
M.M. Begum
Department of Plant Protection, Universiti Putra Malaysia,
43400 UPM, Serdang, Selangor D.E., Malaysia
M. Sariah
Department of Plant Protection, Universiti Putra Malaysia,
43400 UPM, Serdang, Selangor D.E., Malaysia
M.A. Zainal Abidin
Department of Plant Protection, Universiti Putra Malaysia,
43400 UPM, Serdang, Selangor D.E., Malaysia
A.B. Puteh
Department of Plant Science, Faculty of Agriculture, Universiti Putra Malaysia,
43400 UPM, Serdang, Selangor D.E., Malaysia
M.A. Rahman
Department of Plant Science, Faculty of Agriculture, Universiti Putra Malaysia,
43400 UPM, Serdang, Selangor D.E., Malaysia
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 12 | Page No.: 900-909







ABSTRACT
Histopathological studies on natural infections by Fusarium oxysporum f. sp. glycines in soybean seeds were conducted using Light Microscopy (LM) and Scanning Electron Microscopy (SEM). Asymptomatic (healthy) seeds were found free from pathogens. Infected seeds were slightly irregular in shape and appeared whitish moldy. The fungus mycelia colonized the external surface and inner tissues of the seed coat, but not in cotyledon or in embryo. Upper surfaces of seeds showed profuse colonization by mycelia and seed coat tissues became ruptured and distorted in the severely infected seeds. Vigorous mycelial growth was found in the hourglass layer of the seed coat. Effect of artificial inoculation on soybean seed germination and seedling survivability under glasshouse conditions was also studies. F. oxysporum f. sp. glycines reduced seed germination and seedling survivability by 40% and caused pre-emergence damping off of seedlings. Trichoderma harzianum isolate UPM40 and Pseudomonas aeruginosa isolate UPM13B8 were most effective candidates in inhibiting the mycelial growth of F. oxysporum f. sp. glycines in vitro.
PDF Abstract XML References
How to cite this article
M.M. Begum, M. Sariah, M.A. Zainal Abidin, A.B. Puteh and M.A. Rahman, 2007. Histopathological Studies on Soybean Seeds Infected by Fusarium oxysporum f. sp. glycines and Screening of Potential Biocontrol Agents. Research Journal of Microbiology, 2: 900-909.
URL: https://scialert.net/abstract/?doi=jm.2007.900.909
URL: https://scialert.net/abstract/?doi=jm.2007.900.909
INTRODUCTION
Fusarium oxysporum species are widespread throughout the world in a variety of climates and on many plant species as epiphytes, parasites, or pathogens (Alabouvette et al., 1998). Fusarium-induced diseases of soybeans have been attributed to different species of fungi identified from soybean pods and seeds (Sinclair, 1995; Meriles et al., 2002). F. oxysporum f. sp. glycines [(Schlect.) emend. Snyd and Hans] is a seed-borne pathogen found frequently in soybean seed lots (Nasir, 2003; Reyes-Ramirez et al., 2004; Agarwal et al., 2006). Infected soybean seeds externally appeared as shrunken, slightly irregular in shape, often with cracks in the seed coat with light pink discolored areas over seed surfaces. F. oxysporum f. sp. glycines reduced germination by rotting in severely infected seeds of soybeans (Velicheti and Sinclair, 1989). Other related diseases on soybeans are Fusarium blight or wilt and root rot (Akinsanmil and Adekunle, 2003). It was reported that F. oxysporum has ability to colonize seed surfaces and seed coats in soybean seeds (Velicheti and Sinclair, 1989). Since little information about site of infection in soybean seeds infected by Fusarium sp. is available, the present research was planned to determine the location of infection by F. oxysporum f. sp. glycines in soybean seeds.
Seed treatment by protectant fungicides provides protection of seed and seedlings against seed and soil-borne fungi. However, fungicidal treatment is not a widely accepted practice in the tropics for soybeans, primarily because of cost and lack of recommendations in some developing countries (Backman and Jacobsen, 1992). Regular use of fungicides has often drastically reduced the viability of Bradyrhizobium cells, decreasing nodulation and nitrogen fixation rates of soybeans (Hungria et al., 2005). However, there are no known resistant varieties of soybean against Fusarium sp. (Reynolds, 2001).
There has been an increase in the use of biological control of pathogens using different antagonists. The most widely used antagonists in the world include species of bacteria such as Pseudomonas, Burkholderia and Serratia. Among fungi different species of Trichoderma are popularly used in biological control programs against different fungi (Kloepper et al., 1992; Gardener and Fravel, 2002). Therefore, the aim of this research was to study the site of infections of soybean seeds by F. oxysporum f. sp. glycines and effect on seed germination and seedling establishment. Attempt was also made to screen potential antagonists against F. oxysporum f. sp. glycines in vitro.
MATERIALS AND METHODS
The experiments were conducted in the Plant Pathology Laboratory, Faculty of Agriculture, Universiti Putra Malaysia (UPM) in Serdang, Selangor in 2006.
Microscopic Observations
Histopathological studies were conducted to determine the sites of infection in naturally infected soybean seeds using Light Microscopy (LM) and Scanning Electron Microscopy (SEM). Seed samples var. Palmetto were collected from stored seeds at the Department of Plant Science, Faculty of Agriculture, Universiti Putra Malaysia. Seeds were produced in 2004 and had been stored for 1 year at 10°C with 9.5% seed moisture content. Seed samples were randomly selected and plated on moist blotter paper and incubated at alternating cycles of 12 h Near-Ultra Violet (NUV) light and darkness. After four days of incubation, seeds showing typical symptoms of disease causing F. oxysporum f. sp. glycines as well as asymptomatic (healthy) seeds were selected. Different seed components viz. seed coat, cotyledon and embryo were separated carefully and cut transversely into 2-3 mm2 pieces.
Light Microscopy (LM)
Individual seeds and their components from both symptomatic and asymptomatic (healthy) seeds were fixed in Bouin’s solution (picric acid, formaldehyde 40%, glacial acetic acid 15:5:1 v/v/v) and vacuum extracted overnight. Samples were then dehydrated in eight a series of concentrations of ethanol (30, 40, 50, 60, 70, 80, 90 and 100%) and washed twice in 98% methyl benzoate plus 2% celloidine for 96 h. Samples were then infiltrated in oven at 60°C overnight in each of the mixture of xylene and paraplast in proportions of 75:25; 50:50; 25:75 and 100%, respectively. Finally, the samples were embedded in paraffin wax and mounted in small paper box. Serial paraffin sections of 10-12 μm thick were cut using an ordinary rotary microtome. Fine sections were placed on glass slides using albumin-glycerin solution. Sections were deparafinized with xylene and stained with 0.05% toluidine blue. All stained sections were mounted in DPX mounting media and viewed under light microscope (Model Nikon FX-35DX) (Johansen, 1940).
Scanning Electron Microscopy (SEM)
Seed samples from symptomatic and asymptomatic (healthy) seeds were fixed separately in 2.5% buffered glutaraldehyde for 24 h at 4°C. Samples were washed in 0.1 M sodium cacodylate buffer (pH 7.7) and post-fixed in 1% osmium tetraoxide for 2 h at 4°C and then washed again in 0.1 M sodium cacodylate buffer three times for 10 min each. A series of dehydration was performed in seven different concentrations of ethanol (30, 40, 50, 60, 70, 80 and 90%) three times for 10 min each and finally 15 min in 100% ethanol. Samples were dried in Baltec 030 Critical Point Drying (CPD) apparatus for 30 min. Dried samples were stuck on stubs and coated with gold in a Polaron Sputter Coater and viewed under SEM (JOEL JSM 6400) (Benhamou and Chet, 1996).
Fungal Isolation, Seed Inoculation and Disease Assessment
F. oxysporum f. sp. glycines was isolated from naturally infected soybean seeds by agar plate method. Fungus was cultured on PDA for 7 days at room temperature to obtain a large amount of spores. The spores were washed off with sterile distilled water containing 1.5% sodium alginate solution. The spore suspension obtained was adjusted to a concentration of 1x107 spores mL-1 by a Neubeaur’s haemocytometer. Soybean seeds were surface sterilized in 10% Clorox® for 3 min and rinsed thrice with sterilized distilled water and dried for 1 h in a laminar flow chamber. Seeds were soaked in spore suspension (1: 2 w/v) of F. oxysporum f. sp. glycines for 1 h and surface dried over night. The number of conidia for each seed was counted by a haemocytometer and approximated at 5x105 conidia seed-1. Uninoculated seeds were soaked in 1.5% sodium alginate solution for 1 h and served as control. Seeds were subsequently sowed at the depth of 2 cm in plastic tray (39x28 x11 cm) containing sterilized soil mixture (top soil: peat soil: sand = 3: 2: 1). The experiment was done in four replications with 25 seeds per replication. Trays were arranged in a completely randomized design in the glasshouse with each tray considered as a replication. The glasshouse temperatures were 31°C (day) and 25°C (night) and RH of 85%. After 14 days, data were recorded for seed germination, pre- and post-emergence damping off and survival of seedlings.
Screening of Antagonists
Six isolates of Trichoderma sp. and three isolates of bacteria obtained from the Plant Pathology laboratory, Faculty of Agriculture, UPM were used in this study. The test was conducted in a completely randomized design with five replications. These isolates were screened for their antagonistic activity against F. oxysporum f. sp. glycines in vitro using dual culture tests based on the Percentage Inhibition of Radial Growth (PIRG).
A 5 mm diameter mycelial agar disc was cut from the margin of 7 day-old cultures of F. oxysporum f. sp. glycines and placed on one side of a 9 cm Petri dish containing PDA medium. Another 5 mm mycelial agar disc from 7 day-old culture of each of Trichoderma isolates tested was placed 3 cm away from the former disc on the same plate. The plates were incubated at ambient temperature (25±1°C) for 15 days. Antagonistic activity of Trichoderma isolates were assessed at seven days after incubation by measuring the radius of the F. oxysporum f. sp. glycines colony using the following Eq:
Where,
R1 = Radial growth of fugal pathogen colony in control plate.
R2 = Radial growth of fungal pathogen colony in dual culture plate.
During the incubation period, data regarding the time needed to grow fully over the fugal colony by Trichoderma sp. were recorded. Attempts were also made to determine the re-growth of the fungus from the inhibition zone. For testing antagonistic bacteria, a 5 mm diameter of fungal agar disc from 7-day-old cultures was placed in the middle area of a 9 cm Petri dish containing Nutrient Agar (NA) medium. The plates were incubated at ambient temperature (25±1°C) for 24 h. A loopful of bacteria from 48 h NA culture was taken and streaked in a circle around 3 cm away from the fungal agar disc. The plates were then incubated for 7 days. Data of PIRG and inhibition were recorded after 7 days of incubation. Attempts were also made to determine the growth of F. oxysporum f. sp. glycines from the zone of contact with bacteria.
Statistical Analysis
Data were analyzed statistically by ANOVA test using SAS software (SAS, 1999). Mean separation was carried out using Tukey’s Studentized Range (HSD) at p≥0.05.
RESULTS
Histopathological examinations of transverse sections of asymptomatic seeds under LM and SEM were observed. The mature soybean seeds consist of a seed coat, cotyledon and embryo. The seed coat contains a cuticle and three distinct layers namely palisade cell layer, hourglass cell layer with prominent air-spaces and parenchyma cell (Carlson and Lersten, 1987). Asymptomatic seeds (healthy) were free from fungal infections and tissues of seed components were well developed and compacted under SEM (Fig. 1A and 2). Similar trends were also observed under LM. Infected seeds by F. oxysporum f. sp. glycines appeared as shrunken externally, slightly irregular in shape and whitish moldy over the seed surface (Fig. 1B).
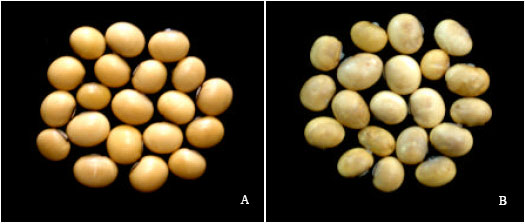 | |
| Fig. 1: | Photomicrographs showing (A) asymptomatic (healthy) soybean seeds and (B) F. oxysporum f. sp. glycines infected shrunken soybean seeds with whitish moldy over seed surfaces |
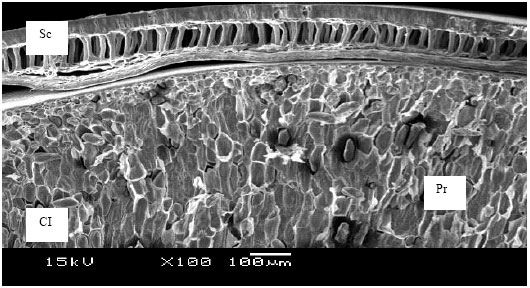 | |
| Fig. 2: | SEM photomicrographs showing transverse sections of seed coat with cotyledon in asymptomatic (healthy) soybean seeds (Abbreviations: Sc, Seed coat; Cl, Cotyledon; Cut, Cuticle; Pl, Palisade layer; Hg, Hourglass cell; Pr, Parenchyma cell) |
 | |
| Fig. 3: | Photomicrographs showing transverse sections of seed coat with cotyledon in soybean seeds naturally infected by F. oxysporum f. sp. glycines (A) Light microscopy of seed coat with cotyledon showing mycelial mat over the seed surface and inside the seed coat layer (B) Magnified SEM view of seed coat from part A showing mycelia in the hourglass and parenchyma tissues of the seed coat (C) SEM view of seed coat showing hyphae in the hourglass cell (D) Magnified SEM view from part C showing mycelial mat beneath the seed coat over cotyledon (Abbreviations: Mym, Mycelial mat; My, Mycelia; Hy, Hyphae; Pl, Palisade layer; Hg, Hourglass cell; Pr, Parenchyma cell) |
| Table 1: | Effect of Fusarium oxysporum f. sp. glycines on seed germination, pre-emergence damping-off and seedling survivability of soybeans after 14 days of seed sowing |
| Means in the same column with different letter (s) are significantly different at p≤0.05 according to HSD of arcsine transformed values | |
Transverse sections of infected seeds using LM and SEM showed the presence of fungal mycelia on the seed surface and internal tissues of the seed coat. Mycelial growth was more abundant in the hourglass layer of the seed coat where large intercellular spaces were present (Fig. 3B and C). Fungal hyphae could be distinguished in the seed tissue based on hyphal morphology. Hyphae of this fungus were hyaline, branched and stained light green with toluidine blue (0.1%). The hyphal breadth ranged from 2.5-3.8 μm (Fig. 3B) (Sharma, 1992). Mycelial mat were formed on the seed surface and beneath the seed coats (Fig. 3A and D). Severely infected seed coat appeared distorted and empty (Fig. 3B). Hyphae were not detected in any tissues of the cotyledons or in embryo in the infected seed.
Effect of infection was assessed on inoculated seeds from observations of seed germination, pre and post-emergence damping off and seedling survivability under glass house conditions (Table 1). F. oxysporum f. sp. glycines significantly reduced seed germination and seedling survivability by 40% when compared with uninoculated seeds (control).
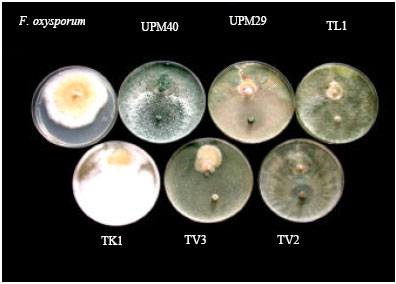 | |
| Fig. 4: | Effect of Trichoderma isolates on the radial growth of F. oxysporum f. sp. glycines in the dual culture test at 7 days of incubation on PDA |
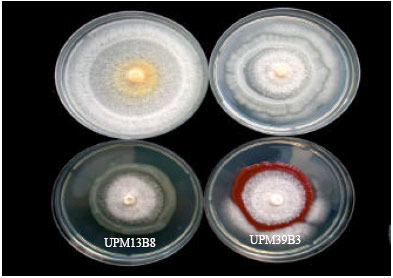 | |
| Fig. 5: | Effect of bacterial isolates on the radial growth of F. oxysporum f. sp. glycines in the dual culture test at 7 days of incubation on NA |
The germination and survivability of seedlings were recorded at 51% in inoculated seeds and of 85% in control seeds. The pathogen also caused pre-emergence damping off. Significantly higher percentage (49.0%) of pre-emergence damping off was recorded in inoculated seeds. Post-emergence damping-off of seedlings was not observed either in inoculated or uninoculated seeds.
All Trichoderma isolates tested inhibited the mycelial growth of F. oxysporum f. sp. glycines at different degrees of inhibition. PIRG by Trichoderma spp. ranged from 47.2 to 75.4%, with the highest PIRG values being recorded for isolate were UPM40 (75.4%) followed by TV2 (56.1%), TL1 (55.4%), UPM29 (50.9%), TV3 (50.3%) and Tk1 (47.2%). UPM40 grew over the fungal pathogen colony fully at 7 days after incubation, whereas TL1 and TV2 grew over the fungal pathogen colony at 9 days and 10 days after incubation, respectively (Table 2 and Fig. 4). The other test isolates did not grow over the fungal pathogen colony within 14 days after incubation.
| Table 2: | Antagonistic effect of Trichoderma isolates against F. oxysporum f. sp. glycines in the dual culture test |
 | |
| -: Indicates no overgrowth up to 14 days; *: Indicates percent inhibition of radial growth (PIRG) at 7 days of incubation Means in the same column with different letter(s) are significantly different at p≤0.05 according to HSD of arcsine transformed values | |
| Table 3: | Antagonistic effect of bacterial isolates against F. oxysporum f. sp. glycines in the dual culture test |
 | |
| *: Indicates percent inhibition of radial growth (PIRG) at 7 days of incubation; Means in the same column with different letter(s) are significantly different at p≤0.05 according to HSD of arcsine transformed values | |
The fungal pathogen was lysed and failed to re-grow when parasitized fungal agar disc by UPM40 cultured on fresh PDA from the interaction and overgrowth zone, whereas parasitized colonies of fungal pathogen by other Trichoderma sp. were shown to re-grow their colonies.
Among bacterial isolates tested, only P. aeruginosa isolate UPM13 B8 showed inhibition with PIRG value of 61.0% against F. oxysporum f. sp. glycines at 7 days of incubation (Table 3 and Fig. 5). Parasitized mycelia of the fungal pathogen were not recoverable on fresh PDA when taken from the inhibition zone.
DISCUSSION
Histopathological studies showed that symptomatic soybean seeds were colonized by F. oxysporum f. sp. glycines on the seed surface and all layers of the seed coat. Results of this study suggested that this fungus was an externally as well as internally seed-borne pathogen and could remain dormant and causing a latent infection without showing any symptom (Sinclair, 1991). However, mycelial growth was more abundant in the hourglass layer. This may be attributed to the large intercellular spaces available that allow for more growth than the closely packed palisade cell layer (Kunwar et al., 1985). The site of infections by this fungus was similarly observed in soybean seeds (Velicheti and Sinclair, 1991). Sharma (1992) stated that this fungus can infect both symptomatic and asymptomatic soybean seeds. Inter- and intracellular mycelia were observed in seed coat and cotyledons. Location of mycelium by fungus in soybean seed tissues depended on the severity of infections, environmental conditions and seed genotypes. Infected tissues of the seed coat seemed to be disintegrated, due to the action of fungal pathogen.
Artificial inoculation of F. oxysporum f. sp. glycines on soybean seeds reduced germination and seedlings survivability confirming pathogenicity to both seeds and seedlings. Infections on seeds caused irregularity in shape and whitish moldy in color. The importance of seed-borne inoculum is in its ability to survive and affect seeds and seedlings at the time of germination (Singh and Mathur, 2004). F. oxysporum f. sp. glycines was confirmed to be an externally and internally seed-borne pathogen leading to local and systemic transmission during germination. Thus, infections caused seeds to rot, with occurrence of pre- emergence damping off in seedlings by progressive rotting of the hypocotyl-radicle axis (Velicheti and Sinclair, 1991).
In vitro screening is considered the most desirable first step to test and select a large number of potential antagonists for possible application in the field. In vitro techniques are also employed later to help identify the mechanisms involved in the biocontrol process (Merriman and Russell, 1990). However, in this study, the in vitro screening procedure was also used for the primary screening of different isolates of Trichoderma and bacteria. Trichoderma harzianum (UPM40) showed the highest potential ability to inhibit the growth of F. oxysporum f. sp. glycines based on higher PIRG values compared to other test isolates. The time needed for colony overgrowth is an important parameter in the assessment of the antagonistic ability to compete against the pathogen for limited nutrient resources and space (Ibrahim, 2005). T. harzianum was able to overgrow fully the fungal pathogen colony and caused lysis within seven days. There was no recovery of F. oxysporum f. sp. glycines from the parasitized mycelia. The inhibition of radial growth of fungal mycelia in this study was considered to be antibiosis, in which cellular degrading enzymes penetrated hyphal cells and inhibited their activity and caused their lyses (Wilson and Wisniewski, 1994; Harman, 2005). T. harzianum was found to be an effective antagonist against F. oxysporum f. sp. cubense in banana wilt pathogen in vitro (Thangavelu et al., 2004).
Among the bacterial isolates tested, Pseudomonas aeruginosa (UPM13B8) was found to inhibit the radial growth of F. oxysporum f. sp. glycines by contact inhibition. Fluorescent pseudomonads isolated from rhizosphere/rhizoplane of crop plants have shown in vitro antibiosis against F. oxysporum f. sp. ciceris of chick-pea (Dileep, 1998). Several mechanisms are responsible for suppression of pathogens by bacteria including competition, antibiotic and metabolite production (Whipps, 2001; Compant et al., 2005). P. aeruginosa is known to produce metabolites such as pyoverdin, pyochelin and salicylic acid which are effective against various pathogens (Buysens et al., 1996; De Meyer and Hofte, 1997). Thus, the activities of T. harzianum and P. aeruginosa in this study suggested that they can be utilized as potential antagonists for use as biological control program for controlling F. oxysporum f. sp. glycines causing soybean seed infections. However, further research is needed to develop good formulation and the most suitable technique for field application.
ACKNOWLEDGMENT
The authors are sincerely grateful to the Third World Organization for Women in Science (TWOWS) for the financial support to conduct this research at Universiti Putra Malaysia.
REFERENCES
- Agarwal, P.C., U. Dev, B. Singh, R. Indra and R.K. Khetarpal, 2006. Seed-borne fungi detected in consignments of soybean seeds (Glycine max) imported into India. Bull. OEPP, 36: 53-58.
Direct Link - Akinsanmil, O.A. and O.K. Adekunle, 2003. Effect of Fusarium oxysporum f. sp. glycines and Sclerotium rolfsii on the pathogenicity of Meloidogyne incognita race 2 to soybean. Plant and Soil, 253: 429-435.
CrossRefDirect Link - Buysens, S., K. Heungens, J. Poppe and M. Hofte, 1996. Involvement of pyochelin and pyoverdin in suppression of Pythium-induced damping-off of tomato by Pseudomonas aeruginosa 7 NSK2. Applied Environ. Microbiol., 62: 865-871.
Direct Link - Compant, S., B. Duffy, J. Nowak, C. Clement and E.A. Barka, 2005. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action and future prospects. Applied Environ. Microbiol., 71: 4951-4959.
CrossRefPubMedDirect Link - De Meyer, G. and M. Hofte, 1997. Salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 induces resistance to leaf infection by Botrytis cinera on bean. Phytopathology, 87: 588-593.
Direct Link - Dileep, K.B.S., 1998. Disease suppression and crop improvement through fluorescent pseudomonads isolated from cultivated soils. World J. Microbiol. Biotechnol., 14: 735-741.
CrossRefDirect Link - Gardener, M.B.B. and D.R. Fravel, 2002. Biological control of plant pathogens research commercialization and application in the USA. Plant Health Progress.
CrossRefDirect Link - Hungria, M., R.J. Campo, I.C. Mendes and P.H. Graham, 2005. Contribution of Biological nitrogen Fixation to the N nutrition of Grain Crops in the Tropics the Success of Soybean Glycine max (L.) Merr.] in South America. In: Nitrogen Nutrition and Sustainable Plant Productivity. Singh, R.P. and N.P.K. Shankar Jaiwal (Eds.), Stadium Press, LLC, Houston, Texas.
- Kloepper, J.W., B. Schippers and P.A.H.M. Bakker, 1992. Proposed elimination of the term endorhizosphere. Phytopathology, 82: 726-727.
Direct Link - Nasir, N., 2003. Detecting seed borne fungi of soybean by different incubation methods. Plant Pathol. J., 2: 114-118.
CrossRefDirect Link - Reyes-Ramirez, A., B.I. Escudero-Abarca, G. Aguilar-Uscanga, P.M. Hayward-Jones and J.E. Barboza-Corona, 2004. Barbozacorona antifungal activity of Bacillus thuringiensis chitinase and its potential for the biocontrol of phytopathogenic fungi in soybean seeds. J. Food Sci., 69: 131-134.
CrossRefDirect Link - Sinclair, J.B., 1991. Latent infection of soybean plants and seeds by fungi. Plant Dis., 75: 220-224.
Direct Link - Sinclair, J.B., 1995. Reevaluation of grading standards and discounts for fungus-damaged soybean seeds. J. Am. Oil Chem. Soc., 72: 1415-1419.
CrossRef - Thangavelu, R., A. Palaniswami and R. Velazhahan, 2004. Mass production of Trichoderma harzianum for managing Fusarium Wilt of banana. Agric. Ecosyst. Environ., 103: 259-263.
CrossRef - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
Direct Link - Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96: 190-194.
CrossRefDirect Link