
Marta Martins
Unit of Mycobacteriology and UPMM (Centre for Medical Parasitology and Microbiology), Institute of Hygiene and Tropical Medicine, Universidade Nova de Lisboa, Rua da Juuqueira 96, 1349-008 Lisbon, Portugal
Miguel Viveiros
Unit of Mycobacteriology and UPMM(Centre for Medical Parasitology and Microbiology), Institute of Hygiene and Tropical Medicine, Universidade Nova de Lisboa, Rua da Juuqueira 96, 1349-008 Lisbon, Portugal
Diane Ordway
Colorado State University, Department of Microbiology, Immunology and
Pathology, Mycobacteria Research Laboratory, 1682 Campus Delivery,
200 West St Fort Collins, CO 80523-1682
Jette E. Kristiansen
South Danish University, Sonderborg Syghus, 64 Sonderborg, Denmark
Joseph Molnar
Department of Microbiology and Immunology, Institute of Medical Microbiology, University of Szeged, H-6720, Szeged, Dom ter 1 0. Hungary
Leonard Amaral
Unit of Mycobacteriology and UPMM Centre for Medical Parasitology and Microbiology), Institute of Hygiene and Tropical Medicine, Universidade Nova de Lisboa, Rua da Juuqueira 96, 1349-008 Lisbon, Portugal
Research Journal of Microbiology
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 203-209







ABSTRACT
Killing of bacteria by neutrophils is dependent upon the availability of potassium. Although macrophages derived from human peripheral blood monocytes have little killing activity of their own, they can be transformed into effective killers of Staphylococcus aureus and Mycobacterium tuberculosis by in vitro exposure of the macrophage to clinically relevant concentrations of phenohiazines, namely, thioridazine or chlorpromazine. Because transport mechanisms dependent upon the availability of calcium are inhibited by these agents, the possibility that other agents which have similar activity also have the ability to enhance the killing of bacteria by the macrophage derived from peripheral blood monocytes was investigated. In this study we show that the presence of increasing concentrations of ouabain, reserpine or verapamil in the medium enhances the killing of Staphylococcus aureus. Because these concentrations have no activity on the replication or killing of the bacterium, killing is deemed to be due to the macrophage itself. A model is presented which describes the mechanism by which these agents and phenothiazines indirectly activate lysosomal enzymes as a result of the inhibition of potassium efflux pumps that would normally pump the ion from the phagocytic vacuole to the cytoplasm of the macrophage.
PDF Abstract XML References
How to cite this article
Marta Martins, Miguel Viveiros, Diane Ordway, Jette E. Kristiansen, Joseph Molnar and Leonard Amaral, 2006. Reserpine, Ouabain and the Calcium Channel Blocker Verapamil, Cause Intracellular Killing of Staphylococcus aureus. Research Journal of Microbiology, 1: 203-209.
URL: https://scialert.net/abstract/?doi=jm.2006.203.209
URL: https://scialert.net/abstract/?doi=jm.2006.203.209
INTRODUCTION
Recent findings suggest that potassium flux into the phagocytic vacuole is needed for demonstrable killing of phagocytosed bacteria by neutrophils (Ahluwalia et al., 2004). This potassium flux is dependent upon the activation of the potassium channel by calcium as well as by membrane depolarisation (Reeves et al., 2002). Unlike the neutrophil, human macrophages derived from peripheral blood monocytes have little killing action of their own (Ordway et al., 2002). Nevertheless, they can become effective killers of staphylococci (Ordway et al., 2002) and mycobacteria (Ordway et al., 2003) when low concentrations of phenothiazines are added to the medium (Amaral et al., 2004). This intracellular killing activity is attributed to the known ability of the lysosome of some tissues to concentrate compounds (Daniel and Wojcikowski, 1999) to levels compatible with those that kill these bacteria in vitro (Ordway et al., 2002; Ordway et al., 2003). However, phenothiazines deny calcium to flux activities that are dependent upon energy provided by calcium dependent ATPase (Savitha et al., 1990) and many of these activities which are present at the level of the plasma membrane (Sun-Wada et al., 2004) are also subject to similar calcium denial by verapamil (Lu et al., 1994), an inhibitor of efflux pumps that results from the over-expressed Pgp transporter (Loscher and Potschka, 2002). Verapamil also inhibits plasma membrane mediated transport of potassium into the macrophage by preventing access to calcium (Galli et al., 1988). Verapamil sensitive calcium dependent influx pumps present in the membranes of macrophages could be expected to be retained by that component of the plasma membrane section that becomes the phagosome and eventually fuses with a specific lysosome. However, the orientation of these pumps would now favour the extrusion of potassium from the phagocytic vacuole to the cytoplasm of the macrophage. If this is the case, then the activation of potassium dependent killing noted by others for the neutrophil (Ahluwalia et al., 2004; Reeves et al., 2002) may not be readily possible unless the activity of this potassium pump could be inhibited. The possibility that such inhibition by phenothiazines is the cause for the enhanced killing of intracellular bacteria could be perhaps mimicked by agents that have similar activity against calcium-dependent potassium pumps such as verapamil and by ouabain and reserpine, agents known to have direct (Ohmori et al., 1991) or indirect (Kanoh, 1999) effects on the potassium pumps of mammalian cells, respectively. In this respect, we have studied this possibility by first loading the monocyte derived macrophage with Staphylococcus aureus (Ordway et al., 2002), removing any non-phagocytosed bacteria with repeated washings and then added varying concentrations of ouabain, verapamil, reserpine, chlorpromazine and thioridazine, all of which are below those that have an inhibitory effect on the in vitro growth of the bacterium and produce no toxicity to the macrophage.
MATERIALS AND METHODS
The study to be described was conducted at the Mycobacteriology Laboratory of the Institute of Hygiene and Tropical Medicine, Lisbon, Portugal.
Reagents and Media Used
Reserpine, ouabain, verapamil, chlorpromazine, thioridazine, trypan blue, RPMI medium, Hank’s Balanced Salt Solution and Sodium dodecyl sulphate (SDS) were purchased from Sigma Aldrich Química SA(Madrid, Spain). Trypticase Soy Broth (TSB) and Trypticase Soy Agar (TSA) were purchased from Difco (Detroit, Mi). All solutions of reagents were prepared in distilled, sterile water on the day of the experiment.
Bacterial Strain and Preparation of Monocyte Derived Macrophages from Human Blood
Bacterial strain employed in this study was Staphylococcus aureus ATCC25923. For each experiment individual colonies were obtained from TSA plates, transferred to 10 mL of saline, mixed and an aliquot of 100 μL transferred to 10 mL of TSB and incubated at 37°C until they reached their stationary phase. The isolation and preparation of monocyte derived macrophages (MDM) from 20 mL peripheral blood donated by the authors have been previously described (Ordway et al., 2002; 2003).
Determination of Toxic Effects of Reserpine, Ouabain, Verapamil, Chlorpromazine and Thioridazine on Human Monocyte Derived Macrophages
The toxicity effect of the compounds was determined by the use of the Trypan Blue exclusion method as previously described (Ordway et al., 2002; 2003).
Determination of Minimum Inhibitory Concentrations (MICs) and Minimum Bactericidal Concentrations (MBC)
The individual MICs of reserpine, ouabain and verapamil were performed in TSB by the broth dilution method as previously described (Ordway et al., 2002; 2003). Those for chlorpromazine and thioridazine had been repeatedly determined in past studies and were not repeated here. The MIC and MBC determinations were repeated three times and the values obtained did not differ.
Determination of the Effects of Reserpine, Ouabain, Verapamil, Chlorpromazine and Thioridazine on the Killing of Intracellular Staphylococcus Aureus
The preparation of isolated macrophages at a ratio of 10 bacteria to 1 macrophage, the incubation of the mixture in RPMI medium, the period of incubation, the removal of free bacteria, the separate addition of each reagent to the culture, extraction of bacteria and performance of colony forming units have been previously described (Ordway et al., 2002; 2003). Details, however, are presented in the legends of the text.
RESULTS
Toxicity of Reserpine, Ouabain, Verapamil, Chlorpromazine and Thioridazine on Monocyte Derived Macrophages
The toxicity of reserpine, ouabain and verapamil on human monocyte derived macrophages (MDM) was determined. That of chlorpromazine and thioridazine was previously reported (Ordway et al., 2002; 2003). Briefly, concentrations as high as 100 mg L-1 of reserpine, ouabain or verapamil did not produce any toxicity against MDM (data not shown). These compounds at the highest concentrations employed for the determination of toxicity have no effect on the growth rate or final growth of Staphylococcus aureus (data not shown).
Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
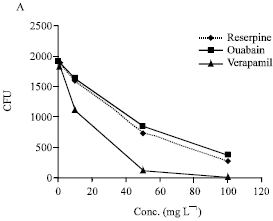
The MIC and MBC for CPZ, TZ, ouabain, reserpine and verapamil are summarised by in Fig. 1. Ouabain, reserpine and verapamil had no effect on the growth of Staphylococcus aureus at the maximum concentrations employed (i.e., 100 mg L-1).
Effects of the Compounds, Reserpine, Ouabain, Verapamil, Chlorpromazine and Thioridazine on the Killing of Intracellular Staphylococcus Aureus
The effect of various concentrations of reserpine, ouabain and verapamil on Staphylococcus aureus that have been phagocytosed b y monocyte derived human macrophages is summarised by Fig. 1A.
 | |
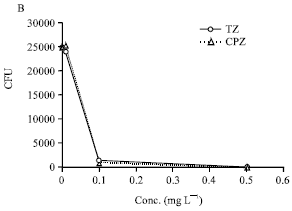 | |
| Fig. 1: | The effect of reserpine, ouabain and verapamil (A) and chlorpromazine and thioridazine (B) on Staphylococcus aureus that have been phagocytosed by human monocyte derived macrophages Legend. Macrophages derived from Ficoll isolated peripheral blood monocytes (1x106 cells mL-1) were suspended in 1 mL RPMI medium and distributed to the wells of a 24 microwell plate. After 6 days of incubation the wells were washed with RPMI medium to remove non-adherent cells (lymphocytes). The adherent monocyte derived macrophages (MDM) were counted and the yield per well was approximately 105 with less than 5% variation of each well from this number. 106 bacteria were added to each well for 30 min and after this period the wells were washed with RPMI medium to remove any non-phagocytosed bacteria and fresh medium (1 mL) added to each well. One hundred microliter of each compound in medium was added to yield the concentration that ranged from 0.0 to the maxima identified in the Figures. The microplates were incubated for a period of 0 to 6 h and the MDM lysed with SDS thereby releasing the bacteria which were then counted by the colony forming unit (CFU) method (Ordway et al., 2002, 2003) . The data presented in 1A was obtained from a number of repeat experiments involving different donations of derived MDM and due to the biological variation resulting from differences of donors, the data was normalised and the curves presented in 1A generated by regression analysis. The data presented by 1B is actual data obtained from a representative experiment |
The addition of each of these agents to the medium after the phagocytosis of the bacterium produced significant enhancement in the killing of Staphylococcus aureus in a concentration dependent manner with a maximum effect on the killing of intracellular staphylococci at the highest concentration employed.
The effects were similar to those obtained with chlorpromazine and thioridazine although these latter agents produced an enhancement of killing at concentrations in the medium that were far lower (Fig. 1B).
DISCUSSION
The results obtained in our current study show, for the first time, that inhibitors of potassium transport have the capacity to enhance the killing of bacteria that have been phagocytosed by derived macrophages that have little killing action of their own. Our previous studies (Ordway et al., 2003, 2002) suggested that the killing activity of phenothiazines was due to the ability of the macrophage to concentrate the agent to levels comparable to those in vitro that had bactericidal activity. The results of the current study suggest that agents that have similar activity as phenothiazines with respect to effects on calcium binding also enhance the killing of intracellular bacteria. Because the agents ouabain, verapamil and reserpine, unlike the phenothiazines (Amaral et al., 2004), are shown in the current study to be devoid of any in vitro activity against S. aureus, we attribute their intracellular activity to their effects on the transport of potassium in as much as these agents inhibit this transport (Kanoh, 1999; Galli, 1988).
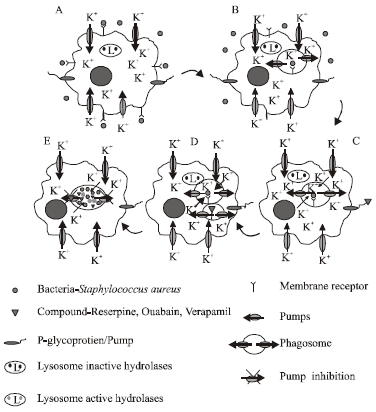 | |
| Fig. 2: | Model suggested for the killing of intracellular Staphylococcus aureus by ouabain, reserpine and verapamil, (A) Internalization of the bacteria by the macrophage, (B) Investigation of the macrophage membrane containing the bacteria, © Phagosome trapping with the bacteria, (D) Fusion of the phagosome with the lysosome and (E) Inhibition of the pumps by the compounds reserpine, ouabain and verapamil |
The results obtained in our current study coupled to those results recently obtained by others (Ahluwalia et al., 2004; Reeves et al., 2002), suggest the mechanisms presented by Fig. 2 as those involved for the killing of intracellular bacteria. Firstly, the transport of potassium into the macrophage is dependent upon calcium dependent type pumps present in the plasma membrane of the macrophage. This process of phagocytosis involves the invagination of the plasma membrane to which the bacteria is first bound (Fig. 2). The relative position of the potassium pump is now reversed in the phagosome and the transport of potassium would effectively be from the inside of the phagocytic vacuole to the cytoplasm of the macrophage (Fig. 2B). Normally, the activity of these pumps would be expected to reduce the potassium concentration of the phagosome and hence the latent hydrolases in the lysosome that now fuses with the phagosome remain inactive. These pumps would be subjected to being inhibited by agents that inhibit calcium dependent ATPase activity and hence the efflux of K+ from the phagosome-lysosome complex would be inhibited. The availability of this ion for the activation of hydrolases present in the lysosome would be insured-hence killing would take place. With respect to the activity of phenothiazines on intracellular bacteria, we should also consider that when the bacteria are trapped in the phagosome-lysosome, there is also leakage of K+ from the bacteria that is promoted by the agent (Kristiansen et al., 1982). The killing activity of macrophages that is enhanced by inhibitors of the potassium pump that is present in the membrane of the phagosome is a subject that is now under investigation and involves the isolation of the phagocytic vacuole and the evaluation of potassium efflux activity.
Acknowledgements
We wish to thank the Institute of Hygiene and Tropical Medicine, Portugal and its Scientific Council for the support given to this project. We are especially grateful to the Management Committee Members of the Cost Action B16 of the European Commission for their many helpful suggestions and to Dr. Jean-Marie Pagès for his inquisitive comments. This work was supported by EU-FSE/FEDER-POCTI-37579/FCB/2001 provided by the Fundação para a Ciência e a Tecnologia (FCT) of Portugal. M. Martins was supported by grant SFRH/BD/14319/2003 (FCT, Portugal).
REFERENCES
- Amaral, L., M. Viveiros and J. Molnar, 2004. Antimicrobial activity of phenothiazines. In Vivo, 18: 725-732.
PubMed - Savitha, G., S. Panchanathan and B.P. Salimath, 1990. Capsaicin inhibits calmodulin-mediated oxidative burst in rat macrophages. Cell. Signal., 2: 577-585.
CrossRefPubMedDirect Link