
A.H. Mohagheghi Fard
Department of Virology, School of Medicine,
Zahedan Medical University, Zahedan, Iran
P. J. Vallely
Department of Virology, School of Medicine, Manchester University, Manchester, UK
Research Journal of Microbiology
Year: 2006 | Volume: 1 | Issue: 6 | Page No.: 467-479







ABSTRACT
We firstly designed an in-house avidine-biotin indirect ELISA for this we grew AD169 strain on HEL cells for antigen isolation and different steps of the ELISA were standardized by checkerboarding. The results were calculated by cut-off and standard binding ratio (SBR). Interassay and intrassay were attempted by testing different sets of sera. Moreover, 556 samples from hospitalized children and 213 samples from leukemic children were assayed by newly developed ELISA. In order to define truly positive samples for HCMV IgG, avidity assay was employed by using 20, 35 and 70 mM Diethylamine (DEA). The comparison of the results by in-house ELISA with those of commercial kits was quite concordant. The ELISA gave more positive results than the RIA (p<0.001). The same conclusion was observed for commercial kits as well. When the sera were subjected to avidity assay positive samples virtually showed high index. Differentiation of seropositivity from seronegativity for HCMV IgG plays a major role in distinguishing subsequent primary from recurrent HCMV infection. Accurate identification of HCMV seropositivity is also crucial in identifying blood, organ or sperm donors who may transmit HCMV infection.
PDF Abstract XML References Citation
How to cite this article
A.H. Mohagheghi Fard and P. J. Vallely, 2006. Development of an In-house ELISA for Detection of HCMV IgG and Finding of Antibody Avidity by Application of Diethylamine. Research Journal of Microbiology, 1: 467-479.
URL: https://scialert.net/abstract/?doi=jm.2006.467.479
URL: https://scialert.net/abstract/?doi=jm.2006.467.479
INTRODUCTION
Human cytomegalovirus (HCMV) is a member of herpesviridae family (β-herpesvirinae subfamily) and is highly linked to its natural host, human beings. The virion structure of HCMV is typical of herpesviruses with the largest DNA genome among these viruses i.e., 230-240 Kbp (Mocarski, 1993). HCMV is highly specie specific and can cause primary and recurrent (or persistent) infections are seen, the latter mostly caused by reactivation of latent virus though, due to antigenic diversity of HCMVs reinfection might occur. There are infections of normal or immunocompromised hosts, congenital and prenatal infections.
HCMV is estimated to infect approximately 70% of individuals living in hygienic conditions, but up to 90% of people living in poor socio-economic circumstances (Griffiths, 1995). Primary HCMV infection is indisputably generalized and portals of entry may include blood (transfusion), oral or congenital contact or possibly genital contact alone (Alford and Britt, 1993). As a result of primary infection, viraemia is detectable for a few weeks or months (Klemola and Kaariainen, 1965; Jordan et al., 1973) and virus is constantly recovered from urine or even pharynx. Viruria may last for one year or more, even though symptoms disappear besides salivary gland involvement is common and possibly chronic, the disease is an infectious mononucleosis-like syndrome (Klemola, 1973; Alford and Britt, 1993).
All antibody classes including IgG, IgM and IgA specific for HCMV have been detected in primary, convalescent and recurrent infections (Doerr et al., 1987). IgG antibodies peak during the first month or two after the onset of infection with two predominant subclasses, IgG1 and IgG3 (Linde et al., 1983; Doerr et al., 1987).
Primary HCMV infections in immunosuppressed (immunocompromised) people are much more severe than in normal hosts (Jawetz et al., 2001). The individuals who are at high risk include those receiving organ transplants, those with malignancies who receive chemotherapy especially AIDS victims. In this group of patients viral shedding is increased, prolonged and disseminated frequently. The immunological status of immunocompromised hosts is interesting because their humoral immune response usually remains intact in spite of immunosuppression (O'Neill et al., 1988) and the antibody response is similar to that of primary infections of normal hosts and detectable by most serological tests (Alford and Britt, 1993).
Immaturity is another pathogenic factor in the expression of HCMV infection hence, disease occurs more frequently and is more severe with foetal infections (Griffiths, 1995). Nevertheless, a vast number of immunodiagnostic tests have been developed to detect HCMV antibodies. IgG antibodies to HCMV develop within 1 to 2 weeks after the onset of an acute primary illness and persist at detectable levels for months to years (Drew, 1988).
ELISA has proved to be a reliable assay for HCMV antibodies (Hopson et al., 1992) therefore, ELISAs are attractive, compared with old serologic assays because ELISA-based assays are sensitive, specific, inexpensive and easily automated (Nielsen et al., 1988). Monoclonal antibody and indirect ELISA have been used for detection of HCMV IgG antibodies in serum (Chou and Scott, 1990; Aulitzky et al., 1991; Tackaberry et al., 1992) or even in urine (Yamanaka et al., 1992).
In this project too, we tried to design and create an in-house indirect avidin-biotin based ELISA as current technique of RIA has certain limitations due to use of radioactive materials. Furthermore, application of diethylamine in different concentrations is an effective tool to determine avidity index of HCMV IgG antibodies. The latter also helps to differentiate primary from remote and recurrent infections. Detection of low avidity specific IgG antibody generally indicates that primary infection has occurred within the last one or two months. Avidity assay have been widely used to differentiate between primary and recurrent infections in case of different viral infections (Thomas, 1995; Soderlund et al., 1995; Gut et al., 1995).
MATERIALS AND METHODS
Antigen Production From Infected and Uninfected Cells
In order to design an in-house avidine-biotin indirect ELISA, proper antigen had to be produced for this we grew AD169 strain on human embryo lung (HEL) cells (<16 passage). To assay the infectivity of the virus, plaque assay was not applied since it took 7-10 days to observe CPE therefore tissue culture infectious dose (TCID50) quantitation was carried out and subsequent steps of the ELISA were standardized and optimized by checkerboarding. Furthermore, in order to proceed a proper ELISA the control antigen was produced from HEL cells. In case of modification with respect to antigen and antibody concentrations, blocking buffer, the optimal dilution of monoclonal antibody (anti-human IgG), conjugate and finally washing of the plates after coating with antigen and after antibody binding and so on were all adjusted and chosen accordingly.
Standard ELISA Protocol
The wells of a microtitre plate (Falcon 3912 Micro Test III) were coated with 140 μL optimally diluted antigen in carbonate bicarbonate buffer (pH: 9.6) and then incubated overnight at 4°C in a moist chamber. The next day the plate was washed with wash buffer 5 times with the same volume i.e., 140 μL (PBS-A with 0.05% tween 20 (Sigma No 5362) and each well was incubated with 140 μL of blocking buffer (1 g skimmed milk powder dissolved in 100 mL of PBS-A) in moist chamber for an hour and later washed 5 times with wash buffer.
In the next step 140 μL of optimally diluted serum in serum diluent (blocking buffer plus 0.5% W/V tween 20) was added to each well. Of course two wells for each serum dilution and on every plate was used as blank (no serum only diluent). The plate was then incubated over night in moist chamber at 37°C. The next day plate was washed according to optimized number of washes i.e., x15. Afterward 140 μL of optimally diluted anti-human IgG mouse monoclonal antibody (biotin labeled; Sigma product No: B-3773) in diluent, which was exactly as blocking buffer, was added to appropriate wells. Incubation period was then half an hour at 37°C in moist chamber. The plate was again washed 5 times and incubated for 5 min at 37°C before addition of 140 μL of optimally diluted avidine peroxidase (Sigma product no: A-3151) in suitable diluent (100 mL PBS-A plus 4.0 g bovine serum albumin and 4.0 g goat serum supplied by Sara Lab Ltd, Sussex, U.K). After incubation and washing of plate 3 times 140 μL of o-phenylenediamine (OPD) dihydrochloride (Sigma product no: P-8287) 10 mg mL-1 dissolved in citrate buffer (pH: 5.0) was added to the wells. The plate was then incubated at 3°C away from light in moist chamber for half an hour. In final step 50 μL stop solution (1N, H2SO4) was added to the wells and the absorbance was read at 492 nm wave length against mean of column blank.
IgG Avidity Assay
As explained earlier, IgG avidity assays plays an important role in distinguishing primary from other categories of viral infections, so as one of the objectives in this project was measurement of IgG avidity (also used as a measure of denaturant-induced dissociation of binding) the approach was as following:
Diethylamine (DEA) (Sigma product No: D-31) in different concentrations (20, 35 and 70 mM) were chosen as IgG denaturants. Standard ELISA was carried out for this purpose except that after antigen-antibody binding, the plate was washed 15 times with wash buffer containing one of the denaturant instead of wash buffer alone. In order to measure IgG avidity it is essential to determine OD of the sample both in standard ELISA and in the avidity assay procedure. Therefore for every batch of samples different plates had to be prepared as:
| • | Plate coated with viral antigen |
| • | Plate coated with control antigen (uninfected cells) |
| • | Plates coated with viral antigen but subjected to avidity assay by adding required denaturants |
Calculation of ELISA Results
The results were calculated by cut-off and Standard Binding Ratio (SBR). In the cut-off method of results calculation a serum was considered positive for IgG antibodies, if the mean of the test serum OD exceeded by more than three standard deviations the mean OD given by 12 replicate test of a negative control serum. Whereas in the SBR method of results calculation a serum was regarded positive if the SBR (mean test serum OD with viral antigen/ mean test serum OD with cell control antigen) was greater than 2 (Klapper et al., 1990a).
Calculation of IgG avidity
Once the OD of the samples from different plates were known as mentioned above IgG avidity was measured by following equation.
Interassay and intrassay were attempted by testing a set of 105 sera already collected on the day of transplantation from renal allograft recipients and were tested for HCMV IgG by RIA now examined using the optimized ELISA. Furthermore, 30 sera of organ donors previously tested in a commercial ELISA (Biokit, Barcelona, Spain) were also used to assess the reproducibility of the in-house ELISA in three successive runs. Moreover, 556 samples from hospitalized children and 213 samples from leukemic children were assayed by in-house ELISA. In order to define truly positive samples for HCMV IgG, avidity assay was employed by using 20, 35 and 70 mM Diethylamine (DEA).
RESULTS
Intra-assay Reproducibility of Optimized HCMV IgG ELISA
The intra-assay coefficient of variation (CV) was calculated by the following equation: Coefficient of variation (CV%) = standard deviation (SD)÷Mean(Xn-1)x100
In an initial experiment using 12 replicates, the intra-assay CVs for the optimized HCMV IgG ELISA were 6, 13 and 15% with strong positive (S +ve), weak positive (W +ve and negative (-ve) sera. In the second experiment using 8 replicates the same CVs were 12, 32 and 54%.
Comparison of HCMV IgG ELISA and RIA
One hundred and five serum samples collected on the day of transplantation from renal allograft recipients had been used for HCMV IgG by RIA in a previous study in 1989. The samples stored in the interim at -30°C were now examined using the optimized HCMV IgG ELISA. With the ELISA the proportion of serum specimens giving positive results was greater with the cut-off rather than ratio method of results calculation [86/105 (82%) versus 75/105 (71%), p = 0.05]. With the RIA no difference was seen [44/105 (42%) versus 43/105 (41%)]. The ELISA gave more positive results than the RIA irrespective of whether the results were determined using cut-off [86/105 (82%) versus 44/105 (42%), p<0.001] or SBR calculation [75/105 (71%) versus 43/105 (41%), p<0.001]. In both assays the ratio were higher for sera positive rather than negative by cut-off (Table 1). Specimens positive by binding ratio but negative cut-off almost always gave low positive ratios only 2 of 7 or 0 of 8 such samples giving high ratios (>8) in the ELISA or RIA.
There was a broad correlation between the SBRs obtained in the two assays although, perhaps more specimens with a negative (=2) RIA ratio gave a high positive (>8) ELISA result than vice-versa [11/62 (18%) versus 1/30 (3%), p = 0.08] (Table 2).
| Table 1: | Comparison of results by cut-off and SBR methods for in-house HCMV ELISA (A) or RIA (B) |
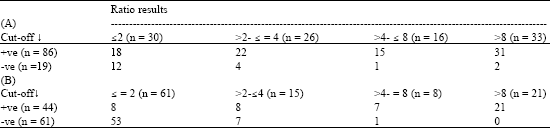 | |
| Table 2: | Comparison of SBRs calculated using in-house ELISA and in-house RIA |
 | |
In the ELISA, 74/105 sera (70%) gave an OD reading with the uninfected cell antigen greater than the mean plus three standard deviations of the ODs obtained with the negative serum. Similar information was not available for the RIA with three specimens.
Comparison of in-house and Commercial HCMV IgG ELISA
Thirty serum specimens obtained from organ donors and previously tested in a commercial ELISA (Biokit, Barcelona, Spain) were used to assess the reproducibility of the in-house ELISA in three successive assay runs.
The SBRs in the in-house ELISA were similar for 8 sera in all three assay runs (<2 for 3, >2-≤4 for 3, 8 for 2) and for a further 16 in two of the three assay runs (<2 for 2, >2-≤4 for 4, >4-≤8 for 2, >8 for 8). If specimens positive in 2 or more of three of the in-house assay runs were considered positive the copositivity and conegativity of the in-house and commercial assays were 24 of 26 (92%) and 4 of 4 (100%) (ratio method) or 20/26 (77%) ≤ and 2 of 4 (50%) (cut-off method). If the results were calculated on the basis that the positives were those reactive in both initial assay runs (i.e., repeatedly positive), the copositivity and conegativity were 22 of 26 (85%) and 4 of 4(100%) (ratio method) or 17 of 26 (65%) and 3 of 4 (75%) (cut-off method). With the six serum specimens positive in the commercial assay but giving one or more negative ratios in the in-house ELISA, the 10 positive ratios were mostly low (8≥2-≤4, 2≥4-≤8).
Evaluation of HCMV IgG ELISA and IgG avidity Assays Using Serial Sera Collected from Sperm Donors
In this section, serial serum samples from immunocompetent healthy adults most unlikely to have been transfused with blood products or immunoglobulin were selected for HCMV IgG antibody and avidity testing. The only available subjects were young male volunteer sperm donors aged 15-25 years. Serial blood samples had been collected from these individuals at irregular intervals for HBS antigen and HIV antibody tests. Approximately equal proportions [10 (29%) and 9 (26%) of the 34 sperm donors were positive or negative for HCMV IgG antibodies by ELISA in all available serum samples by cut-off and when calculated by SBR. The SBRs were generally similar in magnitude for each donor giving only positive or negative readings throughout the period of specimen collection.
Sera from seropositive donors mostly had antibody avidities >70% or >50% with 20 mM [23/33 (70%) or 32/33 (97%)], but both prevalences decreased progressively with DEA concentrations of 35 mM [16/33 (48%) or 27/33 (82%)] or 70 mM [4/33 (12%) or 14/33 (42%), P<0.001 for both trends]. The frequencies of low or very low avidities (<30% or <10%) among the same sera were zero (0%) of 33 with both 20 and 35 mM DEA but respectively 8 (24%) or 2 (6.1%) of 33 with 70 mM DEA.
The proportions of sera from seronegative sperm donors giving very high (>70%) or high (>50%) avidity indices were small [1 (3.3%) or 4 (13%) of 30 with 20 mM DEA, 4 (13%) or 6 (20%) of 30 with 35 mM DEA, 2 (6.7%) or 2 (6.7%)of 30 with 70 mM DEA] and much less than the equivalent frequencies for seropositive sperm donors (as stated above and P<0.001) except with >70% and 70 mM DEA [2/30 (6.7%) with seronegative sera, 4/33 (12%) with seropositive sera, p>0.1]. Not all sera from antibody negative subjects gave low or very low avidity indices (<30% or <10%) [prevalences 23 (77%) or 22 (74%) of 30 for 20 mM DEA, 26 (87%) or 23 (77%) of 30 for 35 mM DEA, 28 (93%) or 27 (90%) of 30 for 70 mM] though the proportions increased with higher DEA concentrations (p<0.05 for both trends). Also, every proportion for low avidities was markedly lower with seropositive rather than seronegative sperm donors (p<0.001). Thus overall the HCMV IgG antibody avidities were strikingly higher with seropositive rather than seronegative donors. Figure 1 shows on the whole comparison of HCMV IgG avidities for serial sera from sperm donors by scatter diagram.
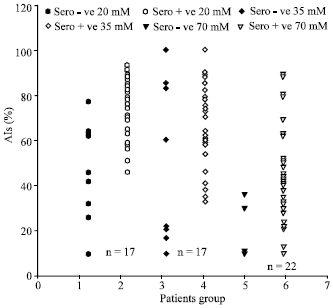 | |
| Fig. 1: | Comparison of HCMV IgG avidities for serial sera from sperm donors. Consistent negative or positive for HCMV IgG antibodies by all 3 denaturants |
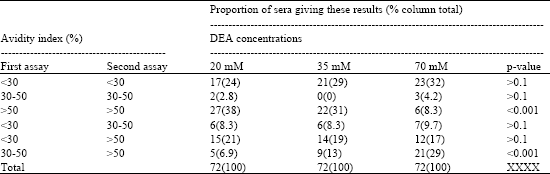
| Table 3: | Results of two HCMV IgG avidity assays on 72 serum samples |
 | |
Reproducibility of HCMV IgG Avidity Assays
Seventy two serum samples selected from those subjected to HCMV IgG avidity testing in former sections were examined for a second time using the same assay. To assess the reproducibility of the HCMV IgG avidity test, avidity indices were grouped (<30, 30-50 and >50) for 20, 35 and 70 mM DEA, the proportions of sera giving indices in the same group in both assays were similar <30% and 30-50% but decreased between the higher two denaturant concentrations for >50% and overall [46/72 (64%), 43 (60%) or 32 (44%), p<0.001]. The fractions of samples with one low avidity (<30%) and either of the other avidities did not vary with the DEA concentration but was greater when the discrepancy was large (<30% and >50%) rather than small (<30% and 30-50%) for 20 mM (p<0.02), 35 mM (p<0.05) and possibly 70 mM denaturant (p>0.05). The frequency of the avidity combination 30-50% and >50% increased between DEA concentrations of 35 and 70 mM (Table 3).
Screening and IgG Avidity Measurement of Sera from Normal Children
A total of 556 sera collected from normal children within 3 years were subjected to HCMV IgG ELISA and avidity assay. These samples were supplied by Dr J Thomas, Preston Public Health laboratory. The proportion of sera giving SBR and cut-off positive readings declined between 1<3 years and 7<9 years (p<0.01) but was relatively stable thereafter (Table 4).
No consistent change with age was seen in the proportion of sera with discrepant ELISA readings. The proportion of samples doubly negative by ELISA did not change throughout age range (p>0.1). In all age groups the fraction of sera positive either SBR or cut-off was high (73-90%).
For SBR and cut-off positive sera the distribution of avidity indices was same for 70 mM and 35 mM DEA but the values were higher for 20 rather than 35 or 70 mM (p<0.001). For sera positive by either SBR or cut-off alone, the avidity indices were similar for 70 mM and 35 mM DEA but higher for 20 rather than 35 mM (p<0.05) or 70 mM DEA (p<0.001) (Table 5).
Sera negative by both methods of result calculation had a similar distribution of avidity indices for all DEA concentrations. Sera positive only by SBR had lower avidities than those positive by cut-off and SBR or positive only by cut-off with all DEA concentrations (p<0.005). Doubly negative samples had lower avidity indices than those positive by cut-off alone at all DEA concentrations (p<0.001). Specimens with negative SBR and cut-off readings had avidity indices similar to those of specimens positive only by SBR at 70 mM and 35 mM (p>0.1) but lower values at 20 mM DEA (p<0.05). For overall results are shown in Fig. 2 and 3.
| Table 4: | HCMV IgG ELISA results for sera from normal children grouped by age |
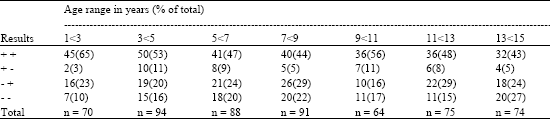 | |
| + + positive by SBR and cut-off, + - positive by SBR and negative by cut-off, - + negative by SBR and positive by cut-off, - - negative by SBR and cut-off | |
| Table 5: | Distribution of HCMV IgG avidity indices in sera from normal children |
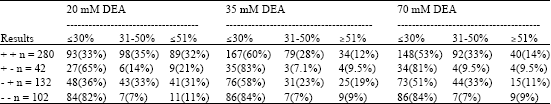 | |
| Each value is number of sera having the given ELISA ratio and cut-off results and an IgG avidity index in the specified range. Results in order ratio then cut-off | |
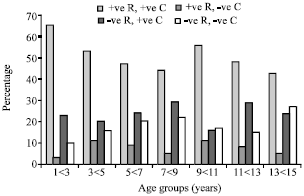 | |
| Fig. 2: | Distribution of IgG antibody amongst normal children against HCMV and HEL antigens. C refers to cut-off and R denotes to ratio |
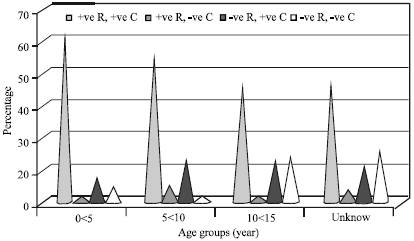 | |
| Fig. 3: | Distribution of IgG antibody amongst leukaemic children against. HCMV and HEL antigens C refers to cut-off and R denotes to ratio |
Screening and IgG Avidity Measurement of Sera from Leukaemic Children
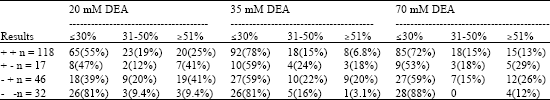
A set of sera from leukaemic children collected at the time of diagnosis of the cancer were also subjected to screening and avidity testing by HCMV ELISA (Table 6).
The percentages of sera giving ratio and cut-off positive or just ratio or cut-off positive results showed no significant change with age (p>0.1). The fraction of samples yielding doubly negative readings was small but increased between 5<10 and 10<15 years (p<0.005). Those specimens, where the age was unknown showed prevalences similar to the average of other values (p>0.1).
Distribution of Avidity Indices in Sera from Leukaemic Children
For doubly positive sera the avidity indices were the similar for 70 and 35 mM DEA but higher for 20 than 35 mM (p<0.005) or 70 mM DEA (p<0.001), (Table 7). For sera positive only by ratio the avidity indices were stable between 70 mM and 35 mM and then perhaps increased between 35 mM and 20 mM DEA (p>0.1). With samples positive only by cut-off, the avidities were stable between 70 and 35 mM DEA and then increased between 35 and 20 mM DEA (p<0.05).
| Table 6: | Age distribution of HCMV IgG ELISA results for children with leukemia |
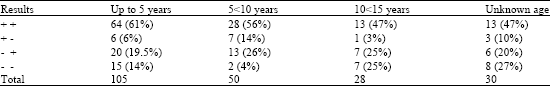 | |
| + + positive by SBR and cut-off, + - positive by SBR and negative by cut-off, - + negative by SBR and positive by cut-off, - - negative by SBR and cut-off | |
| Table 7: | Distribution of HCMV IgG avidity indices in sera from leukaemic children |
 | |
| Each value is number of sera having the given ELISA ratio and cut-off results and an IgG avidity index in the specified range. Results in order ratio then cut-off | |
For doubly negative sera the avidity values were stable throughout. Furthermore, for all DEA concentrations the avidity indices were possibly or definitely higher with solely ratio or cut-off positive than doubly positive samples (p>0.1 and p<0.05, respectively) similar for both categories of sera giving discrepant readings, but lower for doubly negative specimens than those in all other groups (p<0.001).
DISCUSSION
Antigen Production
Prior to optimization of the ELISA, the first requirement was the availability of appropriate and efficient antigen for coating the plate(s) for this reason the virus must be grown in the correct cells. Therefore, HCMV was cultivated on HEL cells which are widely regarded as the most appropriate source for the corresponding virus. However, in order to infect cells with viral inoculum, the infectivity of the virus had to be measured. Hence, technique of TCID50 was employed to find out the infectivity level of HCMV. Furthermore, the concentration of newly produced antigen was determined by protein assay since the antigen was crude. The accuracy of measuring absorbance in the protein assay at 540 and 620 nm rather than recommended 580 nm was assessed wide absorption curve suggested that protein assay suggested that protein assay was probably not greatly affected by chosen strategy.
Another measure to increase the accuracy of in-house ELISA was the employment of control antigen from uninfected cells. The latter antigen helped in producing results in form of specific binding ratio (SBR) readings (Klapper et al., 1990a).
Optimization of In-House ELISA
In order to do this, the ELISA for HCMV IgG antibodies every step of the protocol had to be optimized, the best way of standardization of an ELISA was checkerboarding. For this purpose three sets of sera were chosen for every step of optimization process; namely S +ve, W +ve and -ve. Confirmation of the reactions of these sera was obtained from diagnostic laboratory for HCMV. Therefore, checkerboarding was set up for every step including: antigen, serum, anti-IgG monoclonal antibody and enzyme conjugate dilutions. Besides, washing times both after coating the plates and after antibody binding were investigated however, the optimization of ELISA might have been further improved by using smaller dilution steps in checkerboards. The optimal protocol was chosen after careful consideration of the results of every checkerboarding and in particular where the best discrimination was observed between S +ve, W +ve and –ve sera.
Preference of Type of ELISA
Indirect ELISA has been proved efficient for the detection of antibodies to human herpesviruses (Shen et al., 1992; Yamanaka et al., 1992). The ELISA in this project was therefore indirect and avidine-biotin amplification increased its sensitivity. After adding biotinylated monoclonal antibody the plates were washed and avidine peroxidase was then added to the appropriate wells. By doing this unbound antigen and antibody were washed away before conjugate and subsequently substrate addition. The latter reduced the chances of false positivity and increased specificity and sensitivity.
IgG Avidity and Selection of Denaturant
In this project, in spite of different denaturant used by other workers, DEA was employed which proved satisfactory by other workers (Thomas and Morgan-Capner, 1991; Thomas et al., 1993). It was therefore evaluated at 20, 35 and 70 mM concentrations and not only produced better results but also was easily prepared. The avidity indices for sera from patients with recent primary or recurrent infection declined with increasing DEA concentration because of the effects of more powerful denaturation on antigen-antibody bonds of a given avidity. The greater decline in avidity with increasing DEA concentrations for sera collected after primary rather than recurrent HCMV infection presumably reflected the lower avidities seen after primary infection. It was clear that testing with more than one DEA concentration was helpful for detection of differences in antibody avidity after primary or recurrent HCMV infection and the maturation of that avidity after primary infection. For example, low avidity index (<30%) was always predictive of recent infection except when the denaturant was 70 mM DEA, but the association between recent recurrent infection and a high antibody avidity index (>50%) was strongest with the latter denaturant concentration.
In the presentation of avidity index results, they were mostly divided into 3 groups: = 30% (band 1), 31-50% (band 2) and = 51% (band 3). Samples with an avidity index of = 30% were considered to indicate low avidity, samples with indices of 31-50% were regarded as showing intermediate avidity and samples with high avidity indices were those with indices = 51% (Thomas and Morgan-Capner, 1991).
RIA Versus ELISA for HCMV IgG Antibodies
As we compared the HCMV RIA and the HCMV ELISA the latter showed to be more sensitive. The same observation was reported by Demmler et al. (1986). The added sensitivity of ELISA was more marked when results were determined using cut-off rather than the SBR method. The low ELISA SBRs for ELISA positive versus RIA negative samples confirmed that these sera contained low levels of HCMV antibodies detectable only by ELISA. As RIA has previously been shown to be more sensitive than CFT (Klapper et al., 1990b) and LAT (Morris et al., 1990c), high sensitivity of the in-house HCMV ELISA was hereby confirmed.
Comparison of In-House and Commercial HCMV IgG ELISAs
The in-house ELISA generated marginally fewer positive results than the commercial indirect ELISA. The commercial assay lacked the cell control antigen of the in-house ELISA and therefore had a possible tendency to a lack of specificity particularly as the sera positive by commercial ELISA but negative by in-house gave only very low avidity indices.
HCMV IgG ELISA and Avidity Assay Reproducibility
During the optimization of the HCMV ELISA, the inter-assay coefficient of variation for 30 sera in successive assay runs performed on plates coated with antigen overnight was 31%. This apparently high value had to be interpreted in the light of the use of only three tests. The essentially similar HCMV RIA showed a much lower coefficient of 13% when 12 tests were performed (Klapper et al., 1990b). It is therefore impossible to know whether the inter-assay CV of the HCMV IgG ELISA was truly higher than <15% variation recommended by Balfour and Harford (1990). In addition, the inter-assay reproducibility of the HCMV IgG avidity assay was assessed by comparing the results of two assays done on 72 serum samples. Most sera gave avidity indices within the same band for the lower two DEA concentrations (20 and 35 mM) and even with the most concentrated denaturant (70 mM DEA) the value was 44%. The observations were compatible with the highest DEA concentration providing more variable denaturation of antigen and antibody bonds than the two lower denaturant concentrations.
Differentiation of Primary from Recurrent Herpes Virus Infection by IgG ELISA
The importance of differentiating primary from recurrent HCMV infection lies in the widely-held view that disease is more frequent and more likely to be severe with the former than the latter category of infection (Morris et al., 1990b). The differentiation of apparently primary or recurrent HCMV infection by the absence or presence of HCMV IgG antibodies as assayed by commercial indirect ELISA in pre-infection serum samples was on first impression largely validated by the results obtained with the in-house HCMV IgG ELISA. Where discrepancies occurred in 5 (29%) of 17 patients before presumed primary infection, 4 (80%) of the 5 patients donated other pre-infection samples which gave doubly negative in-house ELISA results. Only one of the five specimens was positive by both methods of results determination. These facts perhaps suggested that discrepant results for samples collected prior to apparently primary infection reflected the known lack of absolute reproducibility of the HCMV IgG ELISA. However, the alternative hypothesis was that, at least in some instances these patients with mixed positive and negative HCMV IgG ELISA readings truly had recurrent HCMV infection following primary infection in the past. Their HCMV antibodies declined to a such low level that variable readings were obtained in the IgG ELISA.
Importance of HCMV: Prevention and Diagnosis
This matter was addressed for HCMV principally by testing serial sera from 37 sperm donors and 27 single serum samples also subjected to testing by commercial indirect ELISA for HCMV IgG antibodies. Differentiation of seropositivity from seronegativity for HCMV IgG antibodies plays a major role in distinguishing subsequent primary from recurrent HCMV infection. Accurate identification of HCMV seropositivity is also crucial in identifying blood, organ, or sperm donors who might transmit HCMV infection (Klapper et al., 1990b; Morris et al., 1990a-c). For susceptible immunocompromised patients, exclusion of seropositive persons as blood donors may be appropriate (Klapper et al., 1990b). Matching of organ transplant donors and recipients for HCMV IgG antibodies status has been proposed as a method of reducing the incidence of severe HCMV disease associated with primary HCMV infection in a seronegative recipient of an organ from seropositive donor (Wreghitt et al., 1988). HCMV IgG antibody matching of semen donor and recipient has been proposed as a means of avoiding primary HCMV infection in pregnancy with its attendant risk of the birth of a baby with HCMV-induced congenital defects.
HCMV and Sera of Normal and Leukaemic Children
Approximately two-thirds of sera from normal and leukaemic children gave uniformly positive or negative ratio and cut-off readings in the HCMV IgG ELISA [382/556 (69%) or 150/213 (70%), Table 4 and 6]. The similarity in these percentages suggested that this was a property of the test which was perhaps independent of the patient population under test. The distribution of avidity indices was similar with normal children when the cut-off reading was positive and the SBR either positive or negative, presumably indicating that specimens with both categories of results contained HCMV IgG antibodies. At least some of the ratio positive cut-off negative ELISA readings probably also presented HCMV seropositivity, in that the avidity indices for these sera from normal children were intermediate between those of samples giving uniformly positive or negative readings. One possibility was that the latter specimens came both from truly seropositive and truly seronegative individuals. Alternatively the HCMV antibody levels and avidities in these persons had fallen to low levels with time following primary infection. With sera from children with leukemia, the avidities were lower with ratio and cut-off positive samples than singly positive samples, again supporting the view that the latter were probably obtained from seropositive individuals. The role of avidity assays in identifying HCMV positive subjects was confirmed here, in that both studies of children revealed mostly low avidity indices (indicative of easy dissociation of serum components from negative) when the ELISA ratio and cut-off readings were both negative.
CONCLUSION
The HCMV IgG antibody data was remarkable in the light of the accepted view of the seroepidemiology of this virus infection in Caucasian populations. The prevalences of ELISA ratio and cut-off positive readings or positive readings by any method of results calculation were stable in normal children throughout all age groups. If only ELISA ratio and cut-off reactive sera were considered truly positive, the antibody prevalence was approximately 50%, much higher than the 20-30% reported in studies using a commercial HCMV IgG ELISA in white US children (White et al., 1989). The higher prevalence of HCMV IgG antibodies in this ELISA rather than previous studies among children most probably reflected the higher sensitivity of in-house ELISA.
REFERENCES
- Nielsen, S.L., I. Sorensen and H.K. Andersen, 1988. Kinetics of specific immunoglobulins M, E, A and G in congenital primary and secondary cytomegalovirus infection studied by antibody-capture enzyme in immunosorbent assay. J. Clin. Microbiol., 26: 654-661.
PubMed - Thomas, H.I.J. and P. Morgan-Capner, 1991. Rubella-specific IgG1 avidity: A comparison of methods. J. Virol. Methods, 31: 219-228.
PubMedDirect Link - Thomas, H.I.J., P. Morgan-Capner and N.S. Connor, 1993. Adaptation of a commercial rubella-specific IgG kit to assess specific IgG avidity. Serodiagn. Immuno. Ther. Infect. Dis., 1: 13-16.
CrossRef - White, N.H., M.D. Yow, G.J. Demmler, H.J. Norton and J. Hoyle et al., 1989. Prevalence of cytomegalovirus antibody in subjects between the ages of 6 and 22 years. J. Infect. Dis., 159: 1013-1017.
Direct Link - Yamanaka, T., K. Kiuoyani, T. Sakaguchi, Y. Fukuda and K. Dohi et al., 1992. Detection of cytomegalovirus in urine samples by an enzyme linked immunosorbent assay using a monoclonal antibody against the viral 150 kilo Dalton protein. J. Clin. Microbiol., 30: 685-690.
Direct Link