
M. Rajkumar
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
R. Thavasi
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
P. Perumal
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
Jean-Paul Trilles
Equipe Adaptation Ecophysiologique et Ontogenese,
UMR 5171 (UM2-IFREMER-CNRS) Genome, Populations, Interactions, Adaptations, University Montpellier II, CP 092, Place Eugene Bataillon, 34095 Montpellier Cedex 5, France
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 12 | Page No.: 972-977







ABSTRACT
The attempt of the present study is to achieve an approach of the magnitude of parasite induced secondary infection estimation of Vibrio, Salmonella and Pseudomonas loads as well as the total heterotrophic bacteria using a battery of biochemical tests. The parasitic isopod Nerocila phaiopleura was collected from commonly available fish Stolephorus commersonii at Parangipettai, India. Parasites settled on the branchial and external body surface of the fish causes red coloured skin lesions, where secondary bacterial infections were detected. THB and Vibrio counts were higher in the infected hosts than in the healthy fish and it was found to 7.2x105 and 3.5x103 cfu g-1 in branchial regions and 5.4x105 and 2.3x103 cfu g-1 in the body surfaces, respectively. Salmonella counts were found as 6.0x103 and 3.6x101 cfu g-1 in branchial regions of the infected and uninfected fish and in body regions, 3.9x103 and 1.03x102 cfu g-1, respectively. Pseudomonas counts were 7.0x102, 4.9x101 cfu g-1 in branchial regions and 6.2x102, 5.8x101 cfu g-1 in the body surfaces of the infected and uninfected fish. Statistical analysis of the results (t-test) showed high significant value (p<0.05).
PDF Abstract XML References
How to cite this article
M. Rajkumar, R. Thavasi, P. Perumal and Jean-Paul Trilles, 2007. Parasite Induced Vibriosis in Stolephorus commersonii. Research Journal of Microbiology, 2: 972-977.
URL: https://scialert.net/abstract/?doi=jm.2007.972.977
URL: https://scialert.net/abstract/?doi=jm.2007.972.977
INTRODUCTION
Aquatic animals are continuously bathed in an aqueous suspension of microorganisms and their external surfaces are always in contact with them. Existence of pathogenic microbes in an aquatic environment is inevitable. Infections generally occur when the animal is immunologically incompetent either due to physical or biological stress such as parasitic attack. Thus, secondary infections are induced by opportunistic bacteria, which invade in to the body of parasitized organisms. Access and settlement of pathogenic microbes are facilitated through the injuries caused by parasites. In general, there are two types of secondary infections; an attack by the harmful organism is sometimes possible given the general debility due to the primary infection; the other is an active transfer of the pathogen by the parasite (Kabata, 1970).
The family Vibrionaceae is an indigenous group of bacteria in the biota of marine and estuarine environments, constituting 0.1-60% of the total heterotrophic bacteria (Simidu and Tsukamoto, 1985). Vibrio species are implicated as primary as well as secondary pathogens in fin and shellfish.
Freshly caught marine fish harbors a large number of bacteria on their skin, gill surface and intestine. Most of them are normally saprophytic but a few become pathogenic either after (parasitic injury) to the animal or when adverse environmental or physiological conditions prevail (Bisset, 1946; Schaperclaus, 1989). Fish are subjected to various diseases, some with infectious aetiology. More than 50 bacterial species have been reported to associate with diseases in freshwater and marine fishes. The greatest economic losses were induced by Aeromonas hydrophila, Renibacterium salmonarum, Streptococcus sp. (Austin and Austin, 1987; Rohovec, 1991). The most severe bacterial diseases encountered in Norwegian aquaculture were vibriosis mainly due to V. salmonicida (Holm and Jorgensen, 1987) and a furunculosis induced by A. salmonicida (Lunder and Hastein, 1990).
Incidence of infecting parasites like isopods may severe enough to produce open lesions on the fish, irritant to the epithelial layers. So, access of the smaller unicellular which can be quite destructive. Some studies suggested that fish parasites might be vectors for viral, bacterial and fungal pathogens. Nigrelli (1950) reported that the ectoparasite copepod Ergasilus sp. is the etiological agent for bacterial diseases in fish. Casack and Cone (1985) observed that the Gyrodactylus avalonia, carry pathogenic bacteria into fish and A. punctata, infect the fish secondarily at the site where the parasite Argulus sp. settle. Parasitic isopods and the copepod Lernaea sp. were reported transferring viruses causing dermal tumors in fish (Simidu and Tsukamoto, 1985). Overstreet and Howse (1977) reported that the haemorrhagic lesions produced by parasitic isopods are possible sites for secondary infections by microorganisms. Bullock (1971 and 1981) and Snieszko (1983) stated that infection might be communicable as long as infected fish are present in the environment with causative agent. A disease outbreak may occur at any time particularly when the fish is under stress.
Outbreak of bacterial diseases in fish subsequent to parasitic isopods infection is scarce with few studies carried out in India. As a result, we are interested to report the incidence of such a secondary bacterial infection in the fish Stolephorus commersonii Lacépède, 1803 (Actinopterygii; Clupeiformes; Engraulidae).
MATERIALS AND METHODS
Identification of Parasite
During the routine observation of S. commersonii in fish landings at Parangipettai (Lat. 11° 29'N; Long. 79° 46'E, India) an isopod parasite was observed on fish with an average length and weight of 11.6±0.21 cm and 9.68±0.14 g, respectively (Fig. 1). The parasite was identified as Nerocila phaiopleura Bleeker, 1857 (Crustacea; Isopoda; Cymothoidae). Incidence of isopods was maximal on the branchial and the body surface of the fish. At the site of parasitic settlement, skin lesions were observed in the infected fish.
 | |
| Fig. 1: | Nerocila phaiopleura on Stolephorus commersonii |
The damaged area was red coloured and devoid of scales. The work was carried out at CAS in marine Biology, Annamalai University, Parangipettai, Tamil Nadu, India in March, 2007.
Microbiological Analysis
Microbiological study was performed after removing 1 g of the injured skin from parasite infected and uninfected fishes. Tissues excised were separately homogenized with sterilized seawater and subjected for further microbiological investigations.
One milliliter of the serially diluted samples were used for the estimation of Total Heterotrophic Bacteria (THB) and Vibrio count (VC). Salmonella and Pseudomonas were also counted. THB was evaluated using Zobell marine agar, VC using thiosulphate citrate bile salts sucrose agar (TCBS), Salmonella in SS agar and Pseudomonas in centrimade agar. All mean counts were expressed as colony forming units g-1 (cfu g-1). Isolates were purified and stored in nutrient agar slants fewer than 4°C. Bacterial strains isolated from the aforementioned media were identified according to Bergey’s manual of determinative bacteriology (Buchanan and Gibbons, 1974) based on their biochemical and physiological characteristics.
RESULTS AND DISCUSSION
In the parasitized fishes, THB, Vibrio, Salmonella and Pseudomonas loads were higher than the uninfected fishes (Table 1).
In the branchial region of the parasitized fish, THB was found to be 7.2x105 cfu g-1 whereas in healthy fish it was 1.2x103 cfu g-1 and the Vibrio loads were 3.5x103 and 1.2x101 cfu g-1, respectively. Salmonella and Pseudomonas counts rose to 6.0x103 and 7.0x102 cfu g-1 in parasitized fish whereas they were 3.6x101 and 4.9x101 cfu g-1, respectively in uninfected fishes.
The parasitized areas of the body surface of fish showed a THB value of 5.4x105 cfu g-1 whereas in uninfected specimens it was only 1.5x103 cfu g-1. The VC from the body surface was 2.3x103 and 2.1x102 cfu g-1 in infected and uninfected fish. The Salmonella counts were of 3.9x103 and 1.03x102 cfu g-1, for Pseudomonas 6.2x102 and 5.8x101 cfu g-1, in parasite infected and uninfected fish, respectively.
The Vibrio isolates were comprised of V. angullarium, V. parahaemolyticus, V. cholerae and V. vulnificus. The two dominant species were V. angullarium and V. parahaemolyticus. Salmonella and Pseudomonas isolates showed only S. typhi and P. aeruginosa.
A number of contaminations were reported resulting from eating Vibrio contaminated raw seafoods (Sanjeev and Stephen, 1994). The major contaminants are V. parahaemolyticus, V. cholerae, V. vulnificus, V. fluvialis and V. alginolyticus. The presence of human enteric organisms on marine food products clearly indicates the contamination of terrigenous origin.
| Table 1: | Total heterotrophic bacteria and Vibrio counts on infected and uninfected Stolephorus commersonii |
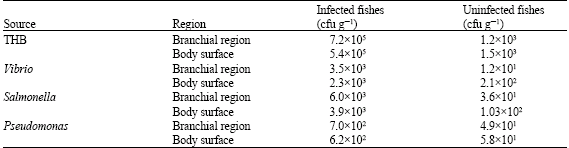 | |
| cfu g-1: Colony forming units/gram | |
Ulcerations or external lesions in fish might have been caused by a number of factors other than bacterial infection induced by parasitic attack (Sindermann, 1979), such as polluted environment and other adverse influences. However, parasites also contribute significantly to the spread of microbial diseases. Parasitic crustaceans are the largest fish parasites, which cause considerable damages to their hosts. They were involved in the spread of Lymphocystis disease (Nigrelli, 1950), Icthyophorrus contamination (Mann, 1970) and other secondary microbial infections. Overstreet and Howse (1977) proposed that haemorrhagic lesions in the spotted gore parasitized by the cymothoid Anilocra acuta, were subjected to secondary infection. In the present study, cymothoid N. phaiopleura parasitizing S. commersonii paved the way for pathogenic germs.
The bacterial load involved in the microbial infection depends on the site infected. In the present study, a regional difference for bacterial proliferation was observed: THB, VC, Salmonella and Pseudomonas counts were higher in the host’s branchial region than in the body surface in both infected and uninfected fish. This may be due to the direct exposure of gills to the outer environment. The higher THB, VC, Salmonella and Pseudomonas counts in the branchial region could be attributable to the severity of the lesions at that level, as reported in the Creole fish parasitized by N. acuminata (Rand, 1986). Also, the recorded frequent contamination of this area may be due to the respiratory water current that carries bacteria along with food materials and could facilitate bacterial invasion. Besides, the increased level of bacterial population may also lead to further infections.
Pathogenic microbes and parasites could damage the physiological and reproductive activities of the host fish (Ranjit Singh and Padmalatha, 1997; Vismanis and Kondratovics, 1997). The bacterial invasion in the branchial region reduces the respiratory area injuring the gill lamellae and affects respiration as well as excretion of nitrogenous materials and disturbs the osmotic balance (Rand, 1986; Meenashi, 1997; Ravichandran et al., 2001). Lesions induced by parasitic attack become the sites for secondary infections by opportunistic bacterial and fungal pathogens that combined with the parasitosis impacts, which cause deleterious effects on the physiology of the animal and leads to death.
The attempt of the present study is to achieve an approach of the magnitude of such a secondary infection counting the Vibrio, Salmonella and Pseudomonas loads as well as the total heterotrophic bacteria using a battery of biochemical tests. THB, VC, Salmonella and Pseudomonas counts were higher in the infected area of fish, vulnerable to attack by the microorganisms. The two dominant isolated Vibrio species are V. angullarium and V. parahaemolyticus. V. angullarium has been earlier reported as a potential fish pathogen by different authors (Kusuda, 1966; Hacking and Budd, 1971; Parker and Smith, 1984; Singh et al., 1996). Vibriosis in fish caused by V. angullarium is known to affect a wide variety of fish species from brackish water to seawater (Loganathan, 1985). Lightner (1977) reported that V. parahaemolyticus is among the causative organisms of Vibriosis in prawn and also associated with outer secondary infections. This species was also reported from cultured marine fishes (Sindermann, 1970; Raghukumar, 1971; Ravichandran et al., 2001). V. parahaemolyticus has not been specifically identified as a pathogen of marine fish although Kusuda (1966) has reported Vibrios causing ulcerous diseases in cultured marine fish with close similarity to intestinal inflammation in man caused by V. parahaemolyticus. According to the above results, V. parahaemolyticus may be considered as a significant pathogen involved in secondary infection. Harrell et al. (1976) demonstrated that isolates of V. anguillarium act as the most important aetiological agent of vibriosis. In the case of uninfected fish, THB and Vibrio counts were very low. This may be due to the healthiness of the fish, which will interfere with the contact between the microbe and fish, as a result of continuous secretion of mucus from the skin of the fish.
Salmonella has been reported in the gut of Tilapia and carps (Iyer and Shrivastava, 1989; Ogbondeminu, 1993) and as a fish pathogen in salmon (Austin et al., 1982) as reposted in the present study. Occurrence of Pseudomonas in the infected fish was witnessed by Eddy and Jones (2002), Simidu and Tsukamoto (1985) and Salmonella by Karunasagar et al. (2004). Pseudomonas exists throughout the aquatic environment associated with both healthy (Evelyn and McDermott, 1961; Bullock and Sniesko, 1969) and unhealthy fish. Presence of Klebsiella pneumoniae in a badly damaged fin and tail has been recorded by Austin and Austin (1999). P. anguilliseptica and P. fluorescens were reported as a causative agent of red spot and generalized septicaemia in rainbow trout and most of the marine species (Austin and Austin, 1999). Statistical analysis of the results obtained from the present study showed a high significance of p<0.05.
ACKNOWLEDGMENTS
The authors are thankful to the authorities of Annamalai University for the facilities provided and to the Indian Council of Agricultural Research for the financial support.
REFERENCES
- Eddy, S.D. and S.H. Jones, 2002. Microbiology of summer flounder Paralichthys dentatus fingerling production at a marine fish hatchery. Aquaculture, 211: 9-28.
Direct Link - Hacking, M.A. and J. Budd, 1971. Vibrio infection in tropical fish in a freshwater aquarium. J. Wildlife Dis., 7: 273-280.
PubMed - Ranjit Singh, A.J.A. and C. Padmaltha, 1997. Parasitic infestation on commercially important sardines. Curr. Sci., 72: 171-171.
Direct Link - Ravichandran, S., A.J.A. Ranjit Singh and N. Veerappan, 2001. Parasite-induced Vibriosis in Chirocentrus dorab off Parangipettai coastal waters. Curr. Sci., 80: 622-623.
Direct Link - Simidu, U. and K. Tsukamoto, 1985. Habitat segregation and biochemical activities of marine members of the family Vibrionaceae. Applied Environ. Microbiol., 50: 781-790.
PubMedDirect Link