
Yasser R. Abdel-Fattah
Department of Bioprocess Development,
Genetic Engineering and Biotechnology Research Institute, Mubarak City for Scientific Research and Technology Applications, New Burg El-Arab City 21934, Alexaudria, Egypt
Ehab R. El-Helow
Department of Botany, Faculty of Science, Alexandria University, Egypt
Khaled M. Ghanem
Department of Botany, Faculty of Science, Alexandria University, Egypt
Walid A. Lotfy
Department of Botany, Faculty of Science, Alexandria University, Egypt
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 13-23







ABSTRACT
Statistically based experimental designs were applied to optimize the production of avicelase by a recently isolated endospore forming strain, WL1, that showed 99.8% 16S rDNA identity to Geobacillus stearothermophilus. Seven culture parameters were examined for their significance as effectors of avicelase expression using the Plackett-Burman factorial design. Concentrations of avicel, yeast extract and ammonium sulfate were the most significant factors affecting the process of enzyme production. The second optimization step was to figure out the levels of these three independent variables that generate maximum avicelase activity, using the Box-Behnken design. Maximal enzyme activity (0.8 U mL-1), which is approximately two folds the activity expressed in the basal medium, has been predicted at concentrations of (g L-1): Avicel (42), yeast extract (3.6) and ammonium sulfate (0.8). A verification experiment was accomplished and revealed approximately 99% model validity.
PDF Abstract XML References
How to cite this article
Yasser R. Abdel-Fattah, Ehab R. El-Helow, Khaled M. Ghanem and Walid A. Lotfy, 2007. Application of Factorial Designs for Optimization of Avicelase Production by a Thermophilic Geobacillus Isolate. Research Journal of Microbiology, 2: 13-23.
URL: https://scialert.net/abstract/?doi=jm.2007.13.23
URL: https://scialert.net/abstract/?doi=jm.2007.13.23
INTRODUCTION
Cellulose, an unbranched glucose polymer composed of anhydro-β-1,4-glucose units linked by β-1,4-glycosidic bonds, is the most abundant biopolymer on the earth. Enzymatic degradation of cellulose, also known as avicel, to glucose requires the co-operative action of at least an endo-1,4-β-glucanase (Avicelase, EC 3.2.1.4), exo-1,4-β glucanase (EC 3.2.1.91) and b-glucosidase (EC 3.2.1.21) (Wood, 1989).
Avicelases were found to have potential applications in the bioconversion of agricultural waste materials to useful products, such as single cell protein, fuels and chemical feed stocks (Kari et al., 1994; Nikolay et al., 1998). A diversity of microorganisms has been shown to produce enzymatic systems capable of degrading cellulosic materials (Spear et al., 1993; Nikolay et al., 2000).
Bacillus species continue to be dominant bacterial workhorses, due to the capacity of some selected species to produce and secrete large quantities of extracellular enzymes (Schallmey et al., 2004). Two distinct avicelase activities in B. circulans were observed (Kim, 1995). Other Bacillus strains belonging to species such as B. sphaericus and B. subtilis express high cellulose degradation activities (Takami and Horokoshi, 2000; Singh et al., 2004). Recently, a thermophilic aerobic bacterium capable of avicelase production, Caldibacillus cellulovorans, has been isolated (Huang and Monk, 2004).
In general, evaluation of nutritional and environmental requirements of the microorganism is an important step for bioprocess development. Optimization studies involving the one-factor-at-a-time approach are not only tedious but also tend to overlook the effects of interacting factors and might lead to misinterpretations of the results. On the other hand, statistically planned experiments effectively solve such problems and minimize the error in determining the effect of parameters and the results are achieved in an economical manner (Abdel-Fattah et al., 2005).
The objective of the present study was to elaborate the best conditions for the production of a thermostable extracellular avicelase by a novel spore former isolate. To the best of our knowledge, this is the first report on production of avicelase by G. stearothermophilus. Optimization of the bioprocess was carried out through a step-wise experimental strategy including: (1) elucidation of medium and environmental aspects that affect enzyme production significantly using a two-level multi-factorial screening design (Plackett and Burman, 1946), (2) optimization of the most significant components and generating a mathematical model expressing the relationship between optimized factors and avicelase production by application of the Box-Behnken experimental design (Box and Behnken, 1960) and (3) verification of the model and monitoring the production pattern. For a more economic bioprocess, cheaper cellulytic carbon sources were tested alternative to avicel.
MATERIALS AND METHODS
Microorganism
The bacterium used throughout this study is a new Egyptian soil isolate. Soil samples were suspended in sterile water in a ratio of 1:1. Three sequential enrichment steps were carried out by inoculating 1 mL of the soil suspension to yeast peptone medium supplemented with 1% carboxymethyl cellulose (CMC) under shaking conditions at 55°C. An inoculum from each enrichment step was streaked onto agar plates of the same medium for further purification. After 24 and 48 h of incubation at 50°C, clear zones were developed by spreading iodine solution on the surface of each culture. A single isolated colony was selected and the bacterium was identified by 16S rRNA gene sequencing, as previously described (El-Helow, 2001) and blasting the results in the GenBank.
Enzyme Production Conditions
Cultures were allowed to grow in 250 mL Erlenmeyer flasks containing 25 mL aliquots of fermentation basal medium of the following composition (g L-1): Avicel, 20; yeast extract, 0.5; (NH4)2SO4, 2; MgCl2.6H2O, 0.2; CaCl2, 0.1; K2HPO4, 2 and KH2PO4, 0.6 (Malfait et al., 1984). Cultures were incubated at 55°C under shaking (250 rpm) or stagnant. Standard overnight seed cultures, prepared in nutrient broth, were used to inoculate fermentation cultures as 1% inoculum. Bacterial growth was monitored turbidimetrically. Turbidity readings were converted to dry weights using a standard curve. Samples were collected after indicated incubation times and cells were isolated by centrifugation at 10,000 rpm for 10 min and the supernatants were separated and stored at -20°C for few hours until the necessary analyses were carried out.
Enzyme Assay
A solution of 0.125% avicel (from Cetraria Islandica-Sigma, USA) dissolved in phosphate buffer of pH 5 was used as a substrate. The reaction was performed by adding 0.5 mL substrate to 0.5 mL diluted enzyme and incubation in a water bath at 55°C for half an hour. The reducing sugars produced were measured by the di-nitrosalisylic acid method (Miller, 1959). An enzyme activity unit is defined as the amount of enzyme producing 1 mg mL-1 reducing sugars (glucose equivalent) 1 h-1.
| Table 1: | Plackett-Burman experimental design for elucidation of factors affecting avicelase production |
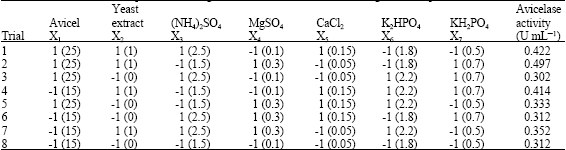 | |
| Natural levels of variables (g L-1) are presented between brackets | |
Statistical Designs
Plackett-Burman Design
As a preliminary optimization experiment, various medium components and environmental factors have been evaluated. Based on the Plackett-Burman factorial design, each factor was examined in two levels: -1 for a low level and 1 for a high level (Plackett and Burman, 1946). Table 1 shows the factors under investigation as well as levels of each factor used in the experimental design.
The Plackett-Burman experimental design is based on the first order model:
Y = β0 + ∑βIxi | (1) |
Where Y is the response (avicelase activity), β0 is the model intercept, βI is the linear coefficient and xi is the level of the independent variable. In the present study, seven assigned variables were screened in eight experimental designs. All experiments were carried out in duplicates and the averages of avicelase activity results were taken as the response. Variables whose confidence levels were 90% or higher were treated as significant variables.
Box-Behnken Design
In order to describe the nature of the response surface in the experimental region, a Box-Behnken design (Box and Behnken, 1960) was applied. Table 3 represents the design matrix of a 13 trials experiment. Using this design, factors of highest confidence levels were prescribed into three levels, coded –1, 0 and +1 for low, middle and high concentrations (or values), respectively. For predicting the optimal point, a second order polynomial function was fitted to correlate relationship between independent variables and response (avicelase activity). For three factors the equation is:
Y = β0 + β1 X1 + β2 X2 + β3 X3 + β12 X1X2 + β13 X1X3 + β23 X2X3 + β11 X12 + β22 X22 + β33 X32 | (2) |
Where Y is the predicted response, β0 is the model constant; X1, X2 and X3 are independent variables; β1, β2 and β3 are linear coefficients; β12, β13 and β23 are cross product coefficients and β11, β22 and β33 are the quadratic coefficients.
Experiments were performed in triplicates and mean values are given. Microsoft Excel was used for the regression analysis of the experimental data obtained. The fitting quality of the polynomial model equation was expressed by the coefficient of determination R2. The optimal value of enzyme activity was estimated using the solver function of Microsoft Excel tools.
RESULTS AND DISCUSSION
Isolation and Characterization of a Novel Avicelase Producing Bacterium
Based on clear zone diameters on carboxy methyl cellulose (CMC) supplemented plates, the isolate WL1 was selected out of a group of purified bacterial colonies. Microscopic examinations revealed that the bacterium is a Gram-positive rod-shaped endospore former.
| Table 2: | Statistical analyses of the Plackett-Burman experimental results |
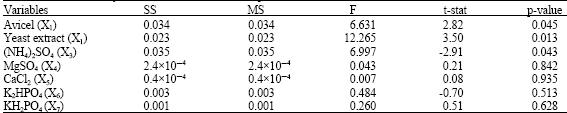 | |
| Table 3a: | Natural values of variables examined in the two sequential optimization trials |
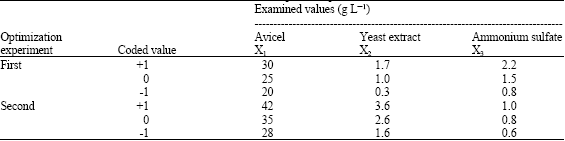 | |
| Table 3b: | The Box-Behnken factorial experimental design |
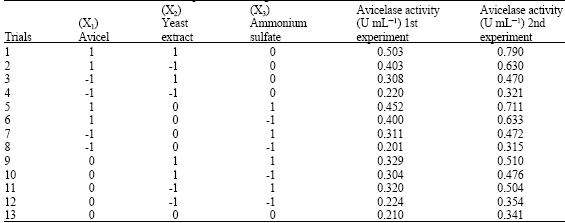 | |
For molecular characterization, genomic DNA of the WL1 isolate was prepared, purified and used for PCR amplification of the gene encoding 16S rRNA using universal primers. Double-strand nucleotide sequences of two fragments (536 and 746 base pairs) were determined and edited. Alignment of these sequences (accession numbers AY 738088 and AY 738089), using Biology WorkBench, showed 99.8% identity to the homologous fragments of G. stearothermophilus. To the best of our knowledge, it is the first time to investigate the celluolytic activity of this species.
Using the basal enzyme production medium, preliminary studies were carried out on WL1 with respect to culture initial pH, temperature and shaking. Enzyme assay results of these experiments revealed that the bacterium expresses a promising level of avicelase activity (0.425 U mL-1) in a shake culture with an initial pH of 7 and an incubation period of 16 h at a temperature of 55°C. Under this culture condition, a maximum specific growth rate (0.94 h-1) was recorded within 24 h of incubation. In a parallel study (data not shown), the partially purified enzyme showed high thermal stability, where it retained 80% of its activity when exposed to 60°C for three hours. The enzyme lost 50% of its activity at 80°C. Optimum enzyme temperature and pH were 60°C and 6.0°C, respectively.
Evaluating the Significance of Medium Constituents
To test the significance of seven culture variables at a time on avicelase formation by the isolate, The Plackett-Burman experimental design was applied. Table 2 represents the design matrix together with the response observations.
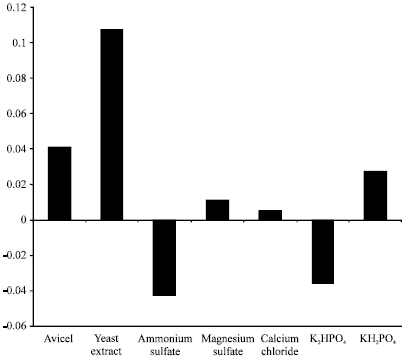 | |
| Fig. 1: | Effect of nutritional factors on avicelase activity produced by G. stearothermophilus in shaked culture at 55°C based on results of Plackett-Burman design |
The linear polynomial model describing the correlation between examined variables and expressed avicelase activity could be presented as follows:
Y= 0.368 + 0.021 X1 + 0.053 X2 – 0.021 X3 + 0.006 X4 + 0.002 X5 – 0.018 X6 + 0.013 X7. |
Based on the analysis of regression coefficients of the seven variables as well as the calculated main effect figures presented graphically in Fig. 1, avicel, yeast extract, magnesium chloride, calcium chloride and dipotassium hydrogen phosphate showed positive effects on the expression of avicelase. On the other hand, ammonium sulfate and potassium dihydrogen phosphate contributed negatively. As shown in Table 2, statistical analyses of the results suggest that yeast extract is the most significant factor affecting avicelase expression by WL1.
The presence of yeast extract as a nitrogen source in the medium stimulates the formation of readily metabolizable compounds that can be utilized for growth and avicelase formation (Mawadza et al., 1996). On the other hand, low ammonium sulfate concentrations and high avicel levels, within the tested constrains, also enhanced the expression of avicelase. In accordance, it has been demonstrated by Bok et al. (1998) that low ammonium sulfate and high avicel levels stimulate the activities of the promoters controlling cellulase genes in Thermotoga neapolitana.
According to the results of this experiments, a medium of the following composition (g L-1) is predicted to be near optimum: avicel, 25; yeast extract, 1; (NH4)2SO4, 1.5; MgCl2.6H2O, 0.3; CaCl2, 0.15; K2HPO4, 1.8 and KH2PO4, 0.7. A verification experiment demonstrated that the enzyme activity expressed in this medium was increased by 1.18 fold (0.5 U mL-1) when compared to the basal condition.
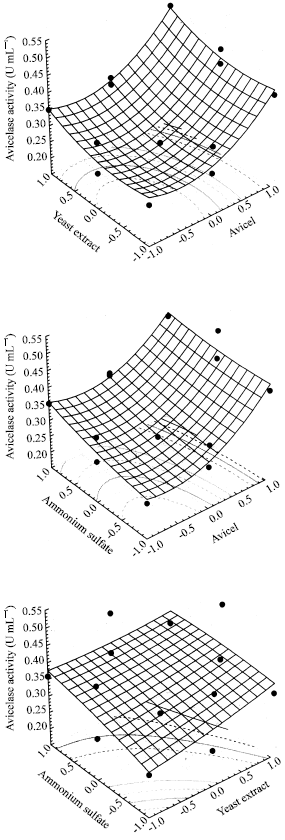 | |
| Fig. 2: | Avicelase activity (U mL-1) response surface from G. stearothermophilus as affected by culture conditions (first optimization set) |
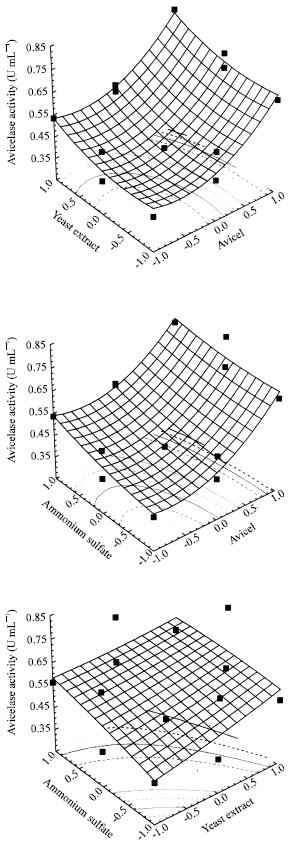 | |
| Fig. 3: | Avicelase activity (U mL-1) response surface from G. stearothermophilus as affected by culture conditions (second optimization set) |
Optimization of Avicelase Production
In this phase of optimization, the most effective variables were further investigated according to the Box-Behnken factorial design (Box and Behnken, 1960). The variables applied were; avicel (X1), yeast extract (X2) and (NH4)2SO4 (X3). Levels of the other medium components were selected based on their main effect signs and treated as constant factors throughout the optimization experiment. Table 3 represents the design matrix, which comprises thirteen trials, accompanied with avicelase activity in two sequential optimization sets with different X1, X2 and X3 levels.
As presented graphically in Fig. 2, the correlation presents a saddle where optimal value could not be found. However, this optimization step reflected an image how the response could be increased and as a result, other levels chosen in the direction of increasing the response were also examined. The second optimization set was run according to the design matrix shown in Table 3.
Again, the response observations presented graphically in Fig. 3, demonstrate a saddle formation. Further application of other levels chosen in the direction of increasing the response in a third experiment decreased the activities when compared to the responses obtained in the second optimization set. Accordingly, the experimental results of the second optimization set were handled and fitted by regression to a polynomial equation as follows:
Y2 = 3.4-0.17 X1– 0.21 X2– 0.87 X3–0.014 X1X3–0.15 X2X3+0.003 X12+0.07 X22+1.25 X32 |
The quality of the fit of this polynomial model equation was expressed by R, the multiple correlation coefficient and R2, the coefficient of determination. The closer the value of R to 1, the better is the correlation between the observed and the predicted values. In the second optimization experiment, the value of R = 0.99. This value indicates high degree of correlation between the experimental and the predicted values. The value of determination coefficient R2 = 0.98, being a measure of goodness of fit of the model, indicates that about 2% of the total variations are not explained by the activity models.
The optimal levels of the three examined independent variables as predicted from the model are (g L-1): Avicel, 42; yeast extract, 3.6 and ammonium sulfate, 0.8 with a predicted avicelase activity of 0.79 (U mL-1). Although, otherwise, Krishna (1999) reported that cellulose production in Bacillus subtilis CBTK 106 increases by increasing the concentration of ammonium sulfate, it is worthwhile to mention that the increment was not significant. On the other hand, Enari and Markenan (1977) showed that good production of cellulase was obtained with organic nitrogen sources like yeast extract and peptone.
In order to verify the optimization results, an experiment was carried out under the predicted optimal condition, where the dry weight and avicelase activity were monitored at different time intervals. The basal culture medium was used as a control. As shown in Fig. 4, the optimized medium recorded a relatively higher exponential growth rate. With respect to enzyme expression this formula showed an actual maximum activity of 0.8 U mL-1, which is 101% of the predicted value with a 1.89 fold increase when compared to the pre-optimized medium control culture. The great similarity between predicted and observed results in this application and others (Bloor and England, 1991; Teruel et al., 1997; Kimmel et al., 1998) confirms the high accuracy and applicability of the Box-Behnken model in the optimization process.
It is also likely that the optimized culture condition accelerated the rate of avicelase expression in WL1 as it showed maximum enzyme activity within 12 h instead of 16 h in the case of the control culture. According to these results, a medium of the formula (g L-1): avicel, 42; yeast extract, 3.6; (NH4)2SO4, 0.8; MgCl2.6H2O, 0.3; CaCl2, 0.15; K2HPO4, 1.8 and KH2PO4, 0.7 is suggested for efficient production of avicelase by G. stearothermophilus WL1.
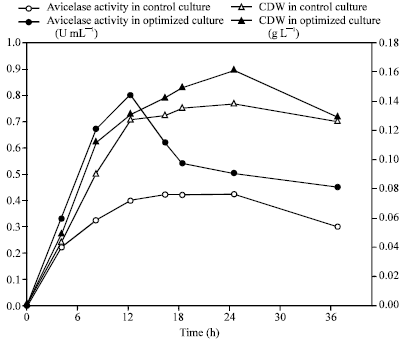 | |
| Fig. 4: | Growth (Δ) and avicelase activity (Ο) of G. stearothermophilus on control culture and growth (▲) and avicelase activity (●) in the optimized culture at optimum concentrations as obtained from the validation experiment |
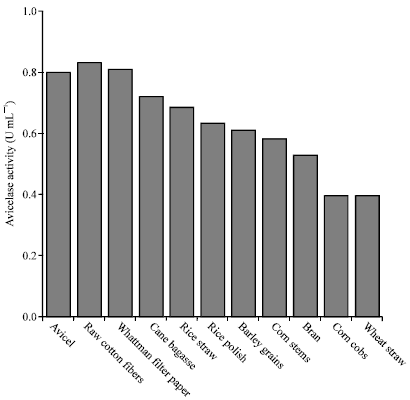 | |
| Fig. 5: | Effect of different cellulosic substrates on the expression of avicelase by G. stearothermophilus |
This optimized medium contains an avicel concentration of more than twice that in the basal medium suggesting that avicelase expression in this bacterium is possibly regulated by induction.
Effect of Different Cellulosic Substrates on Avicelase Production by G. stearothermophilus
An important fermentation consideration is the cost of the carbon source, which contributes significantly to the overall production costs (Li et al., 1998). Also, it has been reported that the type of cellulose has an important role for the production of active cellulases (Li et al., 1997). Accordingly, a group of low cost cellulosic materials were individually examined to replace avicel in the optimized medium. The tested substrates namely; raw cotton fibers, Whattman filter paper No.4, cane bagasse, rice straw, rice polish, barely grains, corn stems, bran, corn cobs and wheat straw, were added separately, at carbon equivalent weights, to the optimized medium formula. Cultures were incubated for 12 h at 55°C under shaking conditions. Clear culture filtrates recorded enzyme assay results that ranged between 0.40 up to 0.83 U mL-1. The observed differences in enzyme production levels could be attributed to structural factors, which affect the expression of individual enzymes of the total cellulase enzyme system (Krishna, 1999). As shown in Fig. 5, the most promising substrates are raw cotton fibers and filter papers, which attained avicelase activities of 104 and 101.3% of the optimized pure avicel culture, respectively.
REFERENCES
- Abdel-Fattah, Y.R., H.M. Saeed, Y.M. Gohar and M.A. El-Baz, 2005. Improved production of Pseudomonas aeruginosa uricase by optimization of process parameters through statistical experimental designs. Process Biochem., 40: 1707-1714.
CrossRefDirect Link - Bloor, S. and R.R. Ebgland, 1991. Elucidation and optimization of the medium constituents controlling antibiotic production by cyanobacerium Nostoc muscorum. Enzyme. Microb. Technol., 13: 76-80.
Direct Link - Bok, J.D., D.A. Yernool and D.E. Eveleigh, 1998. Purification, characterization and molecular analysis of thermostable cellulases celA and celB from Thermotoga neapolitana. Applied Environ. Microbiol., 64: 4774-4781.
Direct Link - Box, G.E.P. and D.W. Behnken, 1960. Some new three level designs for the study of quantitative variables. Technometrics, 2: 455-475.
CrossRefDirect Link - El-Helow, E.R., 2001. Identification and molecular characterization of a novel Bacillus strain capable of degrading Tween-80. FEMS Microbiol. Lett., 196: 119-122.
Direct Link - Enari, T.M. and P. Markenan, 1977. Production of cellulolytic enzymes by fungi. Adv. Biochem. Eng., 5: 1-24.
CrossRefDirect Link - Huang, X.P. and C. Monk, 2004. Purification and characterization of cellulase (CMCase) from a newly isolated thermophilic aerobic bacterium Caldibacillus cellulovorans gen. nov., sp. nov. World J. Microbiol. Biotechnol., 20: 85-92.
Direct Link - Kari, A., R. Flengsrud, V. Lindahl and A. Tronsmo, 1994. Characterization of production and enzyme properties of an endo-β-1,4-glucanase from Bacillus subtilis ek-2 isolated from compost soil. Antonie Van Leeuwenhoek, 66: 319-326.
Direct Link - Kim, C.H., 1995. Characterization and substrate specificity of an endo-β-1,4-glucanase I (Avicelase I) from an extracellular multienzyme complex of Bacillus circulans. Applied Environ. Microbiol., 61: 959-965.
Direct Link - Kimmel, S.A., R.F. Roberts and G.R. Ziegler, 1998. Optimization of exopolysaccharide production by Lactobacillus delbrueckii sub sp. Bulgaricus RR grown in a semidefined medium. Applied Environ. Microbiol., 64: 659-664.
Direct Link - Krishna, C., 1999. Production of bacterial cellulases by solid substrate bioprocessing of banana wastes. Bioresour. Technol., 69: 231-239.
CrossRefDirect Link - Li, X.L., H. Chen and L.G. Ljungdahl, 1997. Two cellulases, celA and celC, from the polycentric fungus Orpinomyces strain PC-2 contain N-terminal docking domains for a cellulase-hemicellulase complex. Applied Environ. Microbiol., 63: 4721-4728.
Direct Link - Li, X., W. Lin, P. Gao and F. Chen, 1998. Endoglucanase S., a novel endocellulase exhibiting exocellulase activity from a newly isolated Streptomyces sp. LX. J. Applied Microbiol., 85: 347-356.
Direct Link - Mawadza, C., F.C. Boogerd, R. Zvauya and W. Henk, 1996. Influence of environmental factors on endo-β-1,4-glucanase production by Bacillus HR 68, isolated from a Zimbabwean hot spring. Antonie Van Leeuwenhoek, 69: 363-369.
Direct Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Nikolay, S.A., K. Robson and D.B. Wilson, 1998. Regulation of biosynthesis of individual cellulases in Thermomonospora fusca. J. Bacteriol., 180: 3529-3532.
Direct Link - Nikolay, S.A., K. Robson and D.B. Wilson, 2000. A celR mutation affecting transcription of cellulase genes in Thermobifida fusca. J. Bacteriol., 182: 252-255.
Direct Link - Plackett, R.L. and J.P. Burman, 1946. The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
CrossRefDirect Link - Schallmey, M., A. Singh and O.P. Ward, 2004. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol., 50: 1-17.
CrossRefPubMedDirect Link - Singh, J., N. Batra and R.C. Sobti, 2004. Purification and characterization of alkaline cellulose produced by a novel isolate, Bacillus sphaericus JS1. J. Ind. Microbiol. Biotechnol., 31: 51-56.
Direct Link - Spear, L., J. Gallagher, L. Mchale and A.P. Mchale, 1993. Production of cellulase and β-glucosidase activities following growth of Streptomyces hygroscopicus on cellulose containing media. Biotechnol. Lett., 15: 1265-1268.
Direct Link - Takami, H. and K. Horokoshi, 2000. Analysis of the genome of an alkalophilic Bacillus strain from an industrial point of view. Extremophiles, 4: 99-108.
Direct Link - Teruel, M.L., E. Gontier, C. Bieneime, J.E.N. Saucedo and J.N. Barbotin, 1997. Response surface analysis of chlortetracycline and tertracycline production with k-carrageenan immobilized Streptomyces aureofaciens. Enzyme. Microb. Technol., 21: 314-320.
Direct Link