substantial influence on plant growth and development (Harman et al., 2004; Adams et al., 2007; Reino et al., 2008). Moreover, Trichoderma are active in the production of fungal cell wall degrading enzymes including pectinases, cellulases and chitinases that are involved in biocontrol process (Barbosa et al., 2001). The dual inoculation of legume seeds with chitinolytic and cellulolytic microbes plus Rhizobium not only protects plants against several diseases but also, necessary to improve growth and nodulation (Sindhu and Dadarwal, 2001). Anyhow, now work available about coinoculation of Rhizobium and Trichoderma.
There is an urgent need to improve Vicia faba yield, since this plant remains an important part of the diet of both humans and domestic animals in many parts of the world, because of its high nutritive value in both energy and protein contents. Furthermore, faba bean supplies an important benefit to the crop by fixing atmospheric nitrogen in symbiosis with R. leguminosarum thus, reducing costs and minimizing impact on the environment, that is why increasing the plant production is one of the major targets of the agricultural policy in several countries (El-Fallal and Migahed, 2003; Mahmoud et al., 2004).
Chocolate spot, caused by Botrytis fabae, is a worldwide disease capable of devastating the unprotected faba bean, result in harmful effects on growth, physiological activities and yield. The problem of adequately protecting plants against the fungus by using fungicides has been complicated by development of fungicidal resistance and/or adverse effects on growth and productivity of faba bean as well as on the accompanying microflora. Therefore, controlling B. fabae by biocontrol agents seemed to be better and preferred than the chemical control (El-Fallal and Migahed, 2003; Mahmoud et al., 2004).
Herein, a novel strategy was conducted for controlling chocolate spot disease as well as, enhancing growth, nodulation, physiological activities and yield of faba bean by Trichoderma species and R. leguminosarum.
MATERIALS AND METHODS
Rhizobium leguminosarum and Faba Bean Seeds
Rhizobium leguminosarum bv. vicia was kindly obtained from the Biofertilizer Production Unit, Agric. Res. Center, Giza, Egypt. Seeds of faba bean; Giza 2 (susceptible for chocolate spot infection) were purchased from Legumes Department, Field Crops Research Institute, Agricultural Research Center, Giza, Egypt.
Isolation of B. fabae and Trichoderma Species
Twenty-one isolates of B. fabae were isolated from naturally infected faba bean leaves (showed typical symptoms of chocolate spot disease) collected from different fields of Dakhlia governorate. Leaves were surface sterilized (with 0.5% sodium hypochlorite solution for 1 to 2 min) and plated on faba bean dextrose agar medium (400 g of faba bean leaves (autoclaved and filtered to obtain faba bean infusion), 20 g of dextrose and 18 g agar) at 20°C (Haggag et al., 2006). Isolates were purified and identified according to Morgan (1971). The most aggressive isolate was selected on the base of pathogenicity test.
Eleven Trichoderma isolates (tag1 to tag11) were isolated from the phyllosphere of healthy faba bean plants grown in different fields at Dakhlia governorate, Egypt. Trichoderma species were isolated on the selective medium of Elad et al. (1991). The developed colonies were transferred onto PDA slants and identified after growing them on malt extract agar for two days at 25°C according to Rifai (1969) and Bissett (1991).
Growth Rate and Dual Culture Tests
For growth rate, plates of PDA were inoculated in the center with a 5 mm disc of individual Trichoderma isolate or B. fabae. After 3 days of incubation at 26°C, the colony diameter of each fungus was measured and the growth rate per day was calculated. For dual culture test, plates of PDA were inoculated with a 5 mm disc of B. fabae, 10 mm from the edge of Petri dish. A 5 mm disc of the tested Trichoderma isolate was placed 60 mm from the B. fabae disc, 24 h after the original inoculation. Paired cultures were incubated at 26°C in the light for 5 days. Plates inoculated with B. fabae only served as control. The growth of individual and dual cultures of the fungi were recorded and interaction between dual mycelia scored for degree of antagonism using a scale of 1 to 5 (Bell et al., 1982), where 1 = Trichoderma overgrowing B. fabae and 5 = B. fabae overgrowing Trichoderma. B. fabae developed from plates of dual cultures were then, microscopically investigated and the changes in the mycelium of B. fabae were recorded.
Survival of B. fabae Mycelium when Paired with Conidia of Trichoderma
Mycelial discs (5 mm) of B. fabae were soaked in a conidial suspension of each of Trichoderma isolates (8 day old culture) and then placed in Petri dishes on moistened filter paper for 12 h. Discs were then removed to PDA supplemented with benomyl (1%) to suppress the growth of Trichoderma. Survival was expressed as the percentage of the mycelial discs from which B. fabae grew (Sivan and Chet, 1989).
Antagonistic Activity of Non-Volatile and Volatile Metabolites of Trichoderma
The effect of non-volatile and volatile metabolites of Trichoderma species against B. fabae was tested following the method of Barbosa et al. (2001). For the non-volatile metabolites, mycelial agar plugs removed from the edge of a young culture of Trichoderma species were transferred to the center of Petri dishes (9 cm diameter) containing PDA and a sterilized cellophane disc (9 cm diameter) adjusted on the medium surface, where the antagonist grown for 60 h. Then the cellophane containing the Trichoderma growth was removed and on the same medium, a disc of B. fabae was placed. The control treatments had B. fabae growing similarly on PDA medium where previously there was a cellophane disc without antagonist. For volatile metabolites, two bottoms of Petri dishes containing PDA were individually inoculated with a disc of pathogen and antagonist; the two bottoms were adjusted and attached by tape. The control sets did not contain the antagonist. The cultures were incubated at 26°C and 60% relative humidity under alternating luminosity (12 h light/12 h darkness). Percent of growth inhibition in both assays were estimated after 3 days.
Extraction of Metabolites with Antibiotic Activity from Trichoderma
Trichoderma isolates were grown in Erlenmeyer flasks (1 L) containing 250 mL PD broth. The cultures were incubated at 28°C on shaker at 140 rpm in the dark for 7 days. Culture filtrates were extracted twice by shaking in a separatory funnel with two third of its volume of ethyl acetate (EtOAc). The EtOAc phases were then pooled and concentrated to give a final volume equivalent to a 250-fold concentration of the original volume of the filtrate (Melo and Faull, 2000). Antifungal activity was tested in vitro against B. fabae. The percentage of reduction in growth and spoulation were calculated.
Extracellular Enzymes of Trichoderma and B. fabae
The activities of some extracellular hydrolytic enzymes of Trichoderma isolates and B. fabae were detected by placing mycelial disks of the tested fungi on solid media containing the enzyme substrate and measuring the zone of degraded substrate formed around the colony. Cellulase and protease activities were studied using microcrystalline cellulose and milk agar as substrates, respectively (Barbosa et al., 2001), chitinase using chitin (Ulhoa and Peberdy, 1991) and amylase activity, using soluble starch (Hankin and Anagnostakis, 1975).
Detection of Growth Substances Secreted by R. leguminosarum
Yeast Mannitol medium (Somasegaran and Hoben, 1985) was used for R. leguminosarum cultivation and inoculum preparation. Auxin-like substances in the supernatant of R. leguminosarum were detected colorimetrically at 530 nm using Ehritch reagent (Larsen et al., 1962). The promoter and inhibitor substances were determined chromatographically according to the method of Hartmann et al. (1967).
Field Experiment
Experiments were carried out at Tag El-Ezz, Agric. Res. Station, Dakhlia, Egypt, during 2007/2008 and 2008/2009 seasons. Soil is clayey in texture containing 54.1 clay, 36.5 silt and 9.4% sand. The Ece in soil paste = 6.04 dS m-1, pH in water suspension (1:2.5) = 8.0, ESP = 8.16%, organic matter = 1.67% and CaCO3 = 2.24%. Available N, P and K were 45.64, 11.21 and 283 mg kg-1, respectively. The soil analysis was carried out in Mansoura Lab. for soil fertility.
The aim was to study the effect of seed inoculation (Rhizobium plus Trichoderma) in combined with foliar spraying of faba bean with T. viride tag3, T. viride tag4 or T. harzianum tag7 on natural infection by chocolate spot disease, as well as, on nodulation, physiological aspects, growth and yield. Before seed inoculation, an antagonism test was carried out between Rhizobium and Trichoderma to insure the compatibility between both microbes. At sowing, R. leguminosarum (108 cfu mL-1) was used individually or mixed in equal portion with each of Trichoderma sp. (107 conidia-1) to inoculate faba bean seeds in the presence of Arabic gum (16%). Inoculated seeds were air dried and sawn immediately. In addition to seed inoculation, developed plants from each assigned treatment were sprayed with the spore suspension of the same Trichoderma sp. that was previously used in seed inoculation, Spraying was performed two times with 20 day interval beginning from 35 day after sowing. All other agricultural practices were carried out as usual. Compete randomize plot design was used. Accordingly, the following five treatments were allocated:
| • | Dual inoculation of seeds with a mixture of R. leguminosarum and T. viride tag3 then foliar spraying of the developed plants with the spore suspension of the same T. viride tag3 after 35 and 55 day from sowing (R+Tv.tag3) |
| • | Dual inoculation of seeds with a mixture of R. leguminosarum and T. viride tag4 then foliar spraying of the developed plants with the spore suspension of the same T. viride tag4 after 35 and 55 day from sowing (R+Tv.tag4) |
| • | Dual inoculation of seeds with a mixture of R. leguminosarum and T. harzianum tag7 then foliar spraying of the developed plants with the spore suspension of the same T. harzianum tag7 after 35 and 55 day from sowing (R+Th.tag7) |
| • | Inoculation of seeds with R. leguminosarum only then foliar spraying of the developed plants with the fungicide; kocide 101 (2.5 g L-1) after 35 and 55 day from sowing (R+ kocide) |
| • | Control; inoculation of seeds with R. leguminosarum only then foliar spraying of the developed plants with tap water after 35 and 55 day from sowing (R) |
Disease Assessment
At 50 and 70 day from sowing (15 days after each spray), plants were rated for both Disease Incidence (DI); as the percentage of infected leaves and Disease Severity (DS) according to the scale (class rate from 1 to 9) of Bernier et al. (1993) following the next formula:
where, NPC is the number of plants in each class rate, CR is the class rate, NIP is the number of infected plants and MSC is the maximum severity class rate.
Nodulation Status and Nitrogenase Activity
Samples of faba bean roots were collected after 50 and 70 days from sowing (20 days after each spray) for the determination of number and dry weight (at 70°C until constant weight) of nodules. Nitrogenase activity was determined (Lethbridage et al., 1982) in the second season only.
Physiological Activities of Faba Bean Plants
After 70 days from sowing in the second season, the following physiological activities were tested; (1) Photosynthetic pigments (chlorophyll (Chl) a, b and total Chl and carotenoids) were determined in the blade of the third leaf of plant tip (terminal leaflet) (Mackinney, 1941), (2) Total phenolic compounds were determined in fresh shoot using the folin-ciocalteau reagent (Malik and Singh, 1980) and (3) Polyphenol oxidase was assayed colorimitrically using catichol as substrate (Matta and Dimond, 1963), the activity was expressed as the change in the absorbance of the reaction mixture every 50 sec for 5 min period at 495 nm.
Morphological Characters and Yield and its Components
After 70 days from sowing, growth parameters per plant (height and number of branches and leaves) were measured. At harvest, pods number plant-1 and seeds number pod-1, seed index (weight of 100-seed) and seed yield were recorded.
Microbial Counts
The media of Bridson (1978), Watanabe and Barraquio (1979) and Martin (1950) were used for counting the total bacteria, nitrogen fixers and total fungi, respectively, in the rhizospheric soil of faba bean plants in the second season at the 50th and at 70th day from sowing.
Statistical Analysis
All data were subjected to analysis of variance using the statistical analysis software; CoStat v6.4. Compare among means was carried out using either Tuckey test (for laboratory) or Duncan's new multiple range test (for field).
RESULTS AND DISCUSSION
Growth Rate of Trichoderma Isolates and the Pathogen
From the biological control view point, one of the most important principals for choosing the antagonistic organism is its capability to grow fast and occupy most of the place in the site of probable exist of the pathogen. For this reason, 11 Trichoderma species (isolated from faba bean phyllosphere) and aggressive isolate (isolated from infected faba bean plants) of chocolate spot causal pathogen (B. fabae) were investigated for the growth rate (Fig. 1). Significant difference in growth was observed between individual Trichoderma isolates. However, all Trichoderma isolates grew considerably faster than B. fabae. The rapid growth gives Trichoderma an important advantage in the competition for space and nutrients with plant pathogenic fungi, even before it deploys its arsenal of mycotoxins (Barbosa et al., 2001). Accordingly, the fastest six Trichoderma isolates in growth rate were identified to species level and used for further studies.
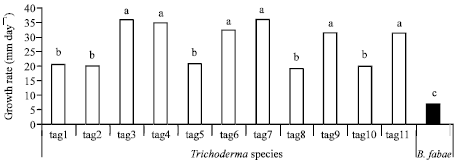 | |
| Fig. 1: | Growth rate of various Trichoderma species and B. fabae on PDA. Different letters indicate significant difference at p≤0.01(Tuckey test) |
Direct Antagonism of Trichoderma Against B. fabae
In dual culture test, the six Trichoderma isolates inhibited the growth of B. fabae (Table 1). T. viride (tag3 and tag4) and T. harzianum tag7 showed high degree of antagonism and reduced the growth of B. fabae by over 60% after 5 days of the dual growth. The best antagonism reaction (1) was recorded by T. viride tag3 and T. harzianum tag7, which means the occurrence of strong mycoparasitism. The light microscope investigation showed B. fabae mycelium to be fragmented hyphae, vacuolated and disrupted. Moreover, when plates were observed for more than 5 days, three species of Trichoderma (tag3, tag4 and tag7) were found to produce inhibition halos and sporulated over the colonies of B. fabae.
| Table 1: | Growth of B. fabae as affected by different Trichoderma species in dual culture test |
 | |
| *(Radius growth of B. Fabae in the direction of Trichoderma/Radius of growth in the absence of Trichoderam)x100. **Numbers refer to antagonism reactions of Trichoderma with B. fabae based on the antagonism scale of Bell et al. (1982) after 5 days of dual growth. Different letters within a column indicate significant difference at p≤0.01 (Tuckey test) | |
These observations were confirmed by studying the survival of B. fabae mycelium when paired with the conidial suspension of Trichoderma (Fig. 2) in which, remarkable inhibition of mycelial growth of B. fabae was observed. T. viride (tag 3 and tag4) and T. harzianum tag7 totally inhibited the mycelial growth of B. fabae. Melo and Faull (2000) recorded similar observations.
Once the fungal pathogen come into contact, Trichoderma sp. attach to and can coil around it, in some cases, form appressoria on the host surface, wherein Trichoderma produce several cell wall degrading enzymes and probably also antibiotics, the combined activities of these compounds result in parasitism and dissolution of the cell walls forming holes which acts as direct entry of Trichoderma hyphae into the target fungus (Melo and Faull, 2000; Harman et al., 2004). This interpretation could be applied only for T. viride (tag3 and tag4), T. harzianum tag7 and Trichoderma sp. tag11. The other two isolates persisted strong antibiosis zone against B. fabae without mycoparasitism.
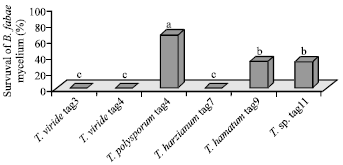 | |
| Fig. 2: | Survival of B. fabae mycelium when paired with conidia of different Trichoderma species. Different letters indicate significant difference at p≤0.01(Tuckey test) |
Antagonistic Activity of Trichoderma Metabolites
Data of Fig. 3 show the effect of non-volatile and volatile metabolites of Trichoderma on B. fabae growth. The non-volatile substances were responsible for most of the growth inhibition, when B. fabae was cultivated in the same Petri dish, where Trichoderma was previously grown, antagonism was readily observed. T. viride (tag3 and tag4) and T. harzianum were more efficient than the other isolates; they reduced the growth of B. fabae by over 70%. For the volatile metabolites, T. viride (tag3 and tag4) and T. harzianum tag7 were the only ones to produce some growth inhibition by volatile substances. These Trichoderma antibiotics were extracted with EtOAc and their impact on the growth of B. fabae, were evaluated. Data of Table 2 show that EtOAc phase extracts of T. viride (tag3 and tag4) and T. harzianum tag7 possessed strong antifungal activity and inhibited B. fabae growth and sporulation by over 73 and 90%, respectively. Trichoderma hamatum did not produce any antibiotic activity.
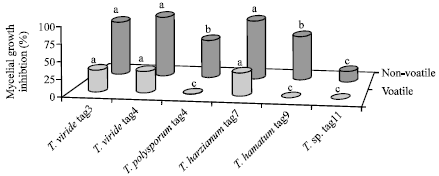 | |
| Fig. 3: | Inhibition of B. fabae growth by non-volatile and volatile metabolites of different Trichoderma species. For each metabolite. Different letters indicate significant difference at p≤0.01(Tuckey test) |
| Table 2: | Antibiotic activity of ethyl acetate extracts (EtOAc phase) of different Trichoderma species against growth and sporulation of B. fabae |
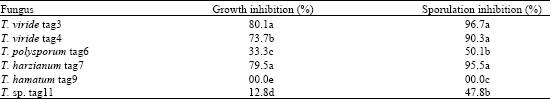 | |
Different letters within a column indicate significant difference at p≤0.01 (Tuckey test) | |
Species of Trichoderma are known to have about 20 to 30 genes, proteins, enzymes and other metabolites that are directly involved in the interaction between Trichoderma and target fungi (Harman et al., 2004). The antifungal activity of Trichoderma metabolites is achieved by biosynthesizing a wide array of non-volatile metabolites e.g., anthraquinones, koninginins, trichodermamides, steroidal antibiotic, viridiofungins, harzialactones, trichothecenes, peptaibols, statins, acoranes and miscellanea, in addition to, volatile metabolites, with strong coconut smell, e.g., simple aromatic compounds, some polyketides (e.g., pyrones (6-pentyl-2-pyrone) and the butenolides), volatile terpenes and the isocyane metabolites (Barbosa et al., 2001; Harman et al., 2004; Shakeri and Foster, 2007; Reino et al., 2008).
Extracellular Enzymatic Activity
The secretion of the extracellular cell wall degrading enzymes were tested for both Trichoderma species and B. fabae (Fig. 4). Species of Trichoderma produced much more cellulolytic, amylolytic and chytinolytic activities than B. fabae. Only the species of Trichoderma secreted amylase and chitinase. With exception to the low proteolytic activity of T. viride (tag3 and tag4) and T. harzianum tag7, protease was, exclusively, secreted by B. fabae only.
Several reports have demonstrated positive relationship between the production of cellulase, chitinase and protease and the ability to control plant diseases, these enzymes are induced in Trichoderma during the parasitic interaction and can inhibit the growth of several fungal plant pathogens by degrading cell walls (Melo and Faull, 2000; Harman et al., 2004; Shakeri and Foster, 2007). The ready expression of amylolytic enzymes in Trichoderna than B. fabae is partially responsible for the rapid growth of Trichoderma on PDA as, previously, shown in Fig. 1. Additionally, there is no immediate explanation for the negative proteolytic activity of some Trichoderma species, except the possible attachment of protease to the mycelium of Trichoderma (Barbosa et al., 2001). Microbes with these enzymatic activities positively induce the nodulation when used in combined with rhizobia to inoculate legumes seeds (Sindhu and Dadarwal, 2001).
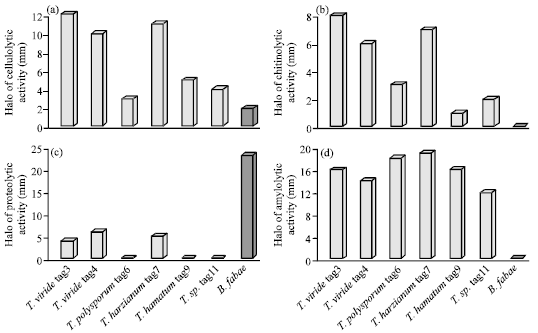 | |
| Fig. 4: | Extracellular enzymatic activities for both Trichoderma species and B. fabae |
Based on the previous results, T. viride (tag3 and tag4) and T. harzianum tag7, as the most efficient mycoparasites and antibiotic producers, were selected to be applied as seed inocluants combined with R. leguminosarumand and, at the same time, as foliar spraying on grown faba bean plants.
Plant Growth Substances of R. leguminosarum
Before using R. leguminosarum in the dual inoculation with the selected Trichoderma species, the growth substances in the supernatant of R. leguminosarum was tested. It is clear from the chromatographical analysis (Fig. 5) that there were growth-promoting substances existing at all values of retention factor (Rf). The net unites of promoter substances being 29.5, while the auxin-like substances being 2.92 mg mL-1. This means that R. leguminosarum has biological activity on faba bean growth. Ahmad et al. (2008) found that 80% of the nitrogen-fixing bacteria produce indol acetic acid. These growth substances lead to increase in total phenol, calcium content and polyphenol oxidase activity, which protect plants against pathogens and improve the growth (Chowdhury, 2003).
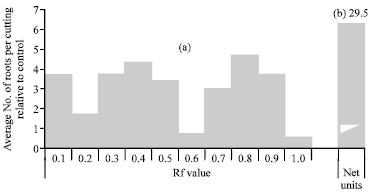 | |
| Fig. 5: | The biological activity of growth substances secreted by R. leguminosarum at (a) different Rf values and (b) net units of growth promoters |
The antagonism between R. leguminosarum and the three Trichoderma isolates that were previously selected from the comparative in vitro experiments was tested. No antagonistic action was observed between R. leguminosarum and any of T. viride (tag3 and tag4) or T. harzianum tag7. So, R. leguminosarum was used to inoculate faba bean seeds in combined with the selected Trichoderma.
Field Evaluation of the Dual Application of R. leguminosarum and Trichoderma Incidence (DI) and Severity (DS) of Chocolate Spot Disease on Faba Bean Plants
Data (Table 3) of the two growing seasons show that, the dual inoculation of seed (R. leguminosarum and Trichoderma) and foliar spraying of grown faba bean with each of Trichoderma isolate (as mentioned in materials and methods) significantly reduced chocolate spot DI and DS caused by B. fabae as compared to control (R). Treatment of faba bean with R+Tv.tag4 reduced DI and DS by 45.3 and 59.6 and 32.8 and 51.0% (average of both seasons) after 50 and 70 days from sowing, respectively. The range of infection level, in the presence of Trichoderma, is economically accepted and could be comparable with that of the treatment with fungicide. In spite of the later recorded the highest reduction in disease parameters. This may be referred to the ability of Trichoderma to cause an inhibition of B. fabae growth (Table 1, 2 and Fig. 2, 3) and secretion of extracellular enzymes (Fig. 4) that can degrade the cell wall of the pathogen (Melo and Faull, 2000).
| Table 3: | Efficacy of dual inoculation of faba bean with R. leguminosarum and different Trichoderma species on chocolate spot disease development in field |
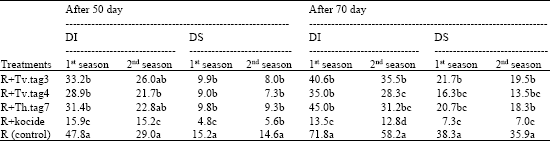 | |
| Different letters within a column indicate significant difference at p≤0.05 (Duncan test) | |
Nodulation Status of Faba Bean
As shown in Table 4 and Fig. 6, the dual inoculation of faba bean seeds with a mixture of R. leguminosarum and each of Trichoderma species followed by foliar spraying with Trichoderma led to significant enhancements in number and dry weight of nodules and nitrogenase activity during the growth period of faba bean. R+Tv.tag4 recorded the highest values of the tested parameters after 50 and 70 days from sowing than that of both fungicide (R+ kocide) and control (R) treatments. Generally, the species of Trichoderma had potential effect on nodulation status.
Chitinase and cellulase may contribute to promotion of nodule formation by the rhizobia after coinoculation with Trichoderma by improving the attack of rhizobia to the specific legume plant and facilitate the penetration of rhizobia into the root hair tissues, causing root hairs more softening and easily curling. Additionally, inoculation by Trichoderma lead to enlargement in root size and rooting depth, all these effects lead to increase the number and biomass of nodules, hence, nitrogenase activity (Sindhu and Dadarwal, 2001; Adams et al., 2007).
| Table 4: | Nodulation status of faba bean as a response to dual inoculation with R. leguminosarum and different Trichoderma species after 70 days from sowing |
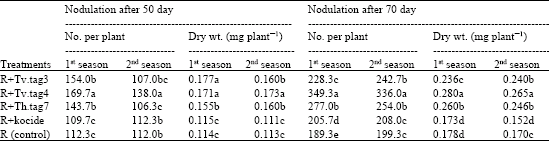 | |
| Different letters within a column indicate significant difference at p≤0.05 (Duncan test) | |
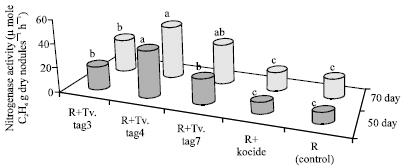 | |
| Fig. 6: | Nitrogenase activity of faba bean nodules after 50 and 70 days from sowing. For each period of time, different letters indicate significant difference at p≤0.05 (Duncan test) |
Physiological Activities of Faba Bean Plants
Photosynthetic pigments is a good parameter reflecting the health condition of plant. As shown in Table 5, Chl a and carotenoids did not significantly affected by any of the treatments. On the other hand, Chl b and total Chl significantly increased in plants developed from faba bean treated with R+Tv.tag4. This increment is by enhancing the efficacy of photosynthetic apparatus with a better potential for disease resistance and decrease in photophosphorylation rate usually occurring after infection (Chandra and Bhatt, 1998).
| Table 5: | Photosynthetic pigments (mg g-1 fresh weight) of faba bean plants as a response of dual inoculation with R. leguminosarum and different Trichoderma species after 70 days from sowing |
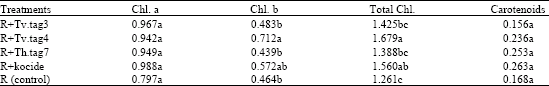 | |
Different letters within a column indicate significant difference at p≤0.05 (Duncan test) | |
Total phenols and polyphenol oxidase activity in faba bean plants were measured (Fig. 7) since they play important role in plant protection against pathogens, both of them were induced greatly because of the dual application of R. leguminosarum and Trichoderma, compared to the fungicide (R+kocide) and control (R) treatments.
The first step of the defense mechanism in plants involves a rapid accumulation of phenols at the infection site, which restricts or slows the growth of the pathogen because of its action as antioxidant, antimicrobial and photoreceptor (Lamba et al., 2008). The suggested mechanisms for the pathogen defense role of polyphenol oxidases (PPO), include; (1) general toxicity of PPO-generated quinones to pathogens and plant cells, accelerating cell death (2) alkylation and reduced bioavailability of cellular proteins to the pathogen (3) cross-linking of quinones with protein or other phenolics, forming a physical barrier to pathogens in the cell wall and (4) quinone redox cycling leading to H2O2 and other reactive oxygen species, which are known to be important factors in plant pathogen interactions and defense signaling (Li and Steffens, 2002; Raj et al., 2006). That is why the levels of both total phenols and PPO are naturally high in resistant varieties (Gogoi et al., 2001; Raj et al., 2006).
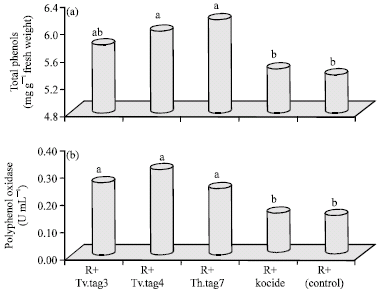 | |
| Fig. 7: | (a) Total phenols and (b) polyphenol oxidase activity in faba bean plants as affected by dual inoculation with R. leguminosarum and different Trichoderma species after 70 days from sowing. In each histogram, different letters indicate significant difference at p≤0.05 (Duncan test) |
Faba Bean Growth, Productivity and Yield
As shown in Tables 6 and 7, seed inoculation with R. leguminosarum plus each of Trichoderma isolate folloewd by foliar spraying of the grown faba bean plants with each of same Trichoderma isolate increased significantly the growth, productivity and yield of faba bean, in this respect, R+Tv.tag4 followed by R+Tv.tag3 gave the highest values of the tested parameters compared with all other treatments. However, number of seeds pod-1 was not affected by any of the treatments. Application of R+Tv.tag4 recorded about 23% (average of the two seasons) increase in faba bean yield compared with that of seed inoculation with R. leguminosarum plus spraying developed faba been plants with the fungicide (R+kocide).
These improvements may be back to the combined action of both R. leguminosarum that stimulated plant growth by nitrogen fixation (Fig. 6) and secretion of growth promoter substances (Fig. 5) and also, due to Trichoderma which have been, recently, reported as plant growth promoting fungi (Adams et al., 2007). However, it is important to note that growth promotion by Trichoderma is not simply a result of reduced pathogenicity (although it is undoubtedly a contributing factor) but it is a combination of direct Trichoderma effects i.e., Trichoderma colonization, control of deleterious microflora, increasing the dissolution of nutrients, increasing the nitrogen usage efficiency, increasing the root size and rooting depth, (which aid nutrient absorption and increase resistance to drought) and increasing the capable of tolerating high levels of metals in metal contaminated soils (Harman, 2000; Harman et al., 2004; Adams et al., 2007).
| Table 6: | Efficacy of dual inoculation with R. leguminosarum and different Trichoderma species on vegetative growth of faba bean after 70 days from sowing |
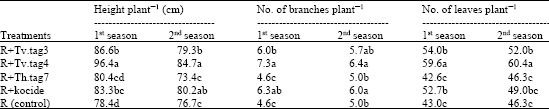 | |
| Different letters within a column indicate significant difference at p≤0.05 (Duncan test) | |
| Table 7: | Efficacy of dual inoculation with R. leguminosarum and different Trichoderma species on faba bean yield and its attributes |
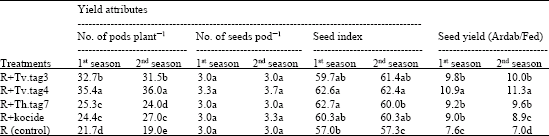 | |
| Different letters within a column indicate significant difference at p≤0.05 (Duncan test) | |
Counts of Microbial Groups
Microbial groups were enumerated in the rhizospheric soil of faba bean plants (Table 8). There were remarkable increases in the counts of total bacteria, total fungi and nitrogen fixers by the time and by the inoculation with Rhizobium combined with Trichoderma. The presence of Trichoderma improved the tested microbial populations, epecially, the ratio of nitrogen fixers to total bacteria which reached its maximum in the presence of Trichoderma inoculation as in the case of R+Tv.tag4; this was in turn reflected on nitrogen fixation and the health and productivity of the plants. These increments in counts of microbial groups may be due to the action of Trichoderma, which colonize on plant root and can control the deleterious microflora as well as increasing root size and depth (Harman et al., 2004; Adams et al., 2007).
| Table 8: | Changes in some microbial populations in the rhizospheric soil of faba bean as a response to dual inoculation of R. leguminosarum and different Trichoderma species |
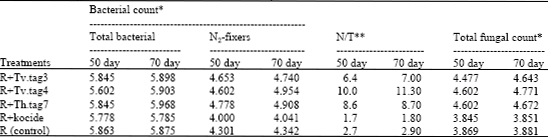 | |
| *The counts of microbial groups were expressed as logarithm cfu g-1. **N/T = (N2-fixers count/total bacterial count)x100. This ratio was calculated from the natural numbers of cfu not from the logarithmic numbers | |
Based on the presented data, the use of T. viride tag4 in combined with R. leguminosarum is an effective strategy for an integrated management of chocolate spot disease as well as increasing growth and productivity of faba bean. Therefore, a commercial production of an inoculum (composed of both microbes) and its incorporation in the production system of Vivia faba is of great benefit.
REFERENCES
- Ahmad, F., I. Ahmad and M.S. Khan, 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res., 168: 173-181.
CrossRefDirect Link - Chandra, A. and R.K. Bhatt, 1998. Biochemical and physiological response to salicylic acid in relation to the systemic acquired resistance. Photosynthetica, 35: 255-258.
CrossRef - Barbosa, M.A.G., K.G. Rehn, M. Menezes and R. de L.R. Mariano, 2001. Antagonism of Trichoderma species on Cladosporium herbarum and their enzymatic characterization. Braz. J. Microbiol., 32: 98-104.
Direct Link - Bell, D.K., H.D. Wells and C.R. Markham, 1982. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology, 72: 379-382.
CrossRefDirect Link - Bissett, J., 1991. A revision of the genus Trichoderma. II. Infrageneric classification. Can. J. Bot., 69: 2357-2372.
CrossRefDirect Link - Chowdhury, A.K., 2003. Control of Sclerotium blight of groundnut by growth substances. Crop Res., 25: 355-359.
Direct Link - El-Fallal, A.A. and F.F. Migahed, 2003. Metabolic changes in broad bean infected by Botrytis fabae in response to mushroom spent straw. Asian J. Plant Sci., 2: 1059-1068.
CrossRefDirect Link - Gogoi, R., D.V. Singh and K.D. Srivastava, 2001. Phenols as a biochemical basis of resistance in wheat against karnal bunt. Plant Pathol., 50: 470-476.
CrossRefDirect Link - Haggag, W.M., A.L. Kansoh and A.M. Aly, 2006. Proteases from Talaromyces flavus and Trichoderma harzianum: Purification, characterization and antifungal activity against brown spot disease on faba bean. Plant Pathol. Bull., 15: 231-239.
Direct Link - Hankin, L. and S.L. Anagnostakis, 1975. The use of solid media for detection of enzyme production by fungi. Mycologia, 67: 597-607.
Direct Link - Harman, G.E., C.R. Howell, A. Viterbo, I. Chet and M. Lorito, 2004. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol., 2: 43-56.
CrossRefPubMedDirect Link - Harman, G.E., 2000. Myths and Dogmas of Biocontrol Changes in Perceptions Derived from Research on Trichoderma harzinum T-22. Plant Dis., 84: 377-393.
CrossRefDirect Link - Hartmann, H.T., M.S. Fadi and W.F. Hackett, 1967. Initiation of flowering and changes in endogenous inhibitors and promoters in olive buds as a result of chilling. Physiol. Plant, 20: 746-759.
CrossRefDirect Link - Kredics, L., L. Manczinger, Z. Antal, Z. Penzes, A. Szekeres, F. Kevei and E. Nagy, 2004. In vitro water activity and pH dependence of mycelial growth and extracellular enzymse activities of Trichoderma strains with biocontrol potential. J. Appl. Microbiol., 96: 491-498.
Direct Link - Lamba, P., S. Sharma, G.D. Munshi and S.K. Munshi, 2008. Biochemical changes in sunflower plants due to seed treatment/spray application with biocontrol agents. Phytoparasitica, 36: 388-399.
CrossRefDirect Link - Larsen, P., A. Harbo, S. Klungsour and T. Asheim, 1962. On the biogenesis of some indole compounds in Acetobacter Xylinum. Physiol. Plant, 15: 552-565.
CrossRefDirect Link - Li, L. and J.C. Steffens, 2002. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta, 215: 239-247.
CrossRefDirect Link - Mackinney, G., 1941. Absorbtion of light by cholorophyll solution. J. Biol. Chem., 140: 315-322.
Direct Link - Mahmoud, Y.A.G., M.K.H. Ebrahim and M.M. Aly, 2004. Influence of plant extracts and microbioagents on physiological traits of faba bean infected with Botrytis fabae. J. Plant Biol., 47: 194-202.
CrossRefDirect Link - Martin, J.P., 1950. Use of acid, rose bengal and streptomycine in the plate method for estimating soil fungi. Soil Sci., 69: 215-232.
CrossRef - De Melo, I.S. and J.L. Faull, 2000. Parasitism of Rhizoctonia solani by strains of Trichoderma spp. Sci. Agric., 57: 55-59.
CrossRefDirect Link - Morgan, D.J., 1971. Numerical toxonomic studies of the genus Botrytis: II. Other Botrytis taxa. Trans. Br. Mycol. Soc., 56: 327-335.
CrossRef - Raj, S.N., B.R. Sarosh and H.S. Shetty, 2006. Induction and accumulation of polyphenol oxidase activities as implicated in development of resistance against pearl millet downy mildew disease. Funct. Plant Biol., 33: 563-571.
Direct Link - Reino, J.L., R.F. Guerrero, R. Hernandez-Galan and I.G. Collado, 2008. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev., 7: 89-123.
CrossRefDirect Link - Shakeri, J. and H.A. Foster, 2007. Proteolytic activity and antibiotic production by Trichoderma harzianum in relation to pathogenicity to insects. Enzyme Microbial. Technol., 40: 961-968.
Direct Link - Sindhu, S.S. and K.R. Dadarwal, 2001. Chitinolytic and cellulolytic Pseudomonas sp. antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in chickpea. Microbiol. Res., 156: 353-358.
CrossRefDirect Link - Ulhoa, C.J. and J.F. Peberdy, 1991. Regulation of chitinases synthesis in Trichoderma harzianum. J. Gen. Microbiol., 137: 2163-2169.
Direct Link - Watanabe, I. and W.L. Barraquio, 1979. Low levels of fixed nitrogen required for isolation of free-living N2-fixing organisms from rice roots. Nature, 277: 565-566.
CrossRefDirect Link