
C. Nwinyi Obinna
1Department of Biological Sciences, College of Science and Technology, Covenant University, KM 10 Idiroko Road, Canaan Land, P.M.B. 1023, Ota, Ogun State, Nigeria
Nwodo-Chinedu Shalom
1Department of Biological Sciences, College of Science and Technology, Covenant University, KM 10 Idiroko Road, Canaan Land, P.M.B. 1023, Ota, Ogun State, Nigeria
O. Amund Olukayode
2Department of Botany and Microbiology, Faculty of Science, University of Lagos, Akoka-Yaba, Lagos, Lagos State, Nigeria
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 99-104







ABSTRACT
Two closely related strains of Rhodococcus species, isolated from aniline polluted tropical ecosystem, were able to utilize aniline as carbon source in 3.0 and 4.0 mM concentrations at 30°C and pH of about 6.4. Rapid increase in turbidity and a sharp decline in pH were observed in the cultures of both organisms within 24 h of incubation. Shortly after the period, growth became slower. Turbidity values obtained at 4.0 mM concentrations of aniline was about twice the values obtained at 3.0 mM concentrations. Aniline concentrations of 10 mM and above were found to be toxic for the organisms. Rhodococcus species because of its significant prevalence in agricultural soils can be used as an effective means of recovering tropical agricultural land polluted with aniline, aniline-based herbicides or its derivatives.
PDF Abstract XML References Citation
How to cite this article
C. Nwinyi Obinna, Nwodo-Chinedu Shalom and O. Amund Olukayode, 2008. Biodegradation Potential of Two Rhodococcus Strains Capable of Utilizing Aniline as Carbon Source in a Tropical Ecosystem. Research Journal of Microbiology, 3: 99-104.
URL: https://scialert.net/abstract/?doi=jm.2008.99.104
URL: https://scialert.net/abstract/?doi=jm.2008.99.104
INTRODUCTION
Aniline is a widely distributed environmental pollutant released into the environment from industrial processes, primarily in the effluents of dye manufacturing (Meyer, 1981) and agricultural chemicals (Kearney and Kaufmann, 1975). According to Neilson (1994), aniline is used in the manufacture of aromatic amide, dyes and pharmaceuticals. Aniline is released mainly during the course of its use as chemical intermediate in the production of polymers, pharmaceuticals and dyes (USEPA, 1985).
They are ubiquitous in the environment and are formed as the initial microbial transformation products from a number of agrochemicals such as carbamates, urea and aromatic amide. Aniline in solution adsorbs to colloidal organic matter which effectively increase its solubility and movement into ground water. Aniline (Pka of 4.596) is also moderately adsorbed to organic matter in the soil and adsorption is dependent upon soil pH (Howard, 1989). It slowly volatilizes from soil and surface water and is biodegradable. Although rapidly degraded in the atmosphere by a series of photochemical reactions (USEPA, 1985), aniline can be deposited in the soil by wet and dry deposition and by adsorption on aerosol particles. The primary toxic effect resulting from acute exposure to aniline, by inhalation or by oral route, is methemoglobinemia, usually accompanied with anoxia, erythrocyte damage and spleen effects (USEPA, 1985). Aniline bound to humic materials in the soil is subject to oxidation (Howard, 1989). Photo degradation of aniline on the soil surface is also thought to be an environmentally important removal process (USEPA 1985). The combination of these processes eventually results in the degradation of aniline to carbondioxide (CO2).
Soil microorganisms had shown that through acclimatization or mutation they alter their metabolic pathways for a more efficient utilization of synthetic compounds. Microbial enzymes often have a surprisingly broad specificity and in addition to their natural substrates will degrade with greater or lesser efficiency numerous analogs and sometimes even structurally quite unrelated compounds (Bollen, 1961). Microbial transformation and degradation are major mechanisms to eliminate aniline from the environment. Degradation of 44.2% of the incubated aniline to CO2 in 10 days and 12% in 20 days, respectively by different isolated soil microorganism have been demonstrated in the laboratory (USEPA, 1985). A number of microorganisms in soil can use aniline as a sole carbon and nitrogen source. Several microorganisms in particular bacteria have been implicated in the utilization of aniline. Bacterial species of Pseudomonas (Hinteregger et al., 1992), Comamonas (Parales et al., 1997) Acinetobacter and Rhodococcus (Aoki et al., 1983), Frateuria (Murakumi et al., 1999) and Norcadia (Bachofer et al., 1975) and Delftia sp. AN 3 ( Liu et al., 2002 ) have been shown to be able to degrade aniline or its derivatives. Products apparently formed from its oxidation include azobenzene, phenazine, formalin and acetanilide (USEPA, 1985).
In the present study, two related strains of aniline-utilizing bacteria from the Nigerian soil, a tropical ecosystem, were isolated and characterized. This is aimed at determining the biological fate of aniline in the tropical ecosystem.
MATERIALS AND METHODS
Chemicals and Reagents
All chemicals and reagents were of analytical grade. Aniline was obtained from Merck, Germany. All other chemicals and reagents were obtained from Sigma Chemicals Co. Ltd., England.
Selection and Preparation of Site
The study site was within the Akoka campus of the University of Lagos. The site had physical characteristics of a loamy soil with high moisture content (55%) which is ideal for agricultural farming. Twenty milliliters of aniline was soaked with a cotton wool and transferred to a beaker wrapped with aluminum foil and autoclaved at 121°C for 15 min. This was cooled and buried, about 12 cm deep, in the tropical soil. The aniline soaked cotton wool was exhumed after 7 days and aseptically transferred into 1 L of a sterilized enrichment medium which consists of (g L-1): K2HPO4, 7. 0; KH2PO4, 3.0; MgSO4.7H2O, 0.l, yeast extract, 0.05 and Aniline (Carbon source) 1.2.0 (Liu et al., 2002). The pH was adjusted to 6.5.
Isolation of Aniline-Degrading Bacteria
Aniline-degrading bacteria were isolated from the distinct bacterial colonies obtained by plating out 0.1 mL aliquots of serial dilutions of the enrichment medium containing the bacteria on sterile aniline agar plates. The aniline agar contained (g L-1): K2HPO4, 7.0; KH2PO4, 3.0, MgSO4.7H2O, 0.1; yeast extract, 0.05; Aniline, 1.2 and Agar, 12.5 (Liu et al., 2002). Pure isolates were obtained by sub-culturing on the aniline agar.
Identification of the Bacterial Isolates
The pure bacterial isolates, strains A-deg 1 and A-deg 2, were identified on the basis of their morphological features and biochemical tests (Cowan, 1985; Olutiola et al., 1991).
Growth Profiles of the Isolates
The isolates were cultured in the enrichment medium containing 3.0 or 4.0 mM aniline as sole carbon source and placed in 200 mL conical flask and incubated on rotary shaker at 30°C for 21 days. Growth was monitored by measuring the turbidity at 540 nm using Biomedical Colorimeter (USA) and pH using a digital pH meter.
RESULTS
Two closely related strains of bacteria capable of degrading aniline (A-deg 1 and A-deg 2) were isolated from Lagos, Nigeria. The isolates of aniline degraders were identified as two different strains of Rhodococcus species by comparing their morphological and biochemical characteristics with standard reference organisms (Cowan, 1985; Olutiola, 1991). The two strains of the organism differed only in the area of nitrate and maltose utilization. The strain of Rhodococcus species (A-deg 1) gave positive growth in the nitrate broth and no growth in maltose medium whereas the other strain (A-deg 2) yielded no growth in the nitrate broth and significant growth in maltose containing medium (Table 1).
The growth studies showed that both organisms (A-deg 1 and A-deg 2) can grow in the minimal medium containing aniline as sole carbon sources. Rapid increase in turbidity and a sharp decline in pH were observed in the cultures of both organisms within 24 h of incubation. Shortly after the period, growth became slower. Higher turbidity was obtained at 4.0 mM concentrations of aniline, about 2-times the values obtained at 3.0 mM concentrations. More growth was obtained at 4.0 mM. When the organism was cultured in medium containing 3.0 mM aniline, the O.D. rose to 0.07 at day 1 and subsequently to 0.082 at day 21 whereas the pH declined from 6.54 to 6.34 within the 21-day period. At 4.0 mM aniline, the OD was 0.135 at day 1 and 0.167 at day 21 while the pH declined from 6.54 to 6.37 within the same period (Fig. 1a and b). Higher turbidity was observed at 4.0 mM aniline concentrations compared to 3.0 mM concentrations. At 3.0 mM concentration, the OD was 0.06 at day 1 and 0.08 at day 21; the pH declined from 6.54 to 6.33 after 12 days of incubation. The OD was 0.13 at day 1 and 0.16 at day 21 whereas the pH decreased from 6.54 to 6.34 within the 21-day period of incubation (Fig. 2).
| Table 1: | Morphological and biochemical characteristics of bacteria isolates capable of degrading aniline |
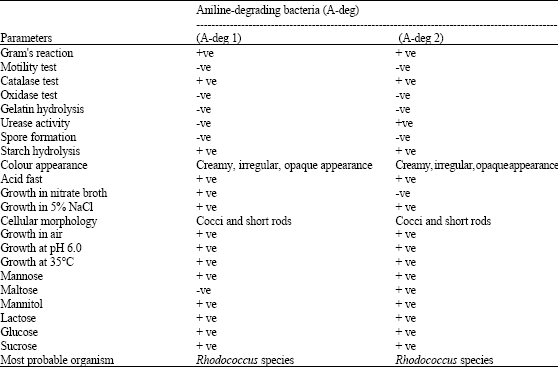 | |
| +ve = Growth; -ve = No growth | |
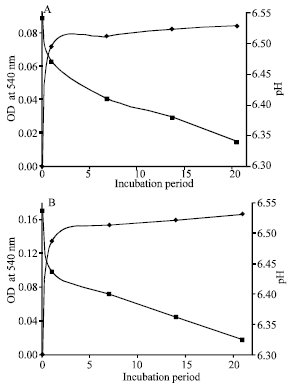 | |
| Fig. 1: | Growth profile [Absorbance (♦), pH (#)] of Aniline-degrading Bacteria (A-deg 1) incubated at 30°C in (A) 3.0 mM and (B) 4.0 mM concentrations of aniline for 1-21 days |
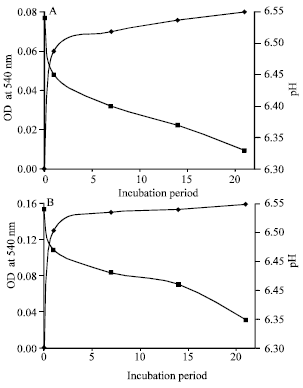 | |
| Fig. 2: | Growth profile [Absorbance (♦), pH (#)] of Aniline-degrading Bacteria (A-deg 2) incubated at 30°C in (A) 3.0 mM and (B) 4.0 mM concentrations of aniline for 1-21 days |
DISCUSSION
In this study, two wild strains of Rhodococcus species were isolated from Nigerian tropical ecosystem with the demonstration of ability to thrive in minimal broth containing 3.0 and 4.0 mM concentration aniline as sole carbon sources. Aniline-degrading Rhodococcus species had been isolated in temperate ecosystem (Aoki et al., 1983). A thorough look at the pH and optical density (turbidity) readings of the growth profile of the Rhodococcus species indicated a similar trend at 3.0 or 4 mM concentrations. As the pH was decreasing, the turbidity increases. This is in consonance with single-substrate enzyme-catalyzed reactions. In terms of tolerance to aniline, no growth of the strains Rhodococcus species was obtained after 10.0 mM concentrations of aniline. This means that higher levels of aniline were toxic to the organisms. This is lower than levels reported for some aniline degrading organisms. Pseudomonas sp. which thrives in concentrations of up to 32.0 mM aniline (Konopka et al., 1989) and Delftia sp. AN3 growing in concentrations of up to 53.8 mM aniline have been described (Liu et al., 2002). This is in view of the fact that the isolates unlike the Pseudomonas sp. and Delftia sp. are Gram-positive bacteria. Gram-positive bacteria are usually more sensitive than Gram-negative ones towards lipophilic toxic substrates, possibly because they lack protection by the outer membrane (Prescott et al., 2002).
From studies on bacterial growth kinetics, the time interval involved in the lag and log phases of growth are very short. This could be due to acclimatization and adaptation of the organisms to aniline during the selection stage. As the concentration of aniline in the minimal broth was increased from 3.0 to 4.0 mM, there appeared to be a corresponding increase or induction of the enzymes involved in the degradation of aniline. This proposes that in the presence of aniline certain enzymes involved for the degradation are produced. The decrease in pH with increasing turbidity is in agreement with the general knowledge about microbial degradation, particularly in batch fermentation. Liu et al. (2002) proposed a pathway for aniline degradation in Delftia sp. AN3 leading to the production of acidic intermediates such as 4-oxalocrotonic acid, 4-hydroxy-2-oxovaleric acid and pyruvic acid. Production of such acidic compounds in the culture medium could account for the decline in pH. According to Wilson et al. (1983) and Swindoll et al. (1988a), subsurface bacteria are known to degrade aniline at concentrations below 0.1 mg L-1, but mineralization often represents less than 10% of the total metabolism while the remaining is incorporated into biomass. Hence, the increase in turbidity signifies growth which invariably means utilization of the aniline.
Rhodococcus species are prevalent in soils. This makes it a good choice for use in bioremediation. And above all, it is safe and cost-effective because of the competitive advantage over non-indigenous or foreign organisms within our environment.
REFERENCES
- Aoki, K., K. Ohtsuka, R. Shinke and H. Nishira, 1983. Isolation of aniline-assimilating bacteria and physiological characterization of aniline biodegradation in Rhodococcus erythropolis AN-13. Agric. Biol. Chem., 47: 2569-2575.
CrossRef - Bachofer, R., F. Lingens and W. Schafer, 1975. Conversion of aniline into pyrocatechol by a Nocardia sp.; incorporation of oxygen-18. FEBS Lett., 50: 288-290.
CrossRefDirect Link - Bollen, W.B., 1961. Interactions between pesticides and soil microorganisms. Ann. Rev. Microbiol., 15: 69-92.
CrossRef - Hinteregger, C., L.M. Loid and F. Streichsbier, 1992. Characterization of isofunctional ring-cleaving enzymes in aniline and 3-chloroaniline degradation by Pseudomonas acidovorans CA28. FEMS Microbiol. Lett., 97: 261-266.
CrossRefDirect Link - Konopka, A., D. Knight and R.F. Turco, 1989. Characterization of a Pseudomonas sp. capable of aniline degradation in the presence of secondary carbon sources. Applied Environ. Microbiol., 55: 385-389.
Direct Link - Liu, Z., H. Yang, Z. Huang, P. Zhou and S.J. Liu, 2002. Degradation of aniline by newly isolated, extremely aniline-tolerant Delftia sp. AN3. Applied Microbiol. Biotechnol., 58: 679-682.
Direct Link - Murakami, S., A. Takashima, J. Takemoto, S. Takenaka, R. Shinke and K. Aoki, 1999. Cloning and sequence analysis of two catechol-degrading gene clusters from the aniline-assimilating bacterium Frateuria species ANA-18. Gene, 226: 189-198.
CrossRefDirect Link - Parales, E.E., T.A. Ontl and D.T. Gibson, 1997. Cloning and sequence analysis of a catechol 2,3-dioxygenase gene from the nitrobenzene-degrading strain Comamonas sp JS765. J. Indian Microbiol. Biotechnol., 19: 385-391.
CrossRefDirect Link - Swindoll, C.M., C.M. Aelion, D.C. Dobbins, O. Jiang, S.C. Long and F.K. Pfaender, 1988. Aerobic biodegradation of natural and xenobiotic organic compounds by subsurface microbial communities. Environ. Toxicol. Chem., 7: 291-299.
CrossRefDirect Link - Wilson, J.T., M.C.J.F. Nabb, B.H. Wilson and M.J. Noonan, 1993. Biotransformation of selected organic pollutants in ground water. Dev. Ind. Microbiol., 24: 225-233.
Direct Link