
W.I.A. Saber
Department of Microbiology, Soils, Water and Environment Research Institute, Agricultural Research Center, Giza, Egypt
K.M. Ghanem
Deparment of Environment and Bio-Agric, Faculty of Agriculture, AL-Azher University, Cairo, Egypt
M.S. El-Hersh
Department of Microbiology, Soils, Water and Environment Research Institute, Agricultural Research Center, Giza, Egypt
Research Journal of Microbiology
Year: 2009 | Volume: 4 | Issue: 7 | Page No.: 235-250







ABSTRACT
Isolation and identification of rock phosphate (RP) solubilizing fungi were studied under laboratory conditions. Fungal isolates that displayed the highest ratio of clear zone/colony diameter on plates of phosphate solubilization medium, were selected and identified as Aspergillus niger and Penicillium sp. The optimum condition for RP solubilization were found to be at the 6th (A. niger) and 7th (Penicillium sp.) day of incubation with shaking (150 rpm) at 30°C and pH ranging from 5.6 to 6.0. Glucose followed by fructose and xylose supported the RP solubilization process in the presence of 2.5 g L-1 RP as the optimum concentration. The overall soluble P after optimization studies on RP were 99.7 (A. niger) and 77.5 mg L-1 (Penicillium sp.). During the fermentation process, there was remarkable reduction in the final culture pH. The titratable acidity was positively correlated with RP solubilization. Under NaCl salt stress both fungi were able to solubilize RP, in which, A. niger was more tolerant than Penicillium sp. The dual and individual cultures of fungi solubilized sources of phosphate commonly exist in soil and also, possessed phytase activity. Under in vivo conditions, the inoculation of mung bean seeds with A. niger and/or Penicillium sp. in the presence of RP or calcium superphosphate (CSP), increased significantly the growth (except for branches No. plant-1), seed yield and P-uptake, as well as, improved the nodulation status and population of total and phosphate dissolving fungi in the rhizospheric soil of mung bean. These inoculations saved about 1/3 phosphate fertilizer dose. Hereby, these combined effects encourage the potential use of the isolated fungi in the biosolubilization of RP in soil plant system.
PDF Abstract XML References Citation
How to cite this article
W.I.A. Saber, K.M. Ghanem and M.S. El-Hersh, 2009. Rock Phosphate Solubilization by Two Isolates of Aspergillus niger and Penicillium sp. and their Promotion to Mung Bean Plants. Research Journal of Microbiology, 4: 235-250.
URL: https://scialert.net/abstract/?doi=jm.2009.235.250
URL: https://scialert.net/abstract/?doi=jm.2009.235.250
INTRODUCTION
Phosphorus (P) is one of the major nutrient elements limiting agricultural production in the world. Phosphorus entering in the composition of adenosine triphosphate (ATP) which is considered by biologists to be the energy currency of life and essential for all the physiological mechanisms that require energy for different operations. Phosphorus, however, plays an important role in N2-fixation process, as the reduction of atmospheric nitrogen requires large amount of energy. It is estimated that 16 molecules of ATP are required for the reduction of one molecule of atmospheric nitrogen (Krishnan and Bennett, 2006). On average, soil contains 0.02 to 0.5% total P. It is added to the soil in the form of phosphate fertilizers, a part of which (1%) is utilized by plants and the rest is rapidly converted into insoluble complexes e.g., calcium phosphate, iron phosphate and aluminum phosphate in the soil. This leads to the need of frequent application of phosphate fertilizers, but its use on a regular basis has become a costly affair and environmentally undesirable. Therefore, the necessity to develop economical and eco-friendly technologies is steadily increasing (Vassileva et al., 1998; Reddy et al., 2002; Chuang et al., 2007).
Natural phosphate rocks have been recognized as a valuable alternative for P fertilizers. In recent years, the possibility of practical use of rock phosphates (RP) as fertilizers has received significant interest. Unfortunately, RP is not plant available in soils with a pH greater than 5.5 to 6.0 and, even when conditions are optimal, plant yields are lower than those obtained with soluble phosphate (Khasawneh and Doll, 1978). Conventionally, RP is chemically processed by reacting with sulphuric acid or phosphoric acid into soluble phosphate fertilizer. The process increases fertilizer cost and makes the environment worse (Reddy et al., 2002; Chuang et al., 2007; Xiao et al., 2008).
An alternative, has been the use of microorganisms with the capability to solubilize RP and release soluble P through the production of organic acids, chelating oxo acids from sugars, reduction of pH and production of enzymes. Several reports have indicated that some microorganisms are capable of solubilizing insoluble RP and releasing soluble P. However, few reported microorganisms represent a high potential to release soluble P from RP and this seriously restrains the biosolubilization of RP and its use as biofertilizer, hence, isolation and application of new and potential phosphate solubilizing microorganisms are significant and necessary (Son et al., 2006; Achala et al., 2007; Xiao et al., 2008). Filamentous fungi are widely used as producers of organic acids, particularly black Aspergilli and some species of Penicillium, these species have been tested for solubilization of RP and have been reported for various properties of biotechnological importance, such as, biocontrol, biodegradation, phosphate solubilization and P fertilizer (Narsian and Patel, 2000; Reddy et al., 2002; Chuang et al., 2007; Richa et al., 2007; Pandey et al., 2008).
Mung bean (Vigna radiata L.) is a new introduced summer pulse crop in Egypt with short growing season and high nutritive value, grown principally for its protein rich edible seeds, this crop can be used for both seed and forage production. It plays an important role not only in human diet, but also in improving the soil fertility by fixing atmospheric nitrogen with the association of Rhizobium species present in the nodules of its roots (Thalooth et al., 2006).
Therefore, this study was conducted to isolate fungi capable of solubilizing RP and studying the biosolubilization process, especially, under stress conditions as well as testing the response of mung bean plants to inoculation by these isolates.
MATERIALS AND METHODS
Media
National Botanical Research Institute's phosphate growth medium (NBRIP) contained L-1: glucose, 10 g; Ca3 (PO4)2 (TCP) 5 g; MgCl2.6H2O, 5 g; MgSO4.7H2O, 0.25 g; KCl, 0.2 g and (NH4)2SO4, 0.1 g.; distilled water, 1000 mL; pH 7.0 was used (Nautiyal, 1999). Plates were prepared by adding agar (1.5%) to NBRIP medium.
Rock Phosphate and Mung Bean Cultivar
RP was kindly obtained from Soils, Water and Environment Research Institute, Agric. Res. Center, Giza, Egypt and analyzed for its P content (7.97%) and added to the growth medium instead of TCP in the amount equivalent to 50 mg P2O5 100 mL-1 (Reddy et al., 2002). Mung bean cultivar; Kawmy 1 was kindly obtained from Legumes Dept., Crop Res. Institute, Agric. Res. Center, Giza Egypt.
Isolation and Identification of Phosphate Solubilizing Fungi (PSF)
Soil samples characterized with high percentage of RP were incorporated in NBRIP agar medium and incubated at 30°C for 5 days. PSF were shown by the formation of a clear zone around fungal colonies. Colonies with clear zones were further purified by replanting on medium mentioned above. Comparisons of phosphate solubilizing capabilities were carried out on the base of ratio of clear zone diameter/colony diameter of each isolate. Two PSF were thus screened and identified as A. niger and Penicillium sp. (Domsch et al., 1980) and used for further studies. The isolates were maintained on PDA slants and sub-cultured weekly.
Solubilization Conditions of RP
The abilities of the tested fungi to dissolve the RP were determined in 250 mL Erlenmeyer flask with 50 mL of the sterile NBRIP broth medium described above and using RP (50 mg P2O5 100 mL-1) as sole P source. Spore suspensions of the isolates were adjusted by a haemocytometer to approximately 2.5x106 spores mL-1. Each flask was inoculated with the spore suspension at 10% (v/v) and incubated at 30°C on a rotary shaker (120 rpm).
The Tested Parameters
The investigated parameters were carried out in NBRIP broth medium as follows: (1) time course of RP solubilization (1 to 9 days), (2) shaking speeds (60 to 180 rpm), (3) incubation temperature (20 to 45°C), (4) initial culture pH (4.5 to 8, using HCl or NaOH), (5) carbon sources at 1% (Xylose, Glucose, Galactose, Fructose, Maltose, Sucrose and Starch), (6) RP concentration (0.5 to 3.5 gL-1), (7) NaCl stress (1 to 7%) and (8) solubilization of different common sources of P i.e., Ca3(PO4)2, aluminium phosphate, ferrous phosphate, sodium phytate in addition to RP. It is worthy to mention that, before using the dual culture of both fungi, an antagonism test was carried out on PDA plates.
After incubation, suspended mycelium was carefully harvested from the medium by centrifugation (4000 rpm), washed several times with distilled water and dried in an oven at 105°C to constant weight. The supernatant was collected to be analyzed for pH (HI 9321 microprocessor pH-meter), titratable acidity (TA) by the method of Cerezine et al. (1988) and soluble phosphate (Jackson, 1967). The uninoculated autoclaved medium with phosphate substrate was incubated under similar conditions to serve as the control.
Assay of Phytase
Phytase activity was assayed in the supernatant of A. niger and Penicillium sp. using sodium phytate as substrate (El-Sawah et al., 2001). One unit of phytase activity was defined as the amount of enzyme releasing one μ mole of inorganic phosphorus/mL/min.
Field Experiment
Field experiment was conducted at Tag El-Ezz Agric. Res. Station, Dakahlia, Egypt, during the summer season of 2008, to study the effect of inoculation by PSF on mung bean plants cultivated in salinity affected soil (E.C. 5.93) (Table 1).
| Table 1: | Physical and chemical characteristics of experimental soil |
 | |
Phosphate fertilization in the form of calcium superphosphate or RP was added to the soil at 1/3, 2/3 and full-recommended dose, on the base of P2O5 content, before sowing. Mung bean seeds were inoculated by soaking in the spore suspension (2.6x106 spore mL-1) of A. niger, Penicillium sp. or both (A. niger and Penicillium sp., half from each) for 15 min in the presence of 16% Arabic gum as adhesive agent, air dried and then sowed immediately. Seeds treated with spore free solution are considered as control treatment. Treatments were arranged in three replicates. All recommended agricultural practices for mung bean crop were carried out. After two weeks of germination, plants were checked for nodulation.
After 30 and 60 days from sowing, plant and rhizospheric soil samples were collected for determination of numbers and dry weight of nodules plant-1 and counts of total fungi and phosphate solubilizers on PDA and NBRIP agar media, respectively. At the end of cultivation period, plant height (cm), number of branches and pods plant-1, seed index (100-seed weight in gram) and seed yield (kg feddan-1) were recorded. The P content of the seeds was determined by the method of Jackson (1967) after digestion with H2SO4.
Statistical Analysis
The study design was fully randomized blocks. Duncan's new multiple range test were used to compare means. Simple correlation coefficient (r) was performed to examine the relationships between individual properties using the statistical analysis software; CoStat v6.4.
RESULTS AND DISCUSSION
Isolation and Identification of PSF
Fungal isolates with the ability to solubilize insoluble P were isolated from soil characterized with high level of RP. The two isolates that displayed the highest ratio of clear zone/colony diameter were selected and identified as A. niger and Penicillium sp. The zone of P biosolubilization appeared on third day of incubation on NBRIP agar. Continuous observation of the halo zone formation indicates phosphate solubilizing ability which was in increasing order up to the 7th day (Fig. 1). The appearance of a clear halo zone around the colony indicated phosphate solubilization by the fungus (Kang et al., 2002; Gupta et al., 2007). The advantage of using natural phosphate solubilizers over the genetically manipulated or ones that have been isolated from a different environmental sets-up is the easier adaptation and succession when inoculated into the medium containing RP (Xiao et al., 2008).
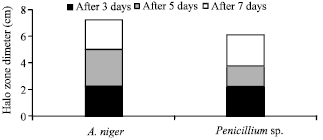 | |
| Fig. 1: | Comparing the efficiency of the selected fungi in releasing P by halo zone formation |
Time Course of RP Biosolubilization
The results (Fig. 2a, b) of the periodic solubilization of RP show that the solubilization of RP was achieved earlier at the third day of incubation on NBRIP broth medium supplemented with RP as a sole P source. Maximum solubilization of RP was observed after 6 and 7 days of incubation by A. niger (67.0 mg L-1) and Penicillium sp. (46.2 mg L-1), respectively. After that, there was no additional solubilization of RP. Maximum P solubilization by A. niger occurred at the end of logarithmic growth phase, whereas, it occurred at the beginning of stationary phase of Penicillium sp. growth.
Vyas et al. (2007) found significant increase with the prolongation of incubation period from 3 to 9 days, followed by a significant decline after 12 day of incubation. Narsian and Patel (2000) reported maximum release of P from China and Udaipur RPs and Sonrai and Hirapur RPs by A. aculeatus after 8 and 14 days of incubation, respectively, they added that variation of time in biosolubilization of RP may have been due to the nature and quantity of organic acids secreted in the medium.
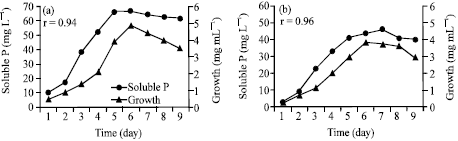 | |
| Fig. 2: | Time course of RP solubilization by the isolated fungi. (a) A. niger and (b) Pencillium sp. |
Data of the analysis of correlation coefficient revealed significant positive correlation between RP solubilization and growth of both A. niger (r = 0.94, p≤0.01) and Penicillium sp. (0.96, p≤0.01). Although, Reddy et al. (2002) mentioned that this correlation is not always showed positive trend.
Biosolubilization of RP at Different Shaking Speeds
Data of the content of soluble P released by the two fungal isolates as affected by different shaking speeds are shown in Fig. 3. The content of soluble P increased with the increase of shaking speed from 60 up to 150 rpm. The maximum contents of soluble P released were 70.5 mg L-1 (A. niger) and 57.2 mg L-1 (Penicillium sp.). Additional shaking speed led to obvious decline in RP solubilization. These results are in consistent with the findings of Xiao et al. (2008), who reported that at the excessive shaking speeds, the growth of the isolates was often weakened by the shear stress resulting from the strong stirring, that was why the released soluble P decreased when the shaking speed increased from 160 to 200 rpm.
At the different shaking speed, decrease in the final pH of the medium of both isolates showed variable values (Fig. 3a, b). As, A. niger recorded slightly lower pHs than that recorded by Penicillium sp. Such decrease in final pH was accompanied by increasing P solubility. The fungus of Penicillium sp. recorded significant correlation between final pH and soluble P (r = -0.92, p≤0.05), whereas, in A. niger, this correlation did not reach to the level of significancy (r = -0.85, ns). Many authors (Cerezine et al., 1988; Vassilev et al., 1995; Vassileva et al., 1998; Barroso and Nahas, 2005) reported reduction in the final culture pH with the increasing of soluble P. However, no significant relationship could be established between the quantities of phosphate solubilized and drop in pH (Narsian and Patel, 2000).
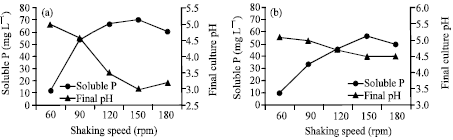 | |
| Fig. 3: | Effect of shaking speed on biosolubilization of RP. (a) A. niger and (b) Penicillium sp. |
Optimum Temperature and pH for Biosolubilization of RP
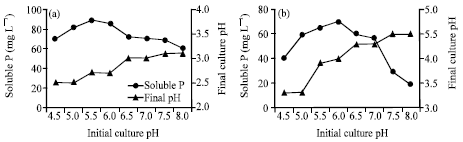
The optimum temperature and initial pH for RP solubilization were investigated as shown in Figures 4 and 5. Temperature and initial pH had significant effects on the biosolubilization of RP (Figures 4a, b and 5a, b).
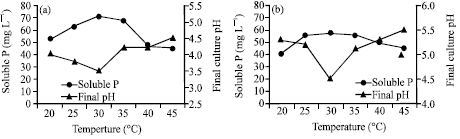 | |
| Fig. 4: | Effect of temperature on the biosolubilization of RP. (a) A. niger and (b) Penicillium sp. |
 | |
| Fig. 5: | Effect of initial culture pH on the biosolubilization of RP. (a) A. niger and (b) Penicillium sp. |
The maximum content of soluble P was recorded at 30°C, which were 70.5 and 57.2 mg L-1 released by A. niger and Penicillium sp., respectively (Fig. 4a, b). Vyas et al. (2007) reported maximum biosolubilization of RP at 36°C. As far as initial pH is concerned, there were differences between the two fungi (Fig. 5a, b). The maximum content of soluble P was recorded at 89.3 mg L-1 released by A. niger at initial pH 5.5, while it was recorded at 70.2 mg L-1 released by Penicillium sp. at initial pH 6.0. Higher or lower than the optimal temperature and initial pH, the content of soluble P decreased. In addition, there was remarkable reduction in final pH for both isolates during the investigations on temperature and initial pHs. The results are in similar to those of Barroso and Nahas (2005) and Xiao et al. (2008).
Biosolubilization of RP as Affected by Carbon Sources
To find out the best source of carbon that can achieve the highest phosphate solubilization, different sources of carbon were added individually in growth medium inoculated with A. niger or Penicillium sp. (Fig. 6). The obtained results showed that glucose was the best followed by fructose and xylose for the two tested fungi in phosphate solubilization. While, the other tested carbon sources showed lower values. According to Cerezine et al. (1988), glucose and fructose are the most frequent and abundant sugars detected in plant exudates that possibly affect the microbial population which solubilizes insoluble phosphates.
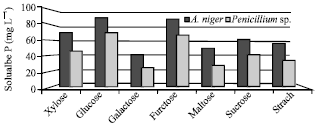 | |
| Fig. 6: | Biosolubilization of RP in the presence of different carbon sources |
Biosolubilization of Different Concentrations of RP
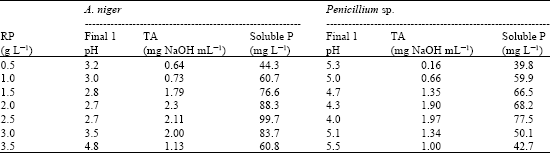
It is clearly obvious from data shown in Table 2 that both A. niger and Penicillium sp. showed positive responses in solubilization efficiency to RP concentration but with various degrees. Generally, a continuous increase in RP solubilization was observed by increasing the concentration of RP added to the growth medium until 2.5 g L-1. This concentration was the optimum in yielding the maximum soluble P. The corresponding values were 99.7 mg L-1 for A. niger and 77.5 mg L-1 for Penicillium sp., above this level of RP (2.5 g L-1) no further increases in solubilization rate was observed.
| Table 2: | Releasing of soluble P by the tested fungi at different RP concentrations |
 | |
| Table 3: | Correlation coefficients between every pairs of RP concentration, final culture pH, TA and soluble P, in culture media of A. niger and Penicillium sp. |
 | |
| ns: Not significant, *p≤0.05, **p≤0.01 | |
The solubilization of RP have been reported to depend on their structure complexity, particle size and quantity of organic acid secreted by microorganisms (Pradhan and Sukla, 2005). Consequently, P is released from mineral phosphate by proton substitution from Ca2+ (Goldstein, 1994), while, Illmer and Schinner (1995) mentioned that RP solubilization is the result of release of protons accompanying respiration or NH4+ assimilation.
At all concentrations of RP, the two tested fungi showed reduction in the final pH, A. niger showed priority in this respect. The titratable acidity (TA) of the supernatant reached its maximum at RP concentration of 2 g L-1 (A. niger) and 2.5 g L-1 (Penicillium sp.). Analysis of correlation coefficient (Table 3) revealed that RP concentration was not related to pH, TA or soluble P, but the later was positively correlated with TA, with respect to pH, results were apparently inconsistent. The results are in accordance with those obtained by Reddy et al. (2002), who reported that phosphate solubilization was not always accompanied by a drop in pH, but always showed the largest production of acids.
Abdel-Hafez (1966), Vassilev et al. (1995), Vassileva et al. (1998) and Barroso and Nahas (2005) explained the biosolubilization of phosphate on the base of acid production. The significant relationship detected between pH and TA shows that the fall in pH may possibly have been the consequence more of acid production than of selective ion absorption by the fungus (Cerezine et al., 1988; Gupta et al., 2007). Indeed, A. niger is characterized by the production of large amount of acids such as oxalic and citric acids (Vassilev et al., 1995; Rashid et al., 2004). Such organic acids have been also, recognized for phosphate solubilization by several species of Penicillium, namely, P. bilaii, P. citrinum, P. janthinellum, P. oxalicum and P. purpurogenum (Cunningham and Kuiack, 1992; Pandey et al., 2008).
Biosolubilization of RP Under Field Simulated Conditions
Aiming to apply the isolated fungi to soil-plant system, the following section was to study their efficiencies in phosphate solubilization in conditions similar to those of field, especially in salinity-affected soils.
RP Solubilization by Isolated Fungi Under NaCl Stress Conditions
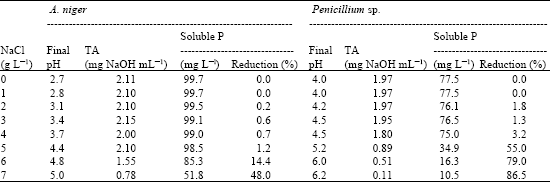
The results of the effect of NaCl stress on solubilization of RP (Table 4) indicate that A. niger solubilized RP under salt stress up to 5%, at this concentration the reduction in soluble P did not exceed 1.2%, however, this fungus was still able to keep 52% of its efficiency at 7% NaCl. Penicillium sp. was less efficient, where; the reduction in soluble P reached 3.2 % at 4% of NaCl and lost 86.5% of its efficiency at 7% NaCl. Pandey et al. (2008) isolated RP solubilizing species of Penicillium tolerated salt concentration up to 20%. On the other side, Kang et al. (2002) observed enhancement of RP solubilization in the presence of 1% NaCl by Fomitopsis sp. PS 102. Higher concentrations of salt are likely to possess mechanisms for survival under stressed environments. Occurrence of such fungal communities may play an important ecological role in low nutrient status and low decomposition rates (Pandy et al., 2008).
Both fungi showed reduction in pH and increasing in TA with the increasing of soluble P. Rajankar et al. (2007) reported that the application of the biofertilizer prepared by fungi may helpful to reduce the salinity of soil by neutralization phenomenon, because these microorganisms release acids in very minute quantity during phosphate solubilization.
| Table 4: | Variation in final culture pH, soluble P and TA by A. niger and Penicillium sp. grown under salt stress conditions |
 | |
Biosolubilization of Common P Sources
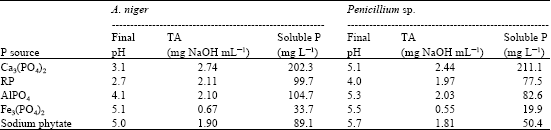
As the initial investigations indicated the potentiality of the two tested fungi in solubilization of RP, it was considered important to extend the study to determine the phosphate dissolving ability of those fungi on some common phosphate forms exist in soil. Data of Table 5 show that, the two isolates were capable of solubilizing all the forms of insoluble P. Solubilization of Ca3(PO4)2 (TCP) was found to be easier than other sources. However, the solubilization levels varied with the two isolates. The released soluble P from the common phosphate sources in descending order was in presence of TCP (202.3 and 211.1 mg L-1), followed by aluminium phosphate (Al-P) (104.7 and 82.6 mg L-1), RP (99.7 and 77.5 mg L-1), sodium phytate (S-P) (89.1 and 50.4 mg L-1) and ferrous phosphate (Fe-P) (33.7 and 19.9 mg L-1) by A. niger and Penicillium sp., respectively. Interestingly, A. niger showed a higher soluble P releasing ability than Penicillium sp. with all the P sources except TCP.
| Table 5: | The efficacy of the tested fungi in solubilization of some common sources of phosphate |
 | |
The microbial solubilization of complex phosphate is not restricted to calcium salts as microorganisms also act upon iron, aluminium and other phosphates (Kang et al., 2002). The majority of phosphate solubilizing microorganisms are able to solubilize calcium-phosphorus complexes but only a few can solubilize iron-phosphorus and aluminium-phosphorus complexes (Kucey et al., 1989). Kang et al. (2002) found that TCP was maximally solubilized by Fomitopsis sp. PS 102 compared with RP and Al-P. In addition, Xiao et al. (2008) concluded that the capability of P solubilization was positively correlated with the grade of P sources, the higher the grade, the higher the content of soluble P released.
The maximum production of acids (Table 5) was observed with TCP (2.74 and 2.44 mg NaOH mL-1) followed by RP (2.11 and 1.97 mg NaOH mL-1) and Al-P (2.1 and 2.3 mg NaOH mL-1), for A. niger and Penicillium sp., respectively. These results are similar to those obtained by Barroso and Nahas (2005), who reported significant positive correlation between the contents of released P and TA.
Antagonism and Biosolubilization of Phosphate Using Dual Culture
The present experiment was initiated to study the interactions between the two tested fungi, aimed at clarifying their antagonistic effect and their solubilization efficiency on different sources of mineral phosphate when applied in a form of dual inoculum. On PDA plates results obtained herein indicated no visible antagonism between A. niger and Penicillium sp. On contrary, a clear compatibility had been confirmed, although, the members of the genus Penicillium are known to be potent producers of antibiotics. Hence, the dual inoculation was applied in solubilization of different P sources.
As illustrated in Fig. 7, the presence of both fungi in the same culture medium increased the efficiency of solubilization of different phosphate sources. The pH and TA of dual inoculation lied within moderate levels between cultures inoculated with individual A. niger or Penicillium sp. as shown previously in Table 5. TCP was the most favorable as P source by the mixed cultures. One very attractive approach for RP solubilization is the application of microorganisms able to excrete low-molecular-mass organic acids, which can strongly increase the concentration of phosphorus by mechanisms involving chelation and exchange reactions (Vassilev et al., 1995; Rashid et al., 2004).
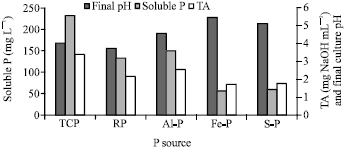 | |
| Fig. 7: | Solubilization of different P sources by mixed culture of A. niger and Penicillium sp. |
Phytase Activity of the Individual and Mixed Fungal Cultures
Data in Fig. 8 show that the tested isolates were active in phytase production, either in individual or in dual culture. Sodium phytate was the most favorable source than the other P forms for phytase activity. Releasing soluble P from different P sources suggest that the enzyme is constitutive and the solubilization process by these isolates is also due to the activity of phytase and acid phosphatase (Achala et al., 2007; Vassilev et al., 2007).
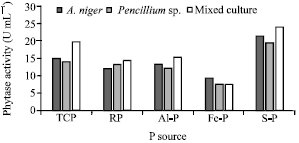 | |
| Fig. 8: | Phytase activity of A. niger, Penicillium sp. and their mixed culture on different P sources |
Application of PSF in Soil-Mung Bean System
Nodulation Status of Mung Bean Plants
Data presented in Table 6 show that inoculation by the two tested fungi in the form of single or dual inoculum significantly increased the number of nodules at all levels of mineral phosphate fertilizer. The stimulation varied however, according to the level of P applied. This was true at early and later stage of growth. The improvement of nodulation was further strengthened, by the dual inoculation, especially, at 2/3 dose of CSP or full dose of RP. This could be observed from checking the differences between those two treatments and uninoculated control after 60 days of planting time. The average increase was about 67.05 and 60.57%, respectively. It is worthy to mention that the dry weight nodular tissues showed similar trend to that observed in case of nodule number. Such results reflect the role of mode of action of the two tested fungi in solubilization efficiency of mineral phosphate through production of a large amount of organic acids in plant rhizosphere, which in return increasing the level of available nutritional elements (Hauka et al., 1996) required at trace level both by the plant and nodule system.
| Table 6: | Nodulation status of mung bean at 30th and 60th day from sowing |
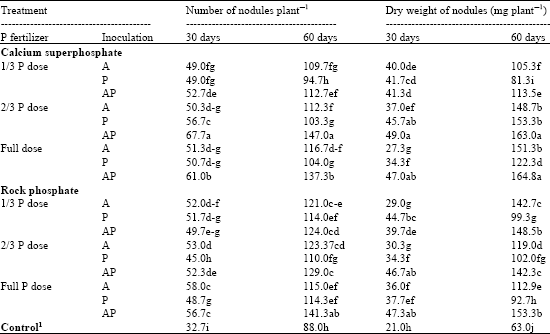 | |
| 1Full dose of recommended phosphate fertilizer (Calcium superphosphate) without inoculation. A: A. niger, P: Penicillium sp., AP: Dual inoculation. Means within the column followed by the same letter(s) are not significantly differed at p≤0.05 | |
Mung Bean Growth and Yield
Concerning the effect of phosphate solubilizing fungi on the growth and yield of mung bean plants under levels of calcium superphosphate (CSP) or RP, data (Table 7) indicated that dual inoculation with A. niger and Penicillium sp. increased significantly plant height in the presence of 2/3 dose of CSP compared to control. Similarly, the dual inoculation showed the same trend in plant height in the presence of full dose of RP. Meanwhile, no significant increase was observed in the inoculation with Penicillium sp. in the presence of 1/3 dose of CSP. In addition, no significant variation among treatments in number of branches per plant. However, the significant effect of the used organisms on plant growth can be attributed not only to nutrient availability (Hauka et al., 1996) but also, to the production of growth substances (Sundaro, 1988).
Concerning the seed yield, the main and final expression of many interacting factors, it could be concluded that the dual inoculation still showed the superiority in achieving seed yield. The beneficial effect of mineral phosphate fertilization in combination with dual inoculation of A. niger and Penicillium sp. could be arranged in descending order (percentage of increase over the control) as follows: 2/3 dose of CSP (18.7%) > full dose CSP (17.8%) > full dose of RP (13.1%) >2/3 dose of RP (10.1%) >1/3 dose of CSP (5.2%) >1/3 dose of RP (3.7%). These results provide sufficient basis to recommend the use of 2/3 or full dose of CSP or RP in combination with dual inoculation with A. niger and Penicillium sp. as successful and proper management of mung bean production.
| Table 7: | Effect of inoculation with phosphate solubilizing fungi on growth and yield of mung bean grown under levels of different phosphate fertilizers |
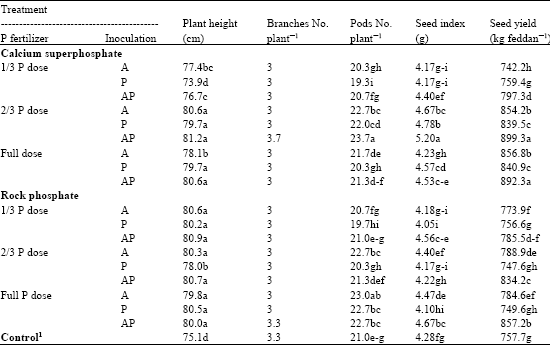 | |
| 1Full dose of recommended phosphate fertilizer (Calcium superphosphate) without inoculation, A: A. niger, P: Penicillium sp., AP: Dual inoculation. Means within the column followed by the same letter(s) are not significantly differed at p≤0.05 | |
Phosphorus Content of Mung Bean
Data of the P content and P uptake of mung bean seeds (Table 8) show that the dual inoculation with 2/3 dose of CSP or RP recorded the highest significantly increment of P content in the seeds of mung bean, compared to control. These treatments also, recorded higher P uptake. These results are inline with the findings of Richa et al. (2007) on maize plants. It is generally accepted that the mechanisms of phosphate solubilization in soils is closely related to the complex forming properties of low molecular weight organic acid (Kucey et al., 1989). Soil microorganisms are deeply involved in this process and their role in solubilization of phosphate bearing materials has been the subject of an interesting number of studies in soil plant system. The attractive approach of microbial mediated solubilization of RP has successfully proved using many filamentous fungi (Vassilev et al., 1995; Reddy et al., 2002). Moreover, Richa et al. (2007) recommended phosphate solubilizing fungi along with RP in alkaline soil (as the soil in the present study), since, the pH of the soil lowered compared to the initial pH. The presence of high P content in mung bean plants may explain the improvement in its growth and yield. Since, P entering in the composition of ATP.
| Table 8: | P content of mung bean as influenced by inoculation with PSF and phosphate fertilization treatments |
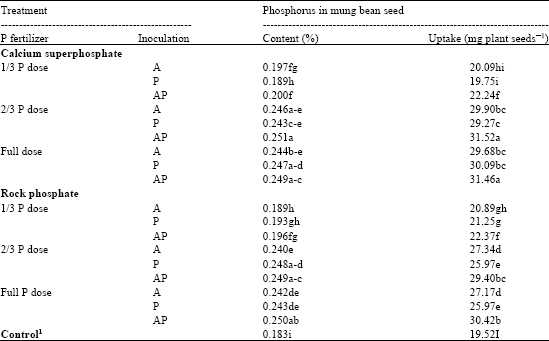 | |
| 1Full dose of recommended phosphate fertilizer (Calcium superphosphate) without inoculation, A: A. niger, P: Penicillium sp., AP: Dual inoculation. Means within the column followed by the same letter(s) are not significantly differed at p≤0.05. | |
Population of Total and Phosphate Dissolving Fungi
Data in Table 9 show the incidence of both total and phosphate dissolving fungi in the rhizospheric soil of mung bean plants as affected by inoculation with A. niger and/or Penicillium sp. Obtained results showed increment in both counts with the increasing of time for all inoculated plants as compared to uninoculated control. These increases in both fungal populations may be due the ability of the isolated fungi to survive under NaCl stress (Table 4) (the experimental soil (Table 1) is saline) and their potential activity in solubilization of different phosphorus forms (Table 5 and Fig. 7) that may commonly exist in the soil, as well as, their phytase activity (Fig. 8) which hydrolysis phytate compounds in the plant residues in the soil.
| Table 9: | Counts of total and phosphate dissolving fungi in the rhizospheric soil of mung bean plants inoculated with A. niger and/or Penicillium sp. |
 | |
| 1Full dose of recommended phosphate fertilizer (Calcium superphosphate) without inoculation. A: A. niger, P: Penicillium sp., AP: Dual inoculation | |
Based on the present data, the amendment of soil with RP along with PSF improves mung bean growth, yield and P-uptake and saves about 1/3 of phosphate fertilizer dose. Thus, it could be recalled that, A. niger and Penicillium sp. could serve as phosphate solubilizers in RP amended soil.
REFERENCES
- Achala, V., V.V. Savantb and M.S. Reddya, 2007. Phosphate solubilization by a wild type strain and UV-induced mutants of Aspergillus tubingensis. Soil Biol. Biochem., 39: 695-699.
CrossRefDirect Link - Barroso, C.B. and E. Nahas, 2005. The status of soil phosphate fractions and the ability of fungi to dissolve hardly soluble phosphates. Applied Soil Ecol., 29: 73-83.
CrossRefDirect Link - Cerezine, P.C., E. Nahas and D.A. Banzatto, 1988. Soluble phosphate accumulation by Aspergillus niger from fluorapatite. Applied Microbiol. Biotechnol., 29: 50l-505.
CrossRefDirect Link - Chuang, C.C., Y.L. Kuo, C.C. Chao and W.L. Chao, 2007. Solubilization of inorganic phosphates and plant growth promotion by Aspergillus niger. Biol. Fert. Soils, 43: 575-584.
CrossRefDirect Link - Cunningham, J.E. and C. Kuiack, 1992. Production of citric and oxalic acids and solubilization of calcium phosphate by Penicillium bilaii. Applied Environ. Microbiol., 58: 1451-1458.
PubMedDirect Link - Domsch, K.H., W. Gams and T.H. Anderson, 1980. Compendium of Soil Fungi. Academic Press, London, United Kingdom, ISBN: 9780122204012, Pages: 1264.
Direct Link - Gupta, N., J. Sabat, R. Parida and D. Kerkatta, 2007. Solubilization of tricalcium phosphate and rock phosphate by microbes isolated from chromite, iron and manganese mines. Acta Botanica Croatica, 66: 197-204.
Direct Link - Hauka, F.I.A., M.M.A. El-Sawah and A.E.I. Selim, 1996. Role of phosphate and silicate solubilizing bacteria in transformation of some macro- and micro-nutrients and their associative effect with Azotobacter on growth and nutrients uptake by plants. Proceedings of the 7th Conference Agronomy, Sept. 9-10, Univ. Mansoura, Fac. Agric., pp: 239-252.
- Illmer, P. and F. Schinner, 1995. Solubilization of inorganic calcium phosphates-solublization mechanisms. Soil Biol. Biochem., 27: 257-263.
CrossRefDirect Link - Kang, S.C., C.G. Ha, T.G. Lee and D.K. Maheshwari, 2002. Solubilization of insoluble inorganic phosphates by a soil-inhabiting fungus Fomitopsis sp. PS 102. Curr. Sci., 82: 439-441.
Direct Link - Khasawneh, F.E. and E.C. Doll, 1979. The use of phosphate rock for direct application to soils. Adv. Agron., 30: 159-206.
Direct Link - Kucey, R.M.N., H.H. Janzen and M.E. Legett, 1989. Microbially mediated increases in plant-available phosphorus. Adv. Agron., 42: 198-228.
Direct Link - Narsian, V. and H.H. Patel, 2000. Aspergillus aculeatus as a rock phosphate solubilizer. Soil Biol. Biochem., 32: 559-565.
CrossRefDirect Link - Nautiyal, C.S., 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett., 170: 265-270.
CrossRefPubMedDirect Link - Pandey, A., N. Das, B. Kumar, K. Rinu and P. Trivedi, 2008. Phosphate solubilization by Penicillium spp. Isolated from soil samples of Indian Himalayan region. World J. Microbiol. Biotechnol., 24: 97-102.
CrossRefDirect Link - Pradhan, N. and L.B. Sukla, 2005. Solubilization of inorganic phosphates by fungi isolated from agriculture soil. Afr. J. Biotechnol., 5: 850-854.
Direct Link - Rajankar, P.N., D.H. Tambekar and S.R. Wate, 2007. Study of phosphate solubilization efficiencies of fungi and bacteria isolated from saline belt of Purna river basin. Res. J. Agric. Biol. Sci., 3: 701-703.
Direct Link - Rashid, M., S. Khalil, N. Ayub, S. Alam and F. Latif, 2004. Organic acids production and phosphate solubilization by Phosphate Solubilizing Microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci., 7: 187-196.
CrossRefDirect Link - Reddy, M.S., S. Kumar, K. Babita and M.S. Reddy, 2002. Biosolubilization of poorly soluble rock phosphates by Aspergillus tubingensis and Aspergillus niger. Bioresour. Technol., 84: 187-189.
CrossRefPubMedDirect Link - Son, H.J., G.T. Park, M.S. Cha and M.S. Heo, 2006. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-42 isolated from soybean rhizosphere. Bioresour. Technol., 97: 204-210.
CrossRefDirect Link - Thalooth, A.T., M.M. Tawfik and M.H. Mohamed, 2006. A comparative study on the effect of foliar application of zinc, potassium and magnesium on growth, yield and some chemical constituents of mungbean plants grown under water stress conditions. World J. Agric. Sci., 2: 37-46.
Direct Link - Vassilev, N., M.T. Baca, M. Vassileva, I. Franco and R. Azcon, 1995. Rock phosphate solubilization by Aspergillus niger grown on sugar-beet wastemedium. Applied Microbiol. Biotechnol., 44: 546-549.
CrossRefDirect Link - Vassilev, N., M. Vassileva, V. Bravo, M.F. Serrano and I. Nikolaeva, 2007. Simultaneous phytase production and rock phosphate solubilization. by Aspergillus niger grown on dry olive wastes. Ind. Crops Prod., 26: 332-336.
CrossRefDirect Link - Vassileva, M., N. Vassilev and R. Azcon, 1998. Rock phosphate solubilization by Aspergillus niger on olive cake-based medium and its further application in a soil-plant system. World. J. Microbio. Biotechnol., 14: 281-284.
CrossRefDirect Link - Vyas, P., P. Rahi, A. Chauhan and A. Gulati, 2007. Phosphate solubilization potential and stress tolerance of Eupenicillium parvum from tea soil. Mycological Res., 111: 931-938.
CrossRefDirect Link - Xiao, C.Q., R.A. Chi, X.H. Huang, W.X. Zhang, G.Z. Qiu and D.Z. Wang, 2008. Optimization for rock phosphate solubilization by phosphate-solubilizing fungi isolated from phosphate mines. Ecol. Eng., 33: 187-193.
CrossRef