
Swati Sucharita Dash
Applied and Industrial Microbiology Laboratory, Department of Biotechnology, Indian Institute of Technology, Madras, Chennai, India
Sathyanarayana N. Gummadi
Applied and Industrial Microbiology Laboratory, Department of Biotechnology, Indian Institute of Technology, Madras, Chennai, India
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 457-465







ABSTRACT
The study was undertaken to investigate the effect of caffeine (1, 3, 7- trimethylxanthine) on growth, morphology and viability of caffeine degrading Pseudomonas sp. and other non caffeine degrading bacterial strains. Growth, morphology and cell viability of the bacterial strains were studied in caffeine medium, minimal medium without caffeine and in minimal medium with caffeine added at log phase of growth. The caffeine degrading Pseudomonas sp. achieved a maximum cell dry weight of 1.1 g L-1 after caffeine addition at log phase of growth without any change in morphology. The growth and viability of E. coli DH5α and other bacterial strains was greatly reduced upon addition of caffeine at log phase of growth. E. coli DH5α strain formed long filamentous cells on caffeine exposure and lysis occurred for other Gram negative bacterial species and Bacillus subtilis. E. coli DH5α transformed with plasmid from caffeine degrading Pseudomonas sp. was found to be tolerant to caffeine. This study shows the susceptibility of non caffeine degrading bacterial strains to caffeine and will be helpful in understanding the basics of the evolution and survival of xenobiotic degrading strains in nature.
PDF Abstract XML References Citation
How to cite this article
Swati Sucharita Dash and Sathyanarayana N. Gummadi, 2008. Inhibitory Effect of Caffeine on Growth of Various Bacterial Strains. Research Journal of Microbiology, 3: 457-465.
URL: https://scialert.net/abstract/?doi=jm.2008.457.465
URL: https://scialert.net/abstract/?doi=jm.2008.457.465
INTRODUCTION
Caffeine (1, 3, 7 trimethylxanthine) is a widely used stimulant of the central nervous system and is consumed worldwide in form of beverages and pharmaceutical preparations. It is known to exert numerous physiological effects on different organisms at micro molar concentrations. The most significant effects being inhibition of phosphodiesterase resulting in increase in intracellular cAMP levels; effect on intracellular calcium levels and antagonism of adenosine receptors (Serafin, 1995). Apart from that, caffeine has also been reported to be an antimicrobial agent most effective against E. coli (Ramanaviciene et al., 2003) and this is attributed to the effect of caffeine on DNA and protein synthesis in E. coli. Reports also indicate that caffeine enhances the inhibitory effect of certain antibacterial agents like penicillin and tetracycline against Staphylococcus aureus and of furazolidone against vibrios (Banerjee and Chatterjee, 1981).
However numerous bacterial species have been known to degrade this compound and use it as a source of nutrient for growth, the most prevalent genus being Pseudomonas (Dash and Gummadi, 2006a). It is therefore interesting to know the effect of caffeine on such a caffeine degrading strain as compared to other strains that are incapable of degrading caffeine. The present study aims to examine the effect of caffeine on Pseudomonas sp. isolated in our laboratory that is capable of degrading high concentrations of caffeine (Dash and Gummadi, 2006b; Gokulakrishnan et al., 2007) and on other bacterial species (both Gram negative and Gram positive bacteria). This information will be relevant to justify the negative effect that caffeine, released from byproducts of coffee processing plants, has on soil and aquatic microflora thereby disturbing ecological balance. The role of plasmid towards caffeine tolerance in this strain was also investigated.
MATERIALS AND METHODS
Microorganism and Media
This study was carried out at the Department of Biotechnology (Applied and Industrial Microbiology Laboratory), Indian Institute of Technology from November 2006 to June 2007. Caffeine degrading strain Pseudomonas sp. NCIM 5235 previously isolated in our laboratory was maintained on nutrient agar medium which had the following composition (g L-1): beef extract 1; yeast extract 2; peptone, 5; NaCl, 5 and agar, 25 and was sub cultured every two weeks. E. coli DH5α, Enterobacter aerogenes NCIM 5139, Proteus vulgaris NCIM 2813, Pseudomonas aeruginosa NCIM 5029, Staphylococcus aureus and Bacillus subtilis were also maintained on nutrient agar medium and sub cultured every two weeks.
Flask Culture Experiments
Experiments were carried out in minimal medium (MM) which had the following composition: (g L-1): Na2HPO4, 0.12; KH2PO4, 1.3; CaCl2, 0.3; MgSO4. 7 H2O, 0.3; glucose, 5 and ammonium sulphate, 2.2 at pH 7.0. Caffeine medium (CAS) had same composition of MM except in which ammonium sulfate was substituted with 1.2 g L-1 caffeine. The bacterial strains were cultured in nutrient broth for 8 h till OD600nm reached ~ 1.0. Inoculum (4% v/v) for each bacterial strain under study was then aseptically transferred to two 250 mL Erlenmeyer`s flask containing 50 mL of MM and one 250 mL Erlenmeyer`s flask containing 50 mL of caffeine medium. Samples were taken every two hours from all the flasks and OD600 nm was measured. At log phase of growth, caffeine was added from stock solution of 20 g L-1 to one of the two 250 mL Erlenmeyer`s flask containing 50 mL of MM to achieve a final caffeine concentration of 2.5 g L-1. Control experiments consisted of strains grown in MM without addition of caffeine at log phase.
Isolation of Plasmid DNA from Pseudomonas sp. and Transformation of E. coli DH5α with the Isolated Plasmid
Plasmid DNA from Pseudomonas sp. was isolated using the MiniPrep plasmid isolation kit (Qiagen). Isolated plasmid was analyzed on 0.8% agarose gel with a 1 kb DNA ladder (Biolabs Inc.) as marker. Transformation of E. coli DH5α was carried out by electroporation in a Biorad Gene Pulser apparatus according to protocol described by Sambrook et al. (1989). Transformed E. coli colonies were screened by spread plate technique on minimal medium substituted with 1.2 g L-1 caffeine and 5 g L-1 sucrose. As control, non-transformed cells were plated onto the same medium. The plates were incubated at 37°C and growth was observed after 48 h of incubation. The transformed bacterial strain was maintained on minimal medium substituted with 1.2 g L-1 caffeine and 5 g L-1 sucrose to retain plasmid and was subcultured every alternate day.
Analysis of Growth of Bacterial Strains
OD600 nm was measured for samples taken at regular intervals. Cell dry weight for the bacterial strains was calculated from previously determined values and plotted against time for getting the growth profile. Cell dry weight for the bacterial strains was calculated from the OD600nm values as per the following correlation: for E. coli, P. vulgaris, E. aerogenes and P. aeruginosa OD600nm of 1 corresponds to 0.25 g L-1 cell dry weight; for Pseudomonas sp. OD600 nm of 1 corresponds to 0.75 g L-1 cell dry weight; for Bacillus subtilis OD600 nm of 1 corresponds to 0.33 g L-1 cell dry weight and for Staphylococcus aureus OD600 nm of 1 corresponds to 0.30 g L-1 cell dry weight.
Study of Cell Morphology
Gram staining was performed on cells before and after caffeine addition at log phase and morphology of the strains was observed under microscope (Carl Zeiss) at 1000X magnification under oil immersion objective.
Cell Viability
Cell viability before and after caffeine addition during growth was estimated by determining the Colony Forming Units (CFU) of culture using standard plate count technique. Briefly, samples were serial diluted in sterile Normal Saline Solution and 100 μL of diluted sample was plated onto nutrient agar plates. The plates were incubated at 37°C for 18 h and CFU were estimated as per the following formula:
CFU mL-1 = No. of colonies x 1/ Volume of culture x dilution factor |
Statistical Analysis
All experiments were performed in quadruplicates and the values presented are the average of four experiments with ±1 to ±5% standard deviation.
RESULTS
Growth Profile of Bacterial Strains
The growth profile of different bacterial strains was analyzed in minimal medium (without caffeine), caffeine medium and minimal medium with caffeine added at the log phase of growth. E. coli DH5α strain achieved maximum growth in minimal medium with maximum cell dry weight of 0.75 g L-1 in the 10th hour of growth (Fig. 1a). No growth was noticed for this strain in caffeine medium. When caffeine was added to actively growing cultures in minimal medium, there was a subsequent decrease in OD600 nm values indicating that the cells failed to grow upon exposure to caffeine (Fig. 1a). A different scenario was observed for the caffeine degrading Pseudomonas sp. In this case, maximum cell dry weight obtained in case of minimal medium was 0.75 g L-1 in 18 h in minimal medium and 0.91 g L-1 in 12 h in caffeine medium (Fig. 1b). However, growth was enhanced and the cell dry weight increased to 1.1 g L-1 at 6 h after addition of caffeine (2.5 g L-1 final concentration). The values presented are the average of data obtained from four experiments with standard deviation of 1-4% about the mean value.
In order to test the effect of caffeine on other bacterial species, experiments were performed with Gram negative bacteria viz. Enterobacter aerogenes, Proteus vulgaris and Pseudomonas aeruginosa; and Gram positive bacteria viz. Bacillus subtilis and Staphylococcus aureus. All the above mentioned strains failed to grow in caffeine medium and achieved maximum growth in minimal medium. Upon caffeine addition at log phase, the growth of all the bacterial strains tested was inhibited as indicated by non increase of cell dry weight values, the effect more pronounced in case of Bacillus subtilis (Fig. 1c).
Morphology
Gram staining was performed to study the morphology of the bacterial strains before and after addition of caffeine to growing cultures. Interesting results were obtained upon microscopic examination at 1000X magnification. The prominent morphological change was observed for E. coli DH5α, which formed long filamentous structures upon addition of caffeine (Fig. 2b) instead of the normal short rods (Fig. 2a) and this was observed after 3 h of caffeine addition. The length of the filaments further increased with time and also a decrease in the number of cells was observed. The
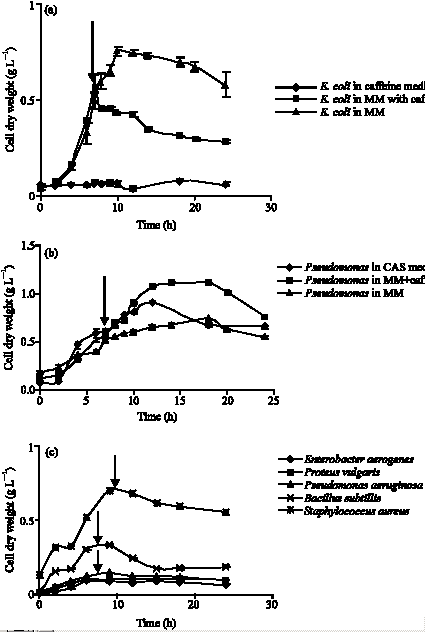 |
| Fig. 1: | Growth profile of (a) E. coli DH5α (b) Pseudomonas sp. and (c) other bacterial strains in minimal medium (MM), minimal medium with caffeine added at log phase and caffeine medium. Arrow indicates time point of caffeine addition |
caffeine degrading Pseudomonas sp. shows no such morphological change and appeared same even after addition of caffeine (Fig. 2c, d). On the other hand, Enterobacter aerogenes, Proteus vulgaris and Pseudomonas aeruginosa were seen to be intact before addition of caffeine but undergo complete lysis upon caffeine addition to growing cells at log phase (Fig. 2e-h). Similar observation was also found for Gram positive bacterium Bacillus subtilis (Fig. 2i, j). However, no distinct change was observed for Staphylococcus aureus apart from the decrease in cell growth after addition of caffeine (Fig. 2k, l).
Cell Viability
The viability of the bacterial strains after addition of caffeine at log phase was measured by standard plate count technique. The Gram negative bacterial species E. coli DH5α, Enterobacter
 |
| Fig. 2: | Morphology of bacterial strains before and 6 h after caffeine addition to actively growing cultures. (a) E. coli DH5α before addition of caffeine and (b) after 6 h of caffeine addition; (c) Pseudomonas sp., before addition of caffeine and (d) after 6 h of caffeine addition; (e) Proteus vulgaris before addition of caffeine and (f) after 6 h of caffeine addition; (g) Enterobacter aerogenes before addition of caffeine and (h) after 6 h of caffeine addition; (i) Pseudomonas aeruginosa before addition of caffeine and (j) after 6 h of caffeine addition and (k) Staphylococcus aureus before addition of caffeine and (l) after 6 h of caffeine addition |
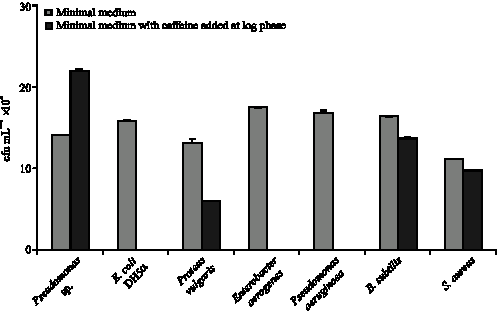 |
| Fig. 3: | Viability of bacterial strains before and 6 h after addition of caffeine at log phase of growth determined by standard plate count technique. Colony counts are expressed as cfu mL-1x108 |
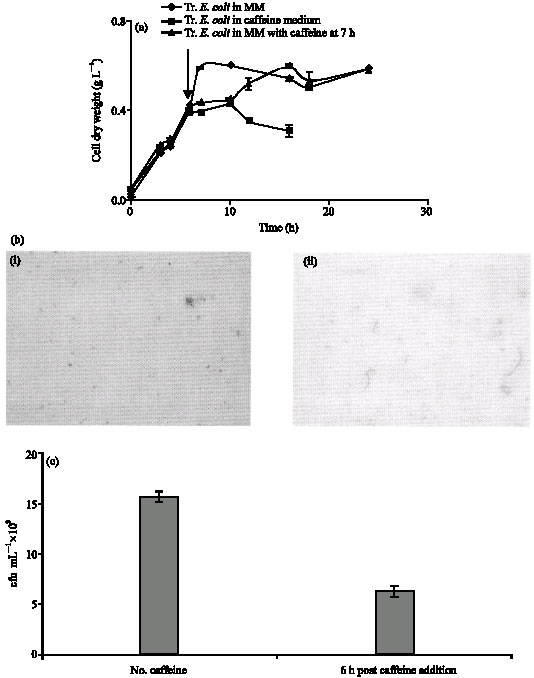 |
| Fig. 4: | Effect of caffeine on E. coli DH5α transformed with plasmid form Pseudomonas sp. (a) Growth profile of transformed E. coli DH5α in Minimal Medium, Minimal medium with caffeine added at log phase and Caffeine medium. (b) Morphology of transformed E. coli DH5α (i) before addition of caffeine and (ii) 6 h after addition of caffeine. (c) Viability of transformed E. coli DH5α |
aerogenes and Pseudomonas aeruginosa were rendered completely non viable after exposure to caffeine as shown in Fig. 3. This was also observed for almost all the dilutions of the bacterial cultures and can be and regarded as the complete loss of viability. However the count of the caffeine degrading Pseudomonas sp. was enhanced from 14±0.04x108 to 22±0.1x108 cfu mL-1 after caffeine addition. Proteus vulgaris also showed some resistance to caffeine as the cells was viable after caffeine addition although the bacterial count was decreased to 6x108 from 13x108 cfu mL-1. In case of Gram positive bacteria Bacillus subtilis and Staphylococcus aureus the viability was not much affected after caffeine addition, the bacterial count only decreased to 14±0.05x108 cfu mL-1 from 16±0.3x108 cfu mL-1 in case of Bacillus subtilis and to 9.8±0.05x108 cfu mL-1 from 11.2±0.2x108 cfu mL-1 in case of Staphylococcus aureus after caffeine exposure. For both the strains, the bacterial counts were same in minimal medium with caffeine added at log phase and in minimal medium without caffeine addition, indicating that caffeine addition does not affect cell viability in Gram positive bacteria as it does for Gram negative bacterial species. The values presented are the average of data obtained from four experiments with standard deviation of 0.3-2% about the mean value.
Effect of Caffeine on E. coli DH5α Transformed with Plasmid from Pseudomonas sp.
The effect of caffeine on E. coli DH5α transformed with plasmid (pCS1182) from caffeine Pseudomonas sp. was studied in minimal medium with and without caffeine addition at log phase and also in caffeine medium. Unlike the non-transformed E. coli DH5α, the transformed strain was found to be capable of growing in caffeine medium upto 10 h after which the growth ceases possibly due to the depletion of nutrient sources (Fig. 4a). The cell dry weight of transformed E. coli drops after 10 h in caffeine medium due to the depletion of nutrients i.e., caffeine and sucrose in the medium. In the case of MM alone, stationary phase was also attained ~ 10 h (similar to MM with caffeine) but the biomass yield was more when compared to caffeine medium. On the other hand in minimal medium substituted with caffeine at 7 h, the increase in growth can be attributed to the use of caffeine as an additional nutrient after the original nutrient in the medium has been exhausted, as seen in case of diauxic growth.
Morphological studies showed that upon caffeine addition the cells became filamentous, as seen in the case of non transformed E. coli DH5α (Fig. 4b). To some extent lysis was observed in the cell population. Viability studies showed that unlike the non transformed E. coli DH5α, the transformed bacteria retained viability. However the count dropped to 6x108 cfu mL-1 from 15x108 cfu mL-1.
DISCUSSION
The genus Pseudomonas has been long implicated in utilizing toxic matter as nutrient for its growth. Most of the time such compounds are toxic to the growth of other microbial strains and this is an important feature for the propagation of pseudomonas in the contaminated site. Pseudomonas sp. isolated in our laboratory from coffee plantation soil is capable of utilizing caffeine (1, 3, 7-trimethylxanthine) as sole carbon and nitrogen source and is capable of degrading high concentrations of caffeine (Dash and Gummadi, 2006b; Gokulakrishnan et al., 2007). It was interesting therefore to see the effect caffeine has on this strain and other microbial strains in terms of fundamental aspects such as growth, morphology and cell viability.
Caffeine when added at log phase of growth retards the growth of E. coli, Enterobacter aerogenes, Proteus vulgaris, Pseudomonas aeruginosa and Bacillus subtilis within a very short time, but the growth of Pseudomonas sp. was enhanced upon addition of caffeine. This is because of the utilization of caffeine as a source of nutrient by the caffeine degrading strain and consequent conversion of caffeine to non toxic intermediates. Previous studies on E. coli have shown that caffeine inhibits synthesis of DNA (Sandlie et al., 1980) and impairs RNA and protein synthesis. This can be the reason for non viability of E. coli cells upon caffeine exposure and lysis in case of other bacterial species.
The reduction in growth of the bacterial strains is very closely associated with changes in cell morphology in the various bacterial strains. Both the transformed and non transformed E. coli DH5α strains form long filamentous structures after addition of caffeine to growing cultures. The length of these filamentous forms was noted to increase with further incubation in the caffeine supplemented medium. Although there are reports on formation of long filamentous structure upon exposure to caffeine for various strains of Aerobacter (SundarRaj and Dhala, 1965) till to date there are no reports on the effect of caffeine on morphology of E. coli and other bacterial strains. However, previous reports show that E. coli subjected to stress factor such as nutrient depletion forms long filaments instead of separating into individual daughter cells (Koch, 2005). Filamentous form of E. coli has been also reported in case of exposure of the bacteria to chromate (Ackerley et al., 2006). Perhaps the same mechanism is adopted by E. coli to overcome the effect of caffeine on growing culture. Complete or partial lysis of the cells is noted in case of other Gram negative bacterial species under study. Similarly cell lysis was noted in the case of Gram positive bacterium, Bacillus subtilis after caffeine addition. This might be due to inhibition of important cellular functions such as protein synthesis or DNA metabolism by caffeine. In contrast, no noticeable change in morphology or lysis was observed in other Gram positive bacterium Staphylococcus aureus after addition of caffeine. These results clearly showed that coccus form is more resistant to caffeine than bacilli form.
Caffeine also affects the viability of the bacterial strains. E. coli loses viability due to the abnormal elongation of cells. The other gram negative bacterial strains studied such as Enterobacter aerogenes and Pseudomonas aeruginosa are non-viable due to complete cell lysis that occurs after addition of caffeine. On the other hand, Gram positive bacteria Bacillus subtilis and Staphylococcus aureus retain cell viability after caffeine exposure. This perhaps can be attributed to the basic differences in cellular structures of Gram positive and Gram negative bacteria.
In previous studies it has been shown that the caffeine degrading Pseudomonas sp. harbors a 12 kb plasmid that is supposed to be involved in caffeine degradation (Dash and Gummadi, 2006b). An attempt was made to evaluate the involvement of this plasmid in caffeine tolerance. Results show that the E. coli DH5α strain is capable of growing in caffeine medium and upon addition of caffeine to growing cells, the cell growth achieves constant value instead of decreasing. Also the transformed E. coli DH5α was viable after caffeine exposure although the viability is reduced to almost 50%. However, the morphological changes observed for non transformed E. coli DH5α are also observed for the transformed one i.e., same filament formation is observed also for the transformed E. coli DH5α. These findings indicate the possible role of the plasmid in conferring resistance to caffeine in E. coli DH5α. However the morphological change due to caffeine appears to be an integral aspect of E. coli cell which is not eliminated by the plasmid. Identification of genes on the plasmid can throw light the subject.
In conclusion, caffeine brings about changes in cell morphology in E. coli and causes cell lysis in certain other bacterial strains. However such an effect was not observed in the caffeine degrading Pseudomonas sp. indicating that the caffeine degrading strain has some intrinsic mechanism to counteract the effect of caffeine. Further studies on the mechanism of action of caffeine on bacterial strains will help in understanding the basics of the evolution and survival of xenobiotic degrading strains in nature.
ACKNOWLEDGMENT
This study was funded by grant from Department of Science and Technology, Government of India (Grant No. BIO/06-07/017/DSTX/GSAT).
REFERENCES
- Ackerley, D.F., Y. Barak, S.V. Lynch, J. Curtin and A. Matin, 2006. Effect of chromate stress on Escherichia coli K-12. J. Bacteriol., 188: 3371-3381.
CrossRefDirect Link - Dash, S.S. and S.N. Gummadi, 2006. Catabolic pathways and biotechnological applications of microbial caffeine degradation. Biotechnol. Lett., 28: 1993-2002.
Direct Link - Dash, S.S. and S.N. Gummadi, 2006. Biodegradation of caffeine by Pseudomonas sp. NCIM 5235. Res. J. Microbiol., 1: 115-123.
CrossRefDirect Link - Gokulakrishnan, S., K. Chandraraj and S.N. Gummadi, 2007. A preliminary study of caffeine degradation by Pseudomonas sp. GSC 1182. Int. J. Food. Microbiol., 113: 346-350.
Direct Link - Koch, A.L., 2005. Shapes that Escherichia coli cells can achieve, as a paradigm for other bacteria. Crit. Rev. Microbiol., 31: 183-190.
Direct Link - Ramanaviciene, A., V. Mostovojus, I. Bachmotova and A. Ramanavicius, 2003. Anti-bacterial effect of caffeine on Escherichia coli and Pseudomonas fluorescens. Acta Medica Lituanica, 10: 185-188.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link