
H. Sasaki
College of Science and Engineering, Iwaki Meisei University, Iwaki, Fukushima 970-8551, Japan
E. Iwata
College of Science and Engineering, Iwaki Meisei University, Iwaki, Fukushima 970-8551, Japan
A. Oshima
Faculty of Life and Environmental Science, Shimane University, Matsue, Shimane 690-8504, Japan
A. Ishida
Graduate School of Science and Technology, Kumamoto University, Kumamoto 860-8555, Japan
S. Nagata
Research Journal of Microbiology
Year: 2009 | Volume: 4 | Issue: 7 | Page No.: 260-268







ABSTRACT
We tried the isolation of halophilic bacteria from Asian desert dust falls in Japan and growth property of these bacteria and their molecular phylogeny were analyzed. Two Gram-positive bacteria designated as IMU-1 and IMU-2 were isolated from Asian desert dust. These two strains were adapted with 0-3 and 0-4 M NaCl under nutrient medium culture conditions, respectively, showing the properties of halotolerance. Under the Davis minimal medium culture condition, IMU-1 attained to the similar level of growth as that of nutrient medium culture and growth was observed at 0-2.5 M NaCl. On the other hand, IMU-2 showed the different growth as that of nutrient medium culture condition and growth was observed at 0-1.2 M NaCl. Phylogenetic analysis using 16S rRNA gene sequences revealed that IMU-1 and IMU-2 belong to Bacillus licheniformis and B. megaterium, respectively. It was first study about the isolation of B. licheniformis as the halophilic bacteria in Japan.
PDF Abstract XML References Citation
How to cite this article
H. Sasaki, E. Iwata, A. Oshima, A. Ishida and S. Nagata, 2009. Isolation of Extreme Halotolerant Bacteria from Asian Desert Dust; Molecular Phylogeny and Growth Properties of their Cells. Research Journal of Microbiology, 4: 260-268.
URL: https://scialert.net/abstract/?doi=jm.2009.260.268
URL: https://scialert.net/abstract/?doi=jm.2009.260.268
INTRODUCTION
Halophilic bacteria are adapted to high osmolarity and can grow in high saline environments. These bacteria are categorized slightly, moderately or extremely halophiles, by the extent of their halotolerant characteristics (Larsen, 1986). These bacteria were isolated from saline environments such as ocean, salt lakes and salt fields (Mimura and Nagata, 2000; Carrasco et al., 2006; Yuan et al., 2007; Fukushima et al., 2007). Halophilic bacteria are able to grow at higher saline environments due to synthesize compatible solutes in cells or possess the transporters to take them up in the medium. Such osmoregulatory solutes as potassium ion, glutamate, proline, ectoine, betaine in bacteria have been extensively reported for bacteria (Galinski, 1993).
Recently, many halophilic bacteria have been isolated from non-saline places and they are not expected to grow there. Echiga et al. (2005) reported the isolation of some halophilic bacteria from garden soil in Japan. They presume that a possible source of the halophilic bacteria is originated from Asian desert dust. Asian desert dust that originates in desert regions of China carry desert aerosols and possibly impact ecosystem in the downwind environment of Japan (Iwasaka et al., 1988). Really, halophilic bacteria have been detected from Asian desert dust gathered in the Chinese sky (Maki et al., 2008; Kakikawa et al., 2009). Furthermore, it was reported that number of airborne microorganisms of the day when Asian desert dust falls tends to increase in comparison with another days in Korea (Choi et al., 1997). In this study, we tried the isolation of halophilic bacteria from Asian desert dust falls in Japan and growth property of these bacteria and their molecular phylogeny were analyzed.
MATERIALS AND METHODS
Collection of Asian Desert Dust and Isolation of Bacteria
It is the season when Asian desert dust falls in Japan in spring. Sampling was performed on Iwaki, Fukushima Pref. and Kobe, Hyogo Pref., Japan on April 2006. Filter paper in glass Petri dish was left the outside for 48 h. Filter paper with dust was directly applied to nutrient medium (pH 7.0) consisting of 5 g L-1 dry yeast extract D-3 (Nihon Pharm. Co., Tokyo, Japan), 10 g L-1 polypepton (Nihon Pharm. Co.) and 2 g L-1 NaCl (Nagata et al., 2005) supplemented with 2 M NaCl. Colonies with different morphologies on agar medium after incubation at 37°C were selected. Hacker’s modified Gram stain was used for the staining of isolated bacteria as well as the studying their morphology.
Culture Conditions
Isolated bacterial strains were pre-cultured in a nutrient medium supplemented with 2 M NaCl. Cell growth was initiated by inoculating pre-culture (0.5 %) in a nutrient medium or Davis minimal medium (pH 7.4) that contains 7 g L-1 K2HPO4, 3 g L-1 KH2PO4, 0.1 g L-1 MgSO4•7H2O, 1 g L-1 (NH4)2SO4, 0.5 g L-1 sodium citrate and 2 g L-1glucose (Davis and Mingioli, 1950). The incubation was performed with 120 strokes at 37°C in a 12x180 m m-1 test tube with 3 mL medium. Growth was determined by measuring the optical density at 600 nm (OD600) using a spectrophotometer PD-303 (Apel, Saitama, Japan).
DNA Extraction and PCR Amplification of Partial 16S rDNA
Genomic DNA was extracted using DNeasy Tissue Kit (Qiagen, Hilden, Germany). Partial 16S rRNA gene was amplified by PCR with Takara Ex Taq Reaction Kit (Takara Bio, Shiga, Japan). Total PCR reaction volume of 30 μL was composed of 3.0 μL 10X Ex Taq Buffer, 3.0 μL dNTP mixture, 2.1 pmol of each primer, 0.8 units Ex Taq and 1.0 μL DNA solution containing 0.15 μg DNA. Degenerate oligonucleotide primers (27f; 5'-GAGTTTGATCCTGGCTCA-3', 1385r; 5’-CGGTGTGTRCAAGGCCC-3’) were used in PCR (Hentschel et al., 2001). The profile of PCR conditions was as follows: initial denaturation at 95°C for 5 min; 35 cycles of denaturation at 95°C for 30 sec, annealing at 54°C for 30 sec, extension at 72°C for 1 min and a final extension at 72°C for 7 min. The PCR product of partial 16S rRNA gene was ligated into the PCR 2.1-TOPO vector by the use of TOPO TA Cloning (Invitrogen, CA, USA). The sequence of partial 16S rRNA gene was confirmed by DNA sequencing with ABI PRISMTM 3730xl DNA Analyzer (Applied Biosystems, CA, USA).
Phylogenetic Analysis
The 16S rRNA gene sequences were compared with all other known rRNA gene sequences through a BLAST search (http://www.ddbj.nig.ac.jp/search/blast-j.html). Similar rDNA sequences were downloaded from the database and aligned with our sequences. The Clustal W program (Thomson et al., 1994) was used for preliminary DNA sequence alignment, followed by a manual final alignment. The aligned sequences were subjected to Maximum Parsimony (MP) analyses in a general heuristic search using PAUP v. 4.0b3a (Swofford, 1999). Twenty random taxon addition replicates were performed in each heuristic search with the Goloboff fit criterion (k = 2), using the TBR branch-swapping option. Gaps were considered as missing data in the MP analysis. From the same alignment, two-parameter distances (Kimura, 1980) between taxa were estimated and a phylogenetic tree was constructed with the neighbour-joining (NJ) method, using PAUP. The robustness of the resulting trees was tested by a bootstrap analysis (Felsenstein, 1985) with 1000 resamplings.
RESULTS
Isolation of Bacteria
Based on differences in colony morphology and pigmentation, we isolated two strains, which were designated to IMU-1 and IMU-2. IMU-1 was originated in dusts that were collected in Kobe, Japan. IMU-2 was isolated from dusts that were collected in Iwaki, Japan. These two strains were gram positive (Fig. 1a, b).
Growth Property of IMU-1
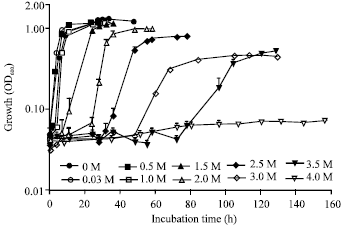
The effect of NaCl on the growth of IMU-1 strain in nutrient medium or Davis minimal medium was examined. As shown in Fig. 2, strain IMU-1 showed the adaptation towards the wide range of NaCl concentrations under nutrient medium culture condition. At 0 to 3.0 M NaCl, sufficient growth was observed, although time lag of growth became longer in proportion to the increase of NaCl concentration in medium. Distinct decrease in growth yield was observed in the presence of 3.5 M NaCl, where growth of strain IMU-1 was not permitted at least during 160 h of incubation. This data suggested strain IMU-1 belongs to halotolerant bacteria.
Under the Davis minimal medium culture condition, IMU-1 attained to the similar growth as that of nutrient medium culture, although the occurrence of a little growth lag time and the reduction of growth rate were recognized (Fig. 3), that is growth was observed at 0-2.5 M NaCl and distinct decrease in growth yield was observed in the presence of 2.8 M NaCl. The impaired growth at 2.8-3.0 M NaCl, however, was recovered to the normal level in growth yield inspite of the occurrence of lag time when 1 mM proline, glutamate, or betaine was supplemented (Fig. 4a-d).
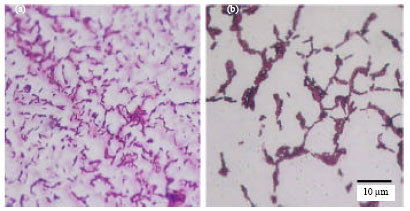 | |
| Fig. 1: | Gram stain of the strain (a) IMU-1 and (b) IMU-2 cells |
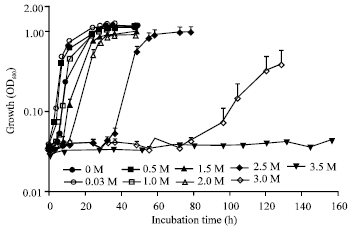 | |
| Fig. 2: | Effect of NaCl concentrations on the growth of the strain IMU-1 in nutrient medium. The cells were grown at 37°C in nutrient medium supplemented with 0-3.5 M NaCl. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
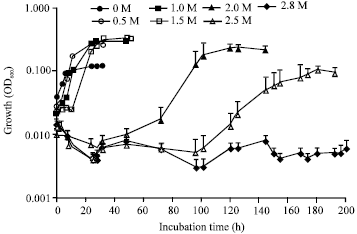 | |
| Fig. 3: | Effect of NaCl concentrations on the growth of the strain IMU-1 in Davis minimal medium. The cells were grown at 37°C in Davis minimal medium supplemented with 0-2.8 M NaCl. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
Growth Property of IMU-2
The effect of NaCl on the growth of IMU-2 strain in nutrient medium or Davis minimal medium was examined. The strain IMU-2 showed the adaptation towards the wide range of NaCl concentrations, which was almost same as IMU-1. Under nutrient medium culture condition, sufficient growth was observed at 0-3.5 M NaCl, although time lag of growth became longer in proportion to the increase of NaCl concentration in medium (Fig. 5). Distinct decrease in growth yield was observed in the presence of 4.0 M NaCl. This data suggested strain IMU-2 belongs to halotolerant bacteria.
Under the Davis minimal medium culture condition, IMU-2 showed the different growth as that of nutrient medium culture. That is, growth was observed at 0-1.2 M NaCl and distinct decrease in growth yield was observed in the presence of 1.5 M NaCl (Fig. 6).
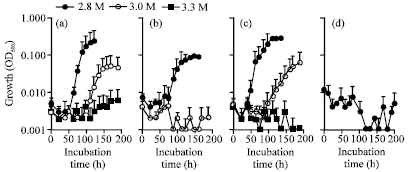 | |
| Fig. 4: | Effect of compatible solutes on the growth of the strain IMU-1 in the presence of 2.8-3.3 M NaCl. The cells were grown at 37°C in Davis minimal medium containing 1 mM each of (a) proline (b) glutamate (c) betaine and (d) ectoine. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
 | |
| Fig. 5: | Effect of NaCl concentrations on the growth of the strain IMU-2 in nutrient medium. The cells were grown at 37°C in nutrient medium supplemented with 0-4.0 M NaCl. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
The impaired growth at 1.5-1.8 M NaCl, however, was recovered when 1 mM proline or ectoine was supplemented (Fig. 7a, b).
Phylogenetic Analysis
For phylogenetic characterization, the 16S rDNA sequences of 15 species were compared with the GeneBank database using the blast program, including two newly reported sequences. The aligned sequences comprised a total of 1399 sites and contained 195 parsimony-informative nucleotide positions. In MP analysis, 3 maximum parsimonious (645 steps, CI = 0.6326, RI = 0.6003) trees were found. Tree topologies were essentially the same for maximum parsimony and neighbour-joining analyses (Fig. 8). Molecular phylogenetic analysis showed all bacterial isolates belong to the family Bacillaceae. Neighbor-joining analysis of 16S rDNA sequences revealed that strains IMU-1 and IMU-2 belong to Bacillus licheniformis and B. megaterium, respectively. The strain IMU-1 showed 99.9% sequence similarity to B. licheniformis.
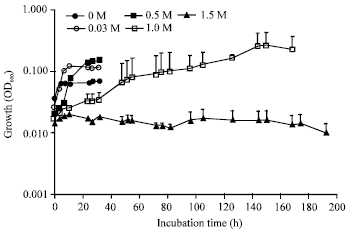 | |
| Fig. 6: | Effect of NaCl concentrations on the growth of the strain IMU-2 in Davis minimal medium. The cells were grown at 37°C in Davis minimal medium supplemented with 0-1.5 M NaCl. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
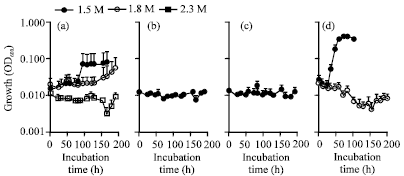 | |
| Fig. 7: | Effect of compatible solutes on the growth of the strain IMU-2 in the presence of 1.5-2.3 M NaCl. The cells were grown at 37°C in Davis minimal medium containing 1 mM each of (a) proline, (b) glutamate, (c) betaine and (d) ectoine. Cell growth was determined by measuring the OD600 as a function of incubation time. The values are the averages±standard deviations from five separate incubations |
The isolate IMU-2 showed 99.9% sequence similarity to B. megaterium. The 16S rRNA gene sequence data reported in this study was submitted to the DDBJ/EMBL/Gen-Bank database and was assigned the accession number AB513628 to IMU-1 and AB513629 to IMU-2.
DISCUSSION
In this study, we isolated bacteria from Asian desert dust falls in Japan and identified two halotolerant Bacillus species, B. licheniformis and B. megaterium. Bacillus species form the spore when incubated in ambient air. It is thought that bacterial spores stick to dusts and come flying from the Eurasian Continent.
In strain IMU-1, growth characteristics suggests that proline, glutamate and betaine were taken up into the cell cytoplasm and played as compatible solute under high concentrations of NaCl. Furthermore, because of the same growth property between nutrient medium and Davis minimal medium, it was thought that IMU-1 cells adapted with high concentrations of NaCl by using compatible solutes produced in cells.
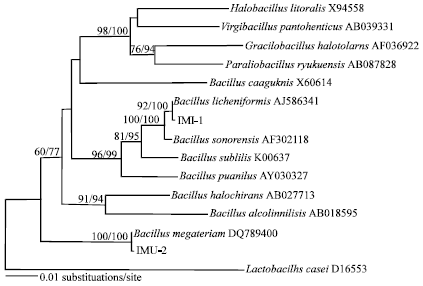 | |
| Fig. 8: | Neighbor-joining analysis of 16S rDNA sequences from several species of Bacillaceae and the strains IMU-1 and IMU-2 isolated from Asian desert dust. The scale bar indicates 1% sequence divergence. Numbers at nodes represent MP bootstrap (left) and NJ bootstrap (right) values in percentage based on 1000 replicates |
On the other hand, in strain IMU-2, growth properties suggest that proline and ectoine were accumulated in the cells by transporters and played as compatible solute under high concentrations of NaCl. Because of the different characteristics of growth in both media, it was thought that IMU-2 cells adapted with high concentrations of NaCl by using compatible solutes produced in cells in case of nutrient medium, or accumulated from nutrient medium. It was reported that compatible solutes such as proline, betaine and glutamate were presence abundantly in the yeast extract that was the constitution ingredient of the nutrient medium (Ishida et al., 1996).
Bacillus licheniformis and B. megaterium produce the proline and glutamate as compatible solutes in cells (Kuhlmann and Bremer, 2002) and this character is almost consistent with growth properties of strains IMU-1 and IMU-2 under high salinity. Strain IMU-1 used proline and glutamate as compatible solute in Davis minimal medium culture condition. In strain IMU-2, proline was effective as compatible solute, but glutamate did not have it. Although, Escherichia coli cells produce glutamate as compatible solute, the addition of the glutamate to the medium is ineffective in cell growth under high salinity (Nagata et al., 2002).
Echiga et al. (2005) reported the isolation of some halophilic bacteria including B. megaterium from Japanese soil, although B. licheniformis was not detected. It was first report about the isolation of B. licheniformis as the halophilic bacteria in Japan. Since, it is know however, that these two species are distributed on the Japanese soil widely (Watanabe and Hayano, 1993), the possibility of the Japanese soil origin should be taken into consideration.
In conclusion, elucidation of the origin of the halophilic bacteria clarifies influence on Japanese ecosystem of the Asian desert dusts. Above-mentioned considerations invite further empirical investigation. It is thought that the collection of the Asian desert dusts in the Japanese sky develops the study.
REFERENCES
- Carrasco, I.J., M.C. Marquez, X. Yanfen, Y. Ma and D.A. Cowan et al., 2006. Gracilibacillus orientalis sp. nov., a novel moderately halophilic bacterium isolated from a salt lake in Inner Mongolia, China. Int. J. Syst. Evol. Microbiol., 567: 599-604.
PubMed - Davis, B.D. and E.S. Mingioli, 1950. Mutants of Escherichia coli requiring methionine or vitamin B12. J. Bacteriol., 60: 17-28.
PubMed - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Galinski, E.A., 1993. Compatible solutes of halophilic eubacteria: Molecular principles, water-solute interaction, stress protection. Cel. Mol. Life Sci., 49: 487-496.
CrossRef - Hentschel, U., M. Schmid, M. Wagner, L. Fieseler, C. Gernert and J. Hacker, 2001. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol., 35: 305-312.
PubMed - Ishida, A., N. Otsuka, S. Nagata, K. Adachi and H. Sano, 1996. The effect of salinity stress on the accumulation of compatible solutes related to the induction of salt-tolerance in Escherichia coli. J. Gen. Applied Microbiol., 42: 331-336.
Direct Link - Kakikawa, M., F. Kobayashi, T. Maki, M. Yamada and T. Higashi et al., 2009. Dustborne microorganisms in the atomosphere over an Asian dust source region, Dunhuang. Air Qual. Atoms. Health, 1: 195-202.
CrossRefDirect Link - Kimura, M., 1980. A simple method for estimating rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120.
PubMed - Maki, T., S. Susuki, F. Kobayashi, M. Kakikawa and M. Yamada et al., 2008. Phylogenetic diversity and vertical distribution of a halobacterial community in the atmosphere of an Asian dust (KOSA) source region, Dunhuang City. Air Qual. Atoms. Health, 1: 81-89.
CrossRef - Mimura, H. and S. Nagata, 2000. Isolation of halotolerant microorganisms from seawater around the Inland Sea in Western Japan. Microb. Environ., 15: 217-221.
CrossRef - Nagata, S., H. Sasaki, A. Oshima, S. Takeda, Y. Hashimoto and A. Ishida, 2005. Effect of proline and K+ on the stimulation of cellular activities in Escherichia coli K-12 under high salinity. Biosci. Biotechnol. Biochem., 69: 740-746.
PubMedDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Watanabe, K. and K. Hayano, 1993. Distribution and identification of proteolytic Bacillus spp. in paddy field soil under rice cultivation. Can. J. Microbiol., 39: 674-680.
PubMed - Yuan, S., P. Ren, J. Liu, Y. Xue, Y. Ma and P. Zhou, 2007. Lentibacillus halodurans sp. nov., a moderately halophilic bacterium isolated from a salt lake in Xin-Jiang, China. Int. J. Syst. Evol. Microbiol., 57: 485-488.
PubMed