
Zamberi Sekawi
Department of Medical Microbiology and Parasitology,Faculty of Medicine and Health Sciences,Universiti Putra Malaysia, 43400 UPM Serdang, Selangor DE, Malaysia
Rusmah Yusof
Department of Medical Microbiology and Parasitology,Faculty of Medicine and Health Sciences,Universiti Putra Malaysia, 43400 UPM Serdang, Selangor DE, Malaysia
Mariana Nor Shamsudin
Department of Medical Microbiology and Parasitology,Faculty of Medicine and Health Sciences,Universiti Putra Malaysia, 43400 UPM Serdang, Selangor DE, Malaysia
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 489-493







ABSTRACT
A study was conducted to portray a preliminary characteristic of extended-spectrum β-lactamases (ESBLs)-producing Escherichia coli in a local tertiary hospital in Malaysia. Sixteen clinical isolates of ESBLs producing E. coli from different sources were examined for blaSHV/TEM/CTX-M ESBL genes by PCR molecular assay. Each strain was found to carry at least one of the genes. This study demonstrated a high prevalence of blaCTX-M (81.3%) and blaTEM genes (75%). Only two strains (12.5%) carried the blaSHV gene. Nucleotide and deduced protein sequences determination showed; 61% produced CTX-M-15, 31% produced CTX-M-14 and 8% produced CTX-M-3. The antimicrobial susceptibility testing data determined that almost all sixteen isolates were resistant to oxyimino-cephalosporins, 46% resistant to gentamicin, 69% resistant to trimethoprim-sulfamethoxazole and 46% resistant to ciprofloxacin. This study preliminary emphasizes the epidemiology of the ESBL-producing E. coli particularly SHV, TEM and CTX-M-type producing E. coli in Malaysia.
PDF Abstract XML References Citation
How to cite this article
Zamberi Sekawi, Rusmah Yusof and Mariana Nor Shamsudin, 2008. Extended-Spectrum β-Lactamases-Producing Escherichia colifrom a Tertiary Hospital in Malaysia: Emergence of CTX-M-Type β-Lactamases Variation. Research Journal of Microbiology, 3: 489-493.
URL: https://scialert.net/abstract/?doi=jm.2008.489.493
URL: https://scialert.net/abstract/?doi=jm.2008.489.493
INTRODUCTION
The increasing resistance of gram negative bacteria is of worldwide concern. Its rapid evolution to become resistant is an immense challenge to clinicians, scientists and researchers worldwide. Despite many new β-lactam antibiotics being developed over the last 20 years, new β-lactamases emerged, causing them to be resistant to these antibiotics (Bradford, 2001). This is presumably due to overuse of new antibiotics resulting in selective pressure to select new variants of β-lactamases. Its infection has now become even more difficult to treat as its resistance level has increased over the years. Now with the threat of the so-called ESBL-producing pathogens, it looks like we are fighting a losing battle.
Extended spectrum β-lactamase or ESBL is a β-lactamase enzyme with increased spectrum of activity especially against the oxyimino-cephalosporins such as cefotaxime and ceftazidime. Most ESBLs are derivatives of TEM or SHV enzymes (Bush et al., 1995). There are now >150 TEM-type β-lactamases and >100 SHV-type enzymes (www.lahey.org/studies/webt.htm). They are easily transmissible from one organism to another. Hence, SHV-and TEM-types which have been recognized since the 1980s are now a major cause of nosocomial infections all over the world.
In Malaysia, a national surveillance on antibiotic resistance was conducted in 2005 and found that ceftazidime and cefotaxime resistance in E. coli was 8.9% (0.9% from the community) and 10.1% (1.0% from the community) respectively (unpublished data). Tests for ESBL production was unfortunately not performed routinely in this survey. Another study done in a university hospital in Malaysia reported blaSHV-5 to be dominant in ESBL-producing E. coli and was postulated to have been acquired from ESBL-producing Klebsiella pneumoniae via plasmid transfer (Subramaniam et al., 2006).
Another family of plasmid-mediated ESBLs which are non-TEM and non-SHV derivatives, called CTX-M has emerged. They preferentially hydrolyze cefotaxime and have mainly found in wide range of bacteria, especially members of the Enterobacteriaceae family (Bonnet, 2004). Strains expressing CTX-M-type β-lactamases have been isolated from many parts of the world. Based on communications with various experts in many local centres, CTX-M-type is also available in Malaysia and it poses a potential threat in healthcare management. Realizing the lack of published local research on this issue, the aim of this study is to demonstrate the emergence of CTX-M-type in ESBL-producing E. coli in a tertiary hospital in Malaysia.
MATERIALS AND METHODS
Bacterial Strains
Sixteen non-duplicate isolates of validated ESBL-producing E. coli from multiple types of specimens collected from a tertiary hospital in Malaysia in 2006 were analyzed in this study.
β-Lactamase Gene Identification
Chromosomal DNA was prepared from an overnight bacterial culture (18-24 h) on a MacConkey agar plate. Approximately ten colonies were re-suspended into 50μL sterile distilled water and the cells were lysed by boiling for 15 min, then immediately cooled on ice for another 2 min. Cellular debris were removed by centrifugation at 10,000 g for 1 min and the supernatant was used as a source of template DNA for amplification of blaTEM and blaSHV genes.
Plasmid DNA was prepared by using a commercial kit (Invitrogen, BioDiagnostic, USA). The plasmid DNA was used as a source of template DNA for amplification of blaCTX-M gene.
PCR primers used for amplification of CTX-M types were universal primers of CTX-MA1 (5`-SCS ATG TGC AGY ACC AGT AA-3`) and CTX-MA2 (5`-CCG CRA TAT GRT TGG TG-3`) described by Saladin et al. (2002) yielding a 544 bp amplicon (S = G or C; Y = C or T; R = A or G). The primers used for PCR amplification of blaTEM and blaSHV were mentioned previously by Perilli et al. (2002) yielding amplicons of 1080 and 1050 bp in size, respectively.
PCR Amplification
The cycling condition for the amplification of blaTEM and blaSHV were described previously by Perilli et al. (2002) and the DNA amplification steps for blaCTX-M were as follows: initial denaturation at 94°C for 4 min followed by 30 cycles of denaturation at 95°C for 30 sec, annealing at 57.6°C for 1 min and elongation at 72°C for 1 min with final extension at 72°C for 4 min. All reactions were performed in a 25μL volume using 6 units of Taq DNA polymerase (Bioron, Germany), 2.5μL of 10x PCR buffer, 2 mM MgCl2, 0.2 mM deoxynucleoside triphosphates, 0.3μL of 10 pmol mL-1 primers each and 1μL DNA template. The PCR reactions were performed using PCR Thermal Cycler (Biometra-TPersonal, Germany). All negative reactions were repeated twice.
DNA Sequencing
All purified PCR product were sent for commercial sequencing (1st Base Laboratories Sdn. Bhd., Malaysia). The sequences were edited and analyzed by using DNAsis for Windows 2.6 (Hitachi Software Engineering, USA). The nucleotide and the deduced protein sequences were analyzed by using the software available online at the National Center for Biotechnology Information website (http://www.ncbi.nlm.nih.gov).
Control Strains
The quality control strains used were E. coli ATCC 25922 (sensitive strain), K. pneumoniae ATCC 700603 (blaSHV-18), E. coli 13-00 (blaCTX-M-14), E. coli 78-01(blaCTX-M-15) and E. coli 188-04 (blaCTX-M-2).
RESULTS AND DISCUSSION
Sixteen clinical isolates of ESBLs-producing E. coli carried at least one of the blaCTX-M, blaTEM, or blaSHV ESBL genes (Table 1). A vast majority of the strains carried blaCTX-M (81.3%) and blaTEM genes (75%). Only two strains (12.5%) carried the blaSHV gene. There were at least three circulating strains of which CTX-M-15 being the most dominant (61%), followed by CTX-M-14 (31%) and CTX-M-3 (8%).
Almost all strains demonstrated resistance to the second, third and fourth generation of cephalosporins (Table 2). There were more resistance to cefotaxime than ceftazidime among CTX-M-types, phenotypically supporting the presence of CTX-M enzymes.
It is clearly shown that at least in this study, CTX-M-types have been the dominant strains. This is in parallel to the worldwide trend where CTX-M-types have overtaken dominance of TEM-and SHV-types globally. Though prior data on the prevalence of TEM-types and SHV-types are not known in Malaysia to demonstrate this fact, we believe similar trend is valid. It is difficult to determine the proportion of our strains which originated from the community as these patients were admitted to the hospital where medical records access is restricted owing to confidentiality and security issues.
In our study, we found at least three circulating strains of which CTX-M-15 being the most dominant (61%), followed by CTX-M-14 (31%) and CTX-M-3 (8%). CTX-M-15 and CTX-M-3 belong to the CTX-M-1 group while CTX-M-14 to the CTX-M-9 group (Bush et al., 1995). Based on this finding, we agree to the hypothesis that states the current spread of CTX-M enzymes, is allodemic in nature, rather than an epidemic pattern, reflecting that the increase of CTX-M enzymes is a result of both multiple specific clones and/ or mobile genetic elements and not dissemination of a particular clone (Baquero et al., 2002). Owing to the dynamic nature of these clones, it is not surprising if the dominant strains would change over time (Bonnet, 2004).
| Table 1: | The β-lactamase types of ESBL-producing Escherichia coli |
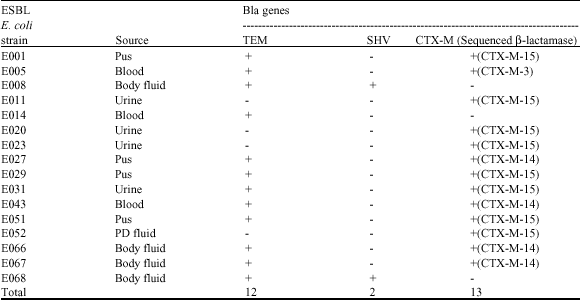 | |
| +: Present; -: Absent | |
| Table 2: | Antibiotic resistant pattern of sixteen ESBL-producing Escherichia coli |
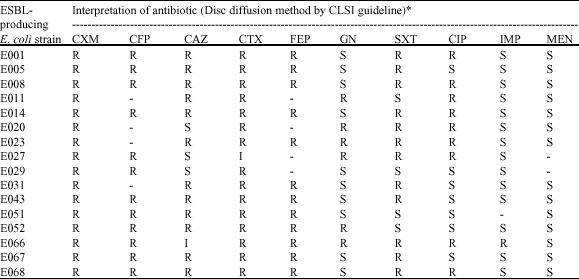 | |
| *CXM: Cefuroxime; CFP: Ceforperazone; CAZ: Ceftazidime; CTX: Cefotaxime; FEP: Cefepime; GN: Gentamicin; SXT: Trimethoprim-sulfamethoxazole; CIP: Ciprofloxacine; IMP: Imipenem; MEM: Meropenem; S: Susceptible; R: Resistance; I: Intermediate; -: Absent | |
Several studies have shown an increasing association with resistance to other classes of antimicrobial agents among ESBL-producing organisms isolated form the community (Pitout et al., 2005). This includes co-resistant with trimethoprim-sulfamethoxazole, tetracycline, gentamicin and ciprofloxacin. This association may be evident in this study as 46% of the strain was resistant to gentamicin, 69% resistant to trimethoprim-sulfamethoxazole and 46% resistant to ciprofloxacin.
In order to further understand the role of ESBL to healthcare, more extensive studies will be conducted. These include linking CTX-M-types with E. coli phylogenetic group. We certainly would like to know this association with our strains, especially the dominant strains. CTX-M-types have been reported to be associated with groups D and B2, both groups of which are mainly associated with extraintestinal infections (Cantun and Conque, 2006). Virulence markers in our local CTX-M-producing E. coli would be investigated. It was reported that there were significant differences from individual virulence factors among CTX-M producers and non-producers; however, aggregate virulence factor scores were similar (Pitout et al., 2007).
A comprehensive surveillance study is in the pipeline which, in our opinion will provide a good overview and more accurate picture of the problem of ESBL in our country.
The increasing evidence of CTX-M-producing bacteria in food-producing bacteria such as cattle and poultry and pets, namely, dogs and cats is worrying (Pitout et al., 2005). This may acts as reservoir for the acquisition of resistant organism which will make the quest for winning this ‘battle’ an uphill task. It is therefore, necessary for us to study this situation in our region.
In conclusion, ESBL is indeed a significant problem in our clinical settings in this tertiary hospital and therefore, it is not incorrect to postulate this problem as also a national issue. We have succeeded in determining the group type of the enzymes and in this preliminary study, the epidemiology of the enzymes seem to give a slightly clearer picture of the impact of ESBL in Malaysia. However, it is still premature to draw conclusions based on the existing data.
ACKNOWLEDGMENTS
This study was funded by Intensified Research in Priority Areas Grant No. 54436 and 5523248 from the Ministry of Science, Technology and Innovation of Malaysia. Our sincere appreciation to the Ministry of Science, Technology and Innovation of Malaysia for the research grants and Dr. Johann Pitout for the control strains E. coli 13-00 (blaCTX-M-14), E. coli 78-01(blaCTX-M-15) and E. coli 188-04 (blaCTX-M-2).
REFERENCES
- Baquero, F., T.M. Coque and R. Cantón, 2002. Allodemics. Lancet Infect. Dis., 2: 591-592.
Direct Link - Bonnet, R., 2004. Growing group of extended-spectrum β-lactamases: The CTX-M enzymes. Antimicrob. Agents Chemother., 48: 1-14.
CrossRefDirect Link - Bradford, P.A., 2001. Extended-spectrum-lactamases in the 21st century: Characterization, epidemiology and detection of this important resistance. Threat. Clin. Microbiol. Rev., 14: 933-951.
Direct Link - Bush, K., G.A. Jacoby and Medeiros, 1995. A functional classification scheme for beta-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother., 39: 1211-1233.
Direct Link - Cantün, R. and T.M. Conque, 2006. The CTX-M-lactamase pandemic. Curr. Opin. Microbiol., 9: 466-475.
PubMed - Perilli, M., E. Dell’Amico, B. Segatore, M.R. De Massis and C. Bianchi et al., 2002. Molecular characterization of extended-spectrum-lactamases produced by nosocomial isolates of enterobacteriacae from an Italian nationwide survey. J. Clin. Microbiol., 40: 611-614.
Direct Link - Pitout, J.D., K.B. Laupland, D.L. Church, M.L. Menard and J.R. Johnson, 2005. Virulence factors of Escherichia coli isolates that produce CTX-M-type extended-spectrum-lactamases. Antimicrob. Agents Chemother., 49: 4667-4670.
Direct Link - Pitout, J.D., D.L. Church, D.B. Gregson, B.L. Chow, M. McCracken, M.R. Mulvey and K.B. Laupland, 2007. Molecular Epidemiology of CTX-M-Producing Escherichia coli in the Calgary Health Region: Emergence of CTX-M-15-Producing Isolates. Antimicrobial. Agents Chemother., 51: 1281-1286.
CrossRef - Saladin, M., V.T. Bao Cao, T. Lambert, J.L. Donay and J.L. Herrmann et al., 2002. Diversity of CTX-M-lactamases and their promoter regions from Enterobacteriacae isolated in the three Parisian hospitals. FEMS. Microbiol. Lett., 209: 161-168.
Direct Link