
Imandi Sarat Babu
Department of Chemical Engineering, Center for Biotechnology, Andhra University, Visakhapatnarn-530003, India
Garapati Hanumantha Rao
Department of Chemical Engineering, Center for Biotechnology, Andhra University, Visakhapatnarn-530003, India
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 88-93







ABSTRACT
The production of extra cellular lipase in submerged fermentation of Yarrowia lipolytica NCIM 3589 has been investigated. Several compounds have been added to the culture medium in order to assess their efficiency as inducers of lipase production. The effect of triglycerides (olive oil, sunflower oil and tributyrin) has been studied. The highest lipase activity was obtained with sunflower oil (2.5 U mL-1) in four days.
PDF Abstract XML References
How to cite this article
Imandi Sarat Babu and Garapati Hanumantha Rao, 2007. Optimization of Process Parameters for the Production of Lipase in Submerged Fermentation by Yarrowia lipolytica NCIM 3589. Research Journal of Microbiology, 2: 88-93.
URL: https://scialert.net/abstract/?doi=jm.2007.88.93
URL: https://scialert.net/abstract/?doi=jm.2007.88.93
INTRODUCTION
Lipases (triacylglycerol acylhydrolases, EC 3.1.1.3) are one of the most important classes of industrial enzymes. They hydrolyse triglycerides into diglyceriods, monoglycerides, glycerol and fatty acids. Lipases occur widely in nature, but only microbial lipases are commercially significant. Commercial preparations of microbial lipases are produced by fermentation of different bacteria, fungi, yeast and actinomycetes (Sharma et al., 2001).
In recent years, there has been an increasing interest in the study of lipases, mainly due to their potential applications as medicines (digestive enzymes), food additives (flavour modifying enzymes), clinical reagents (glyceride-hydrolysing enzymes) and cleaners (detergent additives) (Sugiura, 1984). Additionally, a promising application field for lipases in the biodegradation of plastics such as polyhydroxyalkanoates (PHA) and polycaprolactone (PCL) (Jager et al., 1995; Mochizuki et al., 1995). These enzymes are able to catalyze not only hydrolysis but also synthesis reactions (i.e., esterification, transesterification) in media with low water content (Schmitt-Rozieres et al., 1999; Osorio et al., 2001). More over, they usually retain their structure and activity in organic solvents and at high temperature (Khmelnitsky and Rich, 1999). Also they have several advantages over chemical catalysts: substrate specificity, region- and enatio- selectivity, lower temperature and pressure requirements.
In the present study, several compounds have been added to submerged fermentation of Yarrowia lipolytica NCIM 3589 in order to evaluate their ability to stimulate lipase secretion. The effect of the addition of triglycerides has been studied and the most potent lipase inducer has been selected.
MATERIALS AND METHODS
Microorganism
Yarrowia lipolytica NCIM 3589 obtained from National Chemical Laboratory, Pune, India, was used throughout the study.
Growth Medium and Growth Conditions
The culture was maintained on MGYP slants having the composition (%): malt extract 0.3, glucose 1.0, yeast extract 0.3, peptone 0.5 and agar agar 2.0. The pH of the medium was adjusted to 6.4-6.8 and culture was incubated at 30°C for 48 h. Sub culturing was carried out once in every 2 weeks and the culture was stored at 4°C.
Inoculum Preparation
The yeast Yarrowia strain was cultivated in a medium containing peptone 5 g, yeast extract 3 g and sodium chloride 3 g L-1 of distilled water. The cells were cultivated in this medium at 30°C on a shaker at 200 rpm for 24 h (Oswal et al., 2002).
Production Media
The composition of the basal medium for lipase production was as follows (g L-1): Urea,2; KH2PO4,1; MgSO4.7H2O,0.5; CaCl2,0.1; NaCl,0.1; H3BO3,0.0005; CuSO4.5H2O, 0.00004; KI,0.0001; FeCl3.4H2O,0.0002; ZnSO4.7H2O,0.0004; MnSO4.H2O,0.0004; myo-inositol,0.0000004 and d-biotin, 0.0000008 (Corzo and Revah, 1999). The medium without vitamins was sterilized at 121°C for 15 min. After cooling, the vitamins previously sterilized by filtration were added to the basal medium. A cell suspension of inoculum was added to the basal medium. The flasks were incubated in an orbital shaker at 160 rpm and 30°C for 96 h.
Optimization of Medium Parameters
The strategy adopted was to optimize one particular parameter at a time and then include it at its optimum value in the next optimization step, if found beneficial. The parameters optimized were: (1) carbon level, (2) salt solution concentration, (3) incubation time, (4) temperature, (5) nitrogen level and (6) effect of triglycerides.
Lipase Assay
The activity of lipase was determined as described in the literature (Winkler and Stuckmann, 1979) with the following modifications: 1 mL of isopropanal containing 3 mg of p-nitrophenyl palmitate (pNPP) was mixed with 9 mL of 0.05 M Tris-HCl buffer (pH 8.0), 40 mg of Triton X-100 and 10 mg of gum arabic. Liberation of p-nitrophenol at 28°C was detected at 410 nm. One enzyme unit was defined as 1 μmol of p-nitrophenol enzymatically released from the substrate per minute (Bruno et al., 2004).
RESULTS AND DISCUSSION
Effect of Carbon Source
The carbon sources were selected based on earlier reports that show increase in lipase activity in other microorganisms, such as Beauveria bassiana (Hegedus and Khachatourians, 1988) Rhiozopus oligosporus (Nahas, 1988) Candida and Yarrowia (Novotny, 1988) Humicola lanuginose (Omar et al., 1987) and Acremonium strictum (Okeke and Okolo, 1990). The lipase activity by different carbon sources is presented in Table 1. The lipase activity was not observed on the culture medium containing glycerol because it was an end product of the hydrolysis of triacylglycerols and it might repress lipase induction.
| Table 1: | Effect of the different carbon sources on lipase activity |
 | |
Olive oil and corn oil have increased the yield of extra cellular lipase in microorganisms such as Candida rugosa (Benjamin and Pandey, 1996), Aspergillus niger (Hatzinikolaou et al., 1996) and Aspergillus flavus (Long et al., 1996). Among all the carbon sources, the maximum lipase activity was observed with the glucose. The highest lipase activity in the supernatant of the culture was observed at 2%(w/v) of glucose concentration as shown in Fig. 1. These results were in accordance with the observed lipase production by Yarrowia lipolytica 681 (Corzo and Revah, 1999).
Effect of Salt Solution Concentration
Salt solution of the medium plays a vital role for microbial growth and biochemical activities in submerged fermentation. The maximum yield was obtained at 15% salt solution concentration as shown in Fig. 2. At higher salt solution concentration, lipase production was reduced due to the inhibitory effects of salts.
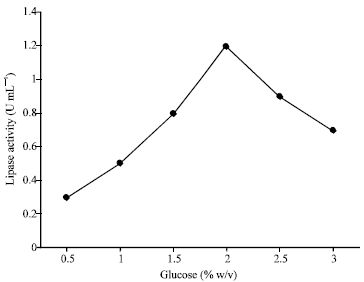 | |
| Fig. 1: | Effect of glucose concentration on lipase activity |
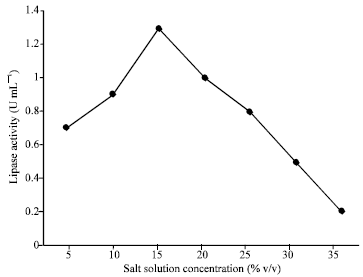 | |
| Fig. 2: | Effect of salt solution concentration on lipase activity |
Effect of Incubation Time
The amount of lipase produced was observed daily during a period of six days. The maximum lipase activity was observed on fourth day as listed in Table 2. After fourth day, it was reduced due to the consumption of nutrient materials.
Effect of Temperature
As yeast strains are very sensitive towards temperature, a slight change in temperature causes reduction in the yield. Yarrowia lipolytica produced maximum amount of lipase at 30°C is presented in Table 3.
Effect of Nitrogen Source
Nitrogen source mediated lipase production was well documented in submerged fermentation (Corzo and Revah, 1999). Table 4 depicts the role of different nitrogen sources on lipase production. Among all the nitrogen sources, urea yielded maximum lipase activity. Lipase production was not detected in the medium containing ammonium sulfate. Urea was the best nitrogen source for increasing lipolytic activity in six different yeast cultures (Novotny et al., 1988).
Effect of Triglycerides
Triacylglycerols and fatty acids have been reported to induce lipase secretion by various microorganisms (Obradors et al., 1993; Berto et al., 1997; Kulkarni and Gadre, 1999). The imperative role of different triglycerides on lipase production by this organism was elucidated by incorporating the selected triglyceride 1.5% (v/v) to the culture medium. The lipase production by different triglycerides are presented in Table 5. Among all the triglycerides, the maximum lipase activity was observed with sunflower oil.
| Table 2: | Effect of incubation time on lipase activity |
 | |
| Table 3: | Effect of temperature on lipase activity |
 | |
| Table 4: | Effect of different nitrogen sources on lipase activity |
 | |
| Table 5: | Effect of different triglycerides on lipase activity |
 | |
CONCLUSIONS
The present study describes the production of extra cellular lipase in submerged fermentation of Yarrowia lipolytica NCIM 3589. The highest lipase activity (2.5 U mL-1) was obtained with the addition of sunflower oil in four days. These results also implied the existence of significant differences between the behavior of Yarrowia lipolytica NCIM 3589 and other wild strains described in the literature.
ACKNOWLEDGMENTS
This research was carried out in the Center for Biotechnology, Department of Chemical Engineering, Andhra University, Visakhapatnam with the financial assistance received from the University Grants Commission (SAP, Phase-III), New Delhi, India.
REFERENCES
- Benjamin, S. and A. Pandey, 1996. Optimization of liquid media for lipase production by Candida rugosa. Bioresour. Technol., 55: 167-170.
Direct Link - Berto, P., L. Belingheri and B. Dehorter, 1997. Production and purification of a novel extracellular lipase from Alternaria brassicicola. Biotechnol. Lett., 19: 533-536.
CrossRefDirect Link - Bruno, L.A., G.A.S. Pinto, H.F. Castro, J.L. Filho and E.H.M. Melo, 2004. Variables that affect immobilization of Mucor miehei lipase on nylon membrane. World J. Microbiol. Biotechnol., 20: 371-375.
Direct Link - Corzo, G. and S. Revah, 1999. Production and characteristics of the lipase from Yarrowia lipolytica 681. Bioresour. Technol., 70: 173-180.
CrossRef - Hatzinikolaou, D., J.B. Macris, P. Christakopoulos, D. Kekos, F.N. Kolisis and G. Fountoukidis, 1996. Production and partial characterization of extracellular lipase from Aspergillus niger. Biotechnol. Lett., 18: 547-552.
Direct Link - Jaeger, K.E., A. Steinbuchel and D. Jendrossek, 1995. Substrate specificities of bacterial polyhydroxyalkanoate depolymerases and lipases: Bacterial lipases hydrolyze poly (omega-hydroxyalkanoates). Applied Environ. Microbiol., 61: 3113-3118.
Direct Link - Khmelnitsky, Y.L. and J.O. Rich, 1999. Biocatalysis in nonaqueous solvents. Curr. Opin. Chem. Biol., 3: 47-53.
Direct Link - Kulkarni, N. and R.V. Gadre, 1999. A novel alkaline thermostable protease-free lipase from Pseudomonas sp. Biotechnol. Lett., 21: 897-899.
Direct Link - Long, K., H.M. Ghazali, A. Ariff, K. Ampon and C. Bucke, 1996. Mycelium-bound lipase from a locally isolated strain of Aspergillus flavus link: Pattern and factors involved in its production. J. Chem. Technol Biotechnol., 67: 157-163.
Direct Link - Mochizuki, M., M. Hirano, Y. Kanmuri, K. Kudo and Y. Tokiwa, 1995. Hydrolysis of polycaprolactone by lipase: Effects of draw ratio on enzymatic degradation. J. Applied Polymer Sci., 55: 289-296.
Direct Link - Novotny, C., L. Dolezalova and J. Lieblova, 1994. Dimorphic growth and lipase production in lipolytic yeasts. Folia Microbiologica Praha, 39: 71-73.
Direct Link - Obradors, N., J.L. Montesinos, F. Valero, F.J. Lafuente and C. Sola, 1993. Effects of different fatty acids in lipase production by Candida rugosa. Biotechnol. Lett., 15: 357-360.
Direct Link - Okeke, C.N. and B.N. Okolo, 1990. The effect of cultural conditions on the production of lipase by Acremonium stricum. Biotechnol. Lett., 12: 747-750.
Direct Link - Omar, I.C., N. Nishio and S. Nagai, 1987. Production of a thermostable lipase by Humicola lanuginose grown on sorbitol-corn steep liquor medium. Agric. Biol. Chem., 51: 2145-2151.
CrossRefDirect Link - Osorio, N.M., S. Ferreira-Dias, J.H. Gusmao and M.M.R. Da-Fonseca, 2001. Response surface modeling of the production of-3 polyunsaturated fatty acids-enriched fats by commercial immobilized lipase. J. Mol. Catal B., 11: 677-686.
Direct Link - Oswal, N., P.M. Sarma, S.S. Zinjarde and A. Pant, 2002. Palm oil mill effluent treatment by a tropical marine yeast. Bioresour. Technol., 85: 35-37.
CrossRefPubMedDirect Link - Schmitt-Rozieres, M., G. Vanot, V. Deyris and L.C. Comeau, 1999. Borago Officinalis oil: Fatty acid fractionation by immobilized Candida rugosa lipase. J. Am. Oil. Chem. Soc., 76: 557-562.
Direct Link - Sharma, R., Y. Chisti and U.C. Banerjee, 2001. Production, purification, characterization and applications of lipases. Biotechnol. Adv., 19: 627-662.
CrossRef - Winkler, U.K. and M. Stuckmann, 1979. Glycogen, hyaluronate and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol., 138: 663-670.
Direct Link