
R. Thavasi
CAS in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
S. Jayalakshmi
CAS in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
T. Balasubramanian
CAS in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
Ibrahim M. Banat
School of Biomedical Sciences, University of Ulster,
Coleraine BT52 1SA, Northern Ireland, UK
Research Journal of Microbiology
Year: 2006 | Volume: 1 | Issue: 5 | Page No.: 401-408







ABSTRACT
The present study deals with the hydrocarbon degrading potential of a marine nitrogen fixing bacterium Azotobacter chroococcum isolated from Tuticorin harbor (Lat. 08o45`N; Long. 78o13`E). Degradation of crude oil (58%) and emulsification (D610) of waste motor oil (1.51), crude oil (1.43), pea nut oil (1.39), diesel (0.69), kerosene (0.81), Naphthalane (0.36), Anthracene (0.33) and xylene (0.42) indicated its potentiality in utilization of various hydrocarbons. Growth of Azotobacter chroococcum in a mineral medium with 0.5% crude oil as sole carbon source resulted maximum cell density at 96 h with an OD value of 0.333. At the end of 96 h, the cell count was 4.5x109 CFU mL-1 and 2.5 mg mL-1 of biomass. The biosurfactant production was found to be 1 mg mL-1 at 96 h. The total nitrogen fixed was 4.2 mg L-1. Two plasmids were found with molecular weight of 4788 and 2400 base pairs, respectively. Loss of biodegradation and biosurfactant production after plasmid curing was observed, which confirmed that, the biosurfactant production and biodegradation process were plasmid mediated. Results of the present study revealed the possibility of using marine nitrogen fixing hydrocarbon degrading bacteria and their biosurfactants in the abatement of marine oil pollution.
PDF Abstract XML References
How to cite this article
R. Thavasi, S. Jayalakshmi, T. Balasubramanian and Ibrahim M. Banat, 2006. Biodegradation of Crude Oil by Nitrogen Fixing Marine Bacteria Azotobacter chroococcum
. Research Journal of Microbiology, 1: 401-408.
URL: https://scialert.net/abstract/?doi=jm.2006.401.408
URL: https://scialert.net/abstract/?doi=jm.2006.401.408
INTRODUCTION
Oil exploration activities have consequential environmental impact on air, water and land. About 40% of the world’s energy supply comes from crude oil (Brown and Skipsey, 1986). With the advent of the industrial revolution and the ever increasing economic growth thereafter, there has been a concomitant increase of problems of accidental oil spills throughout the world. In addition to oil tanker accidents, another important source of oil in the sea in ballast water. It has been estimated that global volumes of ballast water transported are probably in the order of 3 to 4 billion tons per year (Ballast Water News, 2003). Conventional methods employed in the removal of spilled oil have deleterious side effects to the environment and to the biota. Hence, the recent focus is on bioremediation (Atlas and Uterman, 1999). Bioremediation is defined as the process in which microorganisms are stimulated to rapidly degrade hazardous organic contaminants to environmentally safe level. The microbes involved in a particular bioremediation process may obtain both energy and carbon source through the metabolism of target organic contaminants (Bollag et al., 2000). The degradation of hydrocarbons in the marine environment is influenced by the availability of nutrients such as nitrogen and phosphorus (Leahy and Colwell, 1990; Atlas, 1991). Mulkins-Phillips and Stewart (1974) reported that 0.05 mg of elemental nitrogen was required to remove 1 mg of hexadecane by a monoculture. To overcome the problem of nitrogen limitation, the present study was undertaken with a nitrogen fixing bacteria.
MATERIALS AND METHODS
Isolation and Identification of Nitrogen Fixing Oil Degrading Bacteria
Sediment sample was collected from Tuticorin new harbor (Lat. 08o45’N; Long. 78o13’E) at a depth of 5 m using a Petersen grab. The central portion of the sediment was used for the bacterial isolation. One gram of the sediment was transferred to a conical flask containing 50 mL of nitrogen free mineral medium (K2HPO4, 1.0 g; MgSO4.7H2O, 0.2 g; FeSO4.7H2O, 0.05 g; CaCl2.2H2O, 0.1 g; Na2MoO4.2H2O, 0.001 g; Glucose, 10 g and distilled waster 1000 mL, pH 8.0). The flasks were incubated in a rotary shaker at 150 rpm and 30°C for 120 h. One mililiter of serially diluted enriched culture was plated on nitrogen free agar medium containing 0.5% of crude oil. Plates were incubated at 30°C for 24-72 h. Strain was identified up to the species level by following Bergy’s manual of determinative bacteriology (Buchanan et al., 1974).
Isolation and Biochemical Analysis of Biosurfactants
Azotobacter chroococcum was cultured in conical flasks containing 50 mL of nitrogen free medium containing 0.5% of crude oil as a sole carbon source. Flasks were inoculated with 1 mL of inoculum with a cell density of 3.00x104 CFU mL-1 and incubated in a shaker at 150 rpm and 30°C for 120 h. Sampling was made at an interval of 24 h for the estimation of growth of the bacteria. Growth studies were made by measuring optical density of the culture at 610 nm and cell count (CFU mL-1) was estimated by direct plate counting method. When the culture attained its early stationary phase (i.e.) after 96 h of incubation, the broth was centrifuged at 6000 rpm for 20 min and three volumes of ice cold acetone were added to the cell free broth and allowed to stand overnight. The precipitated biosurfactant was collected by centrifugation at 12000 rpm for 20 min and the precipitate was dialyzed against distilled water at 4°C for 12 h and lyophilized (Juwarker and Krishager, 1991). Further purification was done in a silica gel column and the elution were made with chloroform and methanol ranging from 20:1 to 2:1 vol/vol in a gradient manner. The fractions were pooled and the solvents were evaporated and the resulting residue was dialyzed against distilled water and lyophilized. The biochemical composition of biosurfactant was analyzed using standard methods. Carbohydrate content of the biosurfactant was determined by the phenol-sulfuric acid procedure (Dubois et al., 1956). Protein content was determined by the method of Lowry et al. (1951) and lipid content was estimated by adopting the procedure of Folch et al. (1956).
Estimation of Emulsification Activity
Purified biosurfactant (1 mg mL-1) was dissolved in 5 mL of Tris buffer (pH 8.0) in a 30 mL screw capped test tube. Waste motor oil, crude oil, peanut oil, diesel, kerosene, naphthalene, anthracene and xylene were tested for emulsification activity. Five miligram of hydrocarbon was added to the above solution and shaken well for 20 min in a shaker at 150 rpm and the mixture was allowed to stand for 20 min. The Optical Density (OD) of the mixture was measured at 610 nm and the results were expressed as D610 (Rosenberg et al., 1979). Among the hydrocarbons used for emulsification assay, the strains showed maximum emulsification activity against crude oil. Hence crude oil was selected for the biodegradation study.
Estimation of Crude Oil Degradation and Nitrogen Fixation
Cell free culture broth was extracted with three volume of toluene and the extract was made up to 10 mL and the OD was measured at 420 nm. The percentage of degradation was calculated from the standard curve. The standard curve was obtained with different concentration of crude oil in toluene (Rahman et al., 2002b). The experiment was carried out in duplicate and the mean value was expressed as % of crude oil degraded. The crude oil used in this study was obtained from Madras Refineries Ltd., Manali, Chennai, Tamil Nadu, India. It’s specific gravity was reported as 0.8445 at 25°C and 0.8529 at 15°C. Nitrogen fixation was estimated from the cell free culture broth by adopting the micro kjeldhal method (Clesceri et al., 1998).
Bacterial Adhesion to Hydrocarbons (BATH)
The adherence ability of the A. chroococcum with oil was measured using the BATH test as described by Rosenberg et al. (1980). Ten milliliter of culture containing 3.00x104 CFU mL-1 was added to 0.7 mL of crude oil in a 20 mL glass tube and mixed well. The optical density of the cells in the culture broth was determined before the addition of crude oil. After 30 min of static storage, the aqueous phase was carefully removed by a Pasteur pipette and the optical density was measured again. The percentage of bacteria adhered to the oil was calculated using the following formula:
Percentage of bacterial adherence = (1-(OD shaken with oil/OD original))x100 = % |
The method described by Betts et al. (1989) was used to observe the viable bacteria on the oil droplets. Few drops of 2-(4-iodophenyl)-3-(4-nitrophenyl)-5-phenyltetrazolium chloride hydrate (INT) solution was added to the culture broth prior to observation using a light microscope. The INT turned red when reduced inside the cells, indicating the viability and the adherence of the cells with the oil droplets. Biodegradation, emulsification and BATH experiments were done in duplicates and the results expressed here are the mean values.
Isolation of Plasmid and Plasmid Curing
To check whether the biodegradation and biosurfactant is plasmid mediated, the Azotobacter chroococcum strain was screened for the presence of plasmids. Isolation of plasmid was done adopting the method of Sambrook and Russel (2001). The molecular weight of the isolated plasmids was determined by using software (Total Lab software). ECOR1 Hind-III double digest was used as a molecular weight marker.
Plasmid curing was done using acredine orange at a concentration of 500 μg mL-1 which was added to 18 h old broth culture and incubated for 12 h (Fujji et al., 1997). Crude oil degradation and biosurfactant production experiments were repeated for the plasmid cured strains.
RESULTS AND DISCUSSION
The isolate was identified as Azotobacter chroococcum (Table 1). The existence of Azotobacter chroococcum from the oil polluted site was witnessed by Onwurah (1999) and their existence in mangrove environment by Ravikumar et al. (2004). This microorganism is of special interest in contributing nitrogen economy of both terrestrial and marine ecosystems (Dicker and Smith, 1980). Hydrocarbon contamination in a particular area may increase the fraction of hydrocarbon-utilizing microorganisms and it may also increase the capacity of the microbial community to degrade hydrocarbons (Leahy et al., 1990). The present research also confirmed that, Azotobacter chroococcum could contribute substantially to the nitrogen requirement of the oil-degrading bacteria in the marine environment. From a cell density of 3.00x104 CFU mL-1, the strain attained its maximum (4.5x109 CFU mL-1) in 96 h with an optical density of 0.333 (Fig. 1). It had a dry weight of 2.5 mg mL-1 and the biosurfactant concentration obtained was 1 mg mL-1. The ability of the Azotobacter chroococcum to grow and utilize the crude oil in nitrogen free mineral medium indicated that it does not require additional nitrogen to degrade crude oil like other oil degrading bacteria. In the present study 58% of the crude oil was degraded by the organism at the end of 120 h of incubation. Onwurah (1999) reported 62-65% of crude oil degradation by an Azotobacter sp. in 20 days. Tuticorin harbor (Lat. 08o45’N; Long. 78o13’E) is a busy harbor and where the Indian oil Corporation is located and receiving crude oil from ships for distillation process, which might have an impact on the surrounding coastal waters.
| Table 1: | Physiological and biochemical characteristics of A. chroococcum |
 | |
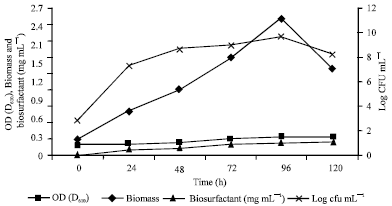 | |
| Fig. 1: | Growth and biosurfactant production of Azotobacter chroococcum |
Emulsification of various hydrocarbons by the biosurfactant (Table 2) and the ability of cell adherence (Fig. 2) with crude oil (21.4%) confirmed the degradation potential of A. chroococcum. Emulsification of crude oil, xylene, pristine, n-octane, n-decane, n-hexadecane and n-dodecane by the emulsification factor produced by a marine bacteria was reported by Juwarkar and Khirsagar (1991). Emulsification of crude oil by an alkane-oxidizing Rhodococcus species isolated from seawater was reported by Bredholt et al. (1998). Bioemulsifiers play an important role in the regulation of the attachment and detachment of microorganisms to and from hydrophobic surfaces (Ron and Rosenberg, 2001). Biosurfactant production accompanying the growth of oil degrading bacteria may help in the adherence of the cells to the substrate molecules and in metabolizing them (Bharathi and Vasudevan, 2001).
Biosurfactants are produced by a variety of microbes, either extracellular or attached to parts of cells (Sydatk and Wagner, 1987). Biosurfactants are often produced when grown on n-alkanes or other water-immiscible substrates (Bredholt et al., 1998). Degradation of oil in the marine environment by autochthonous forms depends on the capacity of these bacteria to solubilize and emulsify the oil (Juwarker and Khirsagar, 1991). Biosurfactants are generally considered to represent a mechanism for the solubilization or emulsification of water-insoluble substrates to facilitate transport by the cells (Singer and Finnerty, 1990). The emulsification property of the biosurfactant indicated that it has solubilized the hydrocarbons to a greater extent. Biochemical analysis of the biosurfactant revealed that it is a glycolipid with 31.3% of carbohydrate and 68.7% of lipid. A similar kind of glycolipid was isolated from Pseudomonas aeruginosa (Zhang and Miller, 1992) and Pseudomonas putida (Tuleva et al., 2001).
| Table 2: | Emulsification of various hydrocarbons by the biosurfactant isolated from A. chroococcum |
 | |
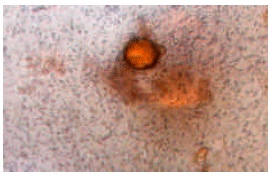 | |
| Fig. 2: | Bacterial cells adhered to the crude oil |
The present observation is highly encouraging and this type of biopolymer has a greater advantage in oil industries and also in marine oil spill problems. Maximum biosurfactant concentration was found at the early stationary phase. This may be due to of the production of biosurfactant as a secondary metabolite (Rahman et al., 2002a). Rosenberg et al. (1979b) also found the accumulation of biosurfactant during the stationary phase in Arthrobacter RAG-I strain using hexadecane and ethanol as carbon sources. In the present study nitrogen fixation was found to be constant through out the period of study (4.2 mg L-1). Similar kind of observation was reported by Deka (1998), in A. chroococcum strain isolated from an oil polluted soil. Degradation of crude oil was found to increase gradually with increasing cell number and incubation period, whereas in the case of nitrogen fixation it was found to be constant throughout the period of incubation. This might be due to the utilization of nitrogen during the growth and degradation process. Thus nitrogen fixing capacity is an added advantage, as the strain may be useful in cleaning the oil that spilled in nitrogen limited environments.
The present study confirmed the presence of plasmids and their involvement in the degradation of crude oil. Two plasmids were found with molecular weight of 4788 and 2400 base pairs (Fig. 3). The presence of multiple plasmids in hydrocarbon degrading bacteria was already reported in P. putida, P. diminuta, Bacillus coagulans, Brevibacterium linesepidermidis (David et al., 1995) and Rhodococcus sp. (Singer and Finnerty, 1990). After plasmid curing, complete loss of degradation and emulsifier production properties were observed. This confirmed the role of plasmids in these activities. The results obtained by Chakrabarty (1976) and David et al. (1995) supporting the findings of the present study. Oxygenase enzymes that are involved in biodegradation of crude oil are encoded by plasmids and others on chromosomal genes (Rosenberg et al., 1996). Heterotrophic bacteria were observed to exhibit a higher incidence of plasmid DNA in hydrocarbon-contaminated environments, such as offshore fields (Hada and Sizemore, 1981).
The results of the present study indicated that the strain A. chroococcum is an efficient strain with very high potential of crude oil degradation, through biosurfactant production and nitrogen fixation. Either live cells (or) biosurfactant alone may be applied to the oil polluted sites. The present study clearly indicated the possibility of developing an environmentally friendly mitigation technology against oil pollution using the indigenous bacterial flora of the ecosystem.
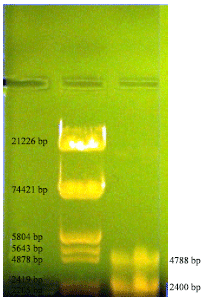 | |
| Fig. 3: | Plasmids of Azotobacter chroococcum, lane 1. molecular marker Hind III double digest |
Future scope of the work is to develop a strain by transforming the plasmids from the A. chroococcum to naturally occurring fast growing marine bacteria and to other nitrogen fixing forms to overcome the problem of oil spills.
REFERENCES
- DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Fujji, T., M. Takeo and Y. Maeda, 1997. Plasmid-encoded genes specifying aniline oxidation from Acinetobacter sp. Strain YAA. Microbiol., 143: 93-99.
Direct Link - Leahy, J.G. and R.R. Colwell, 1990. Microbial degradation of hydrocarbons in the environment. Microbiol. Mol. Biol. Rev., 54: 305-315.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Rahman, K.S.M., I.M. Banat, J. Thahira, T. Thayumanavan and P. Lakshmanaperumalsamy, 2002. Bioremediation of gasoline contaminated soil by a bacterial consortium amended with poultry litter, coir pith and rhamnolipid biosurfactant. Bioresour. Technol., 81: 25-32.
CrossRefDirect Link - Rahman, K.S.M., T.J. Rahman, S. McClean, R. Marchant and I.M. Banat, 2002. Rhamnolipid biosurfactant production by strains of Pseudomonas aeruginosa using low-cost raw materials. Biotechnol. Prog., 18: 1277-1281.
CrossRef - Ron, E.Z. and E. Rosenberg, 2001. Natural roles of biosurfactants. Environ. Microbiol., 3: 229-236.
CrossRefPubMedDirect Link - Rosenberg, M., D. Gutnick and E. Rosenberg, 1980. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett., 9: 29-33.
CrossRefDirect Link - Zhang, Y. and R.M. Miller, 1992. Enhanced octadecane dispersion and biodegradation by a Pseudomonas rhamnolipid surfactant (biosurfactant). Applied Environ. Microbiol., 58: 3276-3282.
Direct Link