
T.O. Femi-Ola
Not Available
E.Y. Aderibigbe
Not Available
L. Awoyemi
Not Available
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 12 | Page No.: 910-917







ABSTRACT
The survival of Amitermes evuncifer (Silvestri) on five different Nigerian wood species: Terminalia superba Engl and Diels, Khaya grandifoliola C.D.C., Milicia excelsa Welw C.C. Berg, Mansonia altissima A. Chev and Brachystegia eurycoma Harms for seven days was investigated. Some physicochemical properties of the wood samples were also determined. The survival rate on Terminalia superba was highest (22.5%) and least on Mansonia altissima and Brachystegia eurycoma (0%), respectively. The other wood types had less than 15% live termites. Micrococcus luteus and Bacillus species were persistently isolated from the hindgut of the termites throughout the period of the no-choice food experiment. The total bacterial population in the hindgut of the termites declined from 104 to 102 cfu mL-1. The moisture content, density and pH of the wood samples ranged from 10 to 18%, 409 to 758 kg m-3 and 5.52 to 5.88, respectively. There was a high degree of correlation (R = 0.96; R2 = 0.922) between the amount of wood consumed and survival of A. evuncifer. However, the correlation coefficient between the amount of wood consumed and density of wood was negative (R = -0.675; R2 = 0.455).
PDF Abstract XML References
How to cite this article
T.O. Femi-Ola, E.Y. Aderibigbe and L. Awoyemi, 2007. Microbiology of the Hindgut and Survival of Amitermes evuncifer (Silvestri) on Some Nigerian Woods. Research Journal of Microbiology, 2: 910-917.
URL: https://scialert.net/abstract/?doi=jm.2007.910.917
URL: https://scialert.net/abstract/?doi=jm.2007.910.917
INTRODUCTION
Termites occur throughout the temperate and tropical regions of the world and they cause majority of damage to wood in use (Duryea et al., 1999; Peralta et al., 2004). Cellulose, the most abundant component of plant biomass and wood is the major constituent of the diet of termites. Termites are destructive when they feed upon wooden structures valuable to man. They also cause damage to living plants and dry construction timbers because of their search for cellulose as food (Myles, 1999). Wood is composed of cellulose, hemicellulose, lignin and extractives (Illston et al., 1981). The extractives do not only impart colouration to the heartwood, but also confer resistance to both fungal and insect attacks (Illston et al., 1981). Extractives that have been reported in woods include terpenoids, flavonoids, quinines, tannins, authocyanins, tropolones, resins, glycosides, sugars and fatty acids (Grace, 1997; Nakayama et al., 2000; Neya et al., 2004). The resistance of certain woods to termite infestation has been attributed to the chemicals make up of the wood, which could be distasteful, repellent or toxic (Carter and Dell, 1981; Duryea et al., 1999). Scheffrahn (1991) reported that extracted allelochemicals from cypress heartwood operated as feeding deterrents while accompanying mortality from cypress heartwood was due to starvation.
Susceptible woods are being protected from termites by the use of toxic or repellent chemical preservatives. Some chemicals are now unacceptable because of their potentially adverse environmental effects (Peralta et al., 2004). The application of extractives of wood with natural resistance to termite attack on susceptible wood could be a better alternative control measure. The present study investigated the microbiology of the hindgut of a wood-eating termite Amitermes evuncifer (Silvestri) and the survival of the termites on some woods commonly used in building construction and furniture work in Nigeria. The woods studied are widely distributed in Africa and some other parts of the world. They include Terminalia superba Engl and Diels (family Combretaceae), Khaya grandifoliola C.D.C. (family Meliaceae), Milicia excelsa (Welw) C.C. Berg (Family Moraceae), Mansonia altissima A. Chev (family Sterculiaceae) and Brachystegia eurycoma Harms (family Fabaceae) (Keay, 1989).
The economic loss imposed by termite attack on valuable woods and wooden structures is enormous. This work is aimed at screening for woods that have natural resistance to termite infestation.
MATERIALS AND METHODS
Collection of Materials
Wood-eating termites were collected from infested planks and bamboo in the University of Ado-Ekiti campus, Ado-Ekiti. Termite infested wood containing both the termite workers and soldiers was moistened with distilled water and kept in plastic boxes prior to processing. Termite samples collected were identified as Amitermes evuncifer of the order Isoptera, family Termitidae at the Department of Zoology, University of Ado-Ekiti, Nigeria.
Wood shavings of five tree species were collected from Bashiri Sawmill in Ado-Ekiti. These species are; Milicia excelsa (Welw) C.C. Berg, Mansonia altissima A. Chev, Khaya grandifolila C.D.C., Brachystegia eurycoma Harms and Terminalia superba Engl and Diels.
Termite Survival Tests
Survival of Amitermes evuncifer on five separate shavings of heartwood was measured. Ten grams of wood shavings were placed on sand in a plastic container (12x11 cm). Each container was filled with 500 g of sand and moistened with 50 mL of water to maintain a relative humidity near saturation (Carter et al., 1978; Duryea et al., 1999). The sand (plus substrate) was placed in a chamber while forty termites were added to each container and their conditions were checked daily for 1 week. The control experiment consists of a dish with no substrate (wood shaving), having forty termites, which was set up to determine if termites needed to eat during the experimental period. The survival rate was determined by the number of live termites found in each treatment after the 7 day period.
Assessment of Microflora in the Hindgut of A. evuncifer
Termite samples were taken on daily basis from each of the separate wood shavings and analyzed to determine the bacterial load in the gut samples. Each termite was surface sterilized by rinsing in 10% (v/v) sodium hypochlorite solution and then sterile distilled water. Each of the termites was dissected aseptically to remove the hindgut, which was then placed in 10 mL sterile Ringer’s solution containing the following chemicals (g L-1): NaCl, 10.0; KCl, 0.42; CaCl2.2H2O, 0.48 and NaHCO3, 0.20 (Amund et al., 1986). The gut sample was then crushed using a sterile glass rod to release the intestinal contents.
Serially diluted gut suspension was plated on Nutrient agar plates in replicates and incubated at 37°C for 48 h. Colonies on plates were counted to determine the total colony forming units of bacteria (cfu) count. Pure cultures of the isolates were obtained by sub culturing serially onto Nutrient agar plates.
Characterization and Identification of Bacterial Isolates
Pure cultures of the isolates were identified based on their cultural, morphological and biochemical characteristics in accordance with the taxonomic scheme of Barrow and Feltham (1999).
Determination of Moisture Content
Moisture content was determined by the AOAC (1990) method. Ten grams of wood shaving of the samples were dried at 50°C to constant weight in a hot air oven (Gallenkamp). The moisture content (%) was calculated using the formula below;
MC = 100(W1 - W2)/W2 |
Where,
| MC | = | Moisture content (%). |
| W1 and W2 | = | Initial and final weight, respectively (g). |
Determination of Wood Density
Wood disk samples were cut from each wood type. The samples were weighed with a weighing balance and the dimensions were also measured using a vernier caliper. The density was calculated using the formula below;
D = M/V |
Where,
| D | = | Density (kg m-3), |
| M | = | Mass (kg) |
| V | = | Volume (m3) |
Determination of pH
Ten grams each of the wood shaving samples were mixed with 100 mL of sterile distilled water in 250 mL conical flask, shaken and allowed to stand for 3 h. The pH of the mixture was read with a pH meter. The mean of triplicate determinations were calculated.
Statistical Analysis
Analysis of Variance (ANOVA) and regression analysis of data obtained were carried out using SPSS (Statistical Package for Social Sciences) version 11.0 software.
RESULTS AND DISCUSSION
The moisture content, density and the pH of wood samples investigated are shown in Table 1. The moisture content ranged from 10 to 18%, while the pH value ranged from 5.52 to 5.88. The density ranged from 409 to 758 kg m-3. At the end of the seven days no-choice food experiment, Khaya grandifoliola and Milicia excelsa were left with 5 and 7.5% live termites, while Terminalia superba had 22.5% surviving termites. Survival on Brachystegia enrycoma and Mansonia altissima were 0%, respectively (Fig. 1). Termites consumed 3.5% of the Terminalia superba, 1.5% of Khaya grandifoliola and 1.2% of Milicia excelsa.
| Table 1: | Physicochemical properties of wood samples |
 | |
| Averages within a column followed by the same letter(s) are not significantly different, as gauged by Duncan multiple range test | |
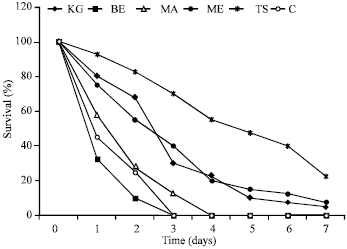 | |
| Fig. 1: | Survival of A. evuncifer on wood shaving during a seven-day no-choice food experiment |
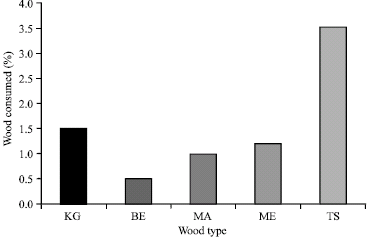 | |
| Fig. 2: | Wood consumption rate (%) by termites during a seven day no-choice food experiment |
However only about 0.5% of Brachystegia eurycoma and 1% of Mansonia altissima was consumed by A. evuncifer (Fig. 2). There was a high degree of correlation (R = 0.96; R2 = 0.922) between the amount of wood consumed and survival of A. evuncifer on the different wood samples (Fig. 3). There was a low level of inverse correlation between the wood density and the wood consumption rate (Fig. 4). The regression analysis of the physicochemical properties and wood consumption rate is shown in Table 2. There was a gradual decrease in the microbial load from the onset of the experiment to the end. There was a decrease from 104 to 102 cfu mL-1 in the bacterial population of the hindgut of the termites (Table 3). The bacterial isolates recovered from the gut include Micrococcus luteus, Serratia marcescens, Bacillus sp. and Streptococcus sp. However only Bacillus sp. and Micrococcus luteus persisted till the end of the experiment (Table 4).
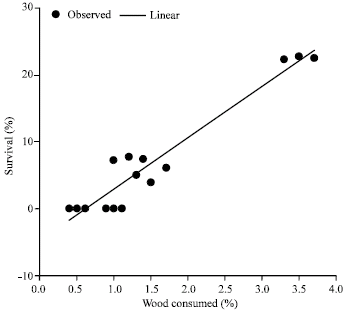 | |
| Fig. 3: | Correlation curve of wood consumed and termite survival |
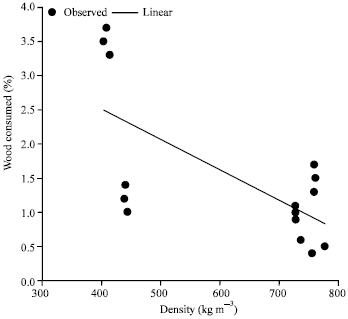 | |
| Fig. 4: | Correlation curve of wood consumed and wood density |
| Table 2: | Regression analysis of physicochemical properties, survival of termites and wood consumed as dependent variable |
 | |
| Table 3: | Microbial load (cfu mL-1) in the hindgut of A. evuncifer during a seven-day no-choice food experiment |
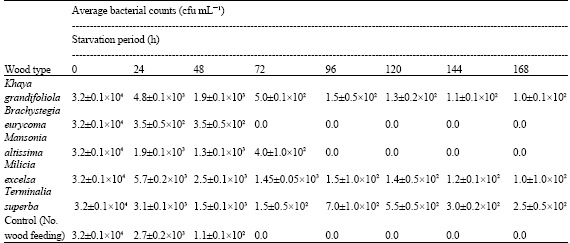 | |
| cfu = Colony forming units | |
| Table 4: | Bacterial isolates recovered from hindgut of A. evuncifer during a 7-day no-choice food experiment |
 | |
| + = Present; - = Absent | |
The study showed that Brachystegia eurycoma and Mansonia altissima were more resistant to termite infestation than the other three species used in this study. The survival rate of termites on these two woods was 0% for Brachystegia eurycoma (within 48 h) and Mansonia altissima (within 72 h). The termites’ samples died within 48 h of exposure to Brachystegia eurycoma. Termites without food starved and also died within 48 h. The high mortality rate observed in termites fed with B. eurycoma and M. altissima may be due to starvation. Among the wood species T. superba was more consumed by A. evuncifer than other wood types, indicating that the former is more susceptible to termite attack. The wood density was inversely correlated with the rate of consumption of the different wood species. This may explain why T. superba with a density of 409 kg m-3 was the highest consumed wood. Bultman et al. (1979) reported a general inverse relationship between the hardness of wood and the amount of damage it received from Coptotermes formosanus with the lighter’ softer woods being more heavily damaged than heavier harder woods. Waller et al. (1990), however reported that the wood densities of pine and mahogany (0.52 and 0.73 g cm-3), respectively did not affect the amount of wood consumed by C. formosanus. Peralta et al. (2004) also reported that wood consumption rates were not correlated significantly with their wood densities; however there was a tendency for softwoods to be more consumed by subterranean termites than the woods of intermediate hardness. The factors affecting wood consumption by termites are multifaceted and complexly linked together (Peralta et al., 2004). Factors affecting wood consumption by termites include wood species and hardness, presence of toxic substances, feeding inhibitors or deterrents, presence or absence of fungi and degree of fungal decay, moisture content of wood and soil (Nagnan and Clement, 1990; Scheffrahn, 1991; Grace, 1997). It has being well documented that in both laboratory and field tests that some antitermic chemical compounds found in woods like terpenoids and quinines acted as a natural repellent for termites (Carter et al., 1978; Scheffrahn, 1991; Grace, 1997).
There was a gradual decline in the microbial load in the hindgut of the termite A. evuncifer maintained on different wood shavings during the seven-day old no-choice food experiment (Table 2). This may be due to the change in the environmental conditions from the natural habitat to the laboratory environment resulting into reduction in the activity of the termites. The amount of wood consumed by the termites correlated with survival rate. However there were no correlations with pH and moisture content (R = 0.5), while the correlation with density of wood was negative.
CONCLUSIONS
Amitermes evuncifer Silvestri consumed more of Terminalia superba than Brachystegia eurycoma and the other wood species tested in this study, indicating that the former is more susceptible to termite attack. Wood consumption correlated with the survival of A. evuncifer, while the wood densities also appeared to influence to some extent the rate of wood consumption. Some other factors such as the chemical components and extractives in the wood may be responsible for the differential rate of wood consumption and termite survival. The present investigation has revealed that B. eurycoma and M. altissima are more resistant to termite attack. Further studies on the effects of the application of the purified extracts of these woods on susceptible woods, will provide additional information on the possibility of using wood extracts as alternative control measure to termite infestation.
REFERENCES
- Amund, O.O., O.S. Yakubu and S.L.O. Malaka, 1986. A study of bacteria from the digestive system of two advanced termites (Isoptera, Termitidae) in Nigeria. Nig. J. Biol. Sci., 1: 19-24.
Direct Link - AOAC, 1990. Official Methods of Analysis. 15th Edn., Association of Official Analytical Chemists, Washington, DC., USA., pp: 200-210.
Direct Link - Carter, F.L., A.M. Garlo and J.B. Stanley, 1978. Termiticidal components of wood extracts: 7-methyljuglone from Diospyros virginiana. J. Agric. Food Chem., 26: 869-873.
CrossRefDirect Link - Duryea, M.L., J.B. Huffman, R.J. English and W. Osbrink, 1999. Will subterranean termites consume landscape mulches? J. Arbor., 25: 143-149.
Direct Link - Grace, J.K., 1997. Influence of tree extractives on foraging preferences of Reticulitermes flaviceps (Isoptera: Rhinotermitidae). Sociobiology, 20: 35-42.
Direct Link - Myles, T.G., 1999. Review of secondary reproduction in termites (Insecta: Isoptera) with comments on role in termite ecology and social evolution. Sociobiology, 33: 1-91.
Direct Link - Nagnan, P. and J.I. Clement, 1990. Terpenes from the maritime pine Pinus pinastar: Toxin from subterranean termites of the genus Reticulitermes (Isoptera: Rhinotermitidae). Biochem. Syst. Ecol., 18: 13-16.
Direct Link - Nakayama, F.S., P. Chow, D.S. Bajiwa, J.A. Youngquist, J.H. Muechl and A.M. Krzysik, 2000. Preliminary investigation on the natural durability of Guayule (Parthenium argentatum) based wood products. Proceedings of the International Research Group on Wood Preservation 31st Annual Meeting Kona, May 14-19, 2000, Hawaii, USA., pp: 9-9.
Direct Link - Neya, B., M. Hakkou, M. Petrissans and P. Gerardin, 2004. On the durability of Burkea africana heartwood, evidence of biocidal and hydrophobic properties responsible for durability. Annu. For. Sci., 61: 277-282.
CrossRefDirect Link - Peralta, R.C.G., E.B. Menezes, E.G. Carvalho and E.D.L. Auilar-Menezes, 2004. Wood consumption rates of forest species by subterranean termites (Isoptera) under field conditions. Revista Arvore, 28: 283-289.
Direct Link - Scheffrahn, R.H., 1991. Allelochemical resistance of wood to termites. Sociobiology, 1: 257-281.
Direct Link - Waller, D.A., C.G. Jones and J.P. Lafage, 1990. Measuring wood preference in termites. Entomol. Exp. Applic., 56: 117-123.
Direct Link
malik zulfiqar Reply
1.WATER ABSORPTION
2. TENSILE
3. NAIL RETENTION
4. FLAMMABILITY
5. UV EXPOSURE
i need these tests for (plastic wood) Tiles
that,sample i will sent you by DHL, i need first your conformation and also price.
malik zulfiqar
jeddah (SAUDI ARABIA)