
Constantine Chobet Ondhoro
Department of Zoology, Entomology and Fisheries Sciences, Makerere University, P.O. Box 7062, Kampala, Uganda
LiveDNA: 256.30498
Gregory E. Maes
Aquaculture, Department of Biosystems, KU Leuven, 3001, Leuven, Belgium
Nalukenge Winnie Nkalubo
National Fisheries Resources Research Institute, P.O. Box 343, Jinja, Uganda
John K. Walakira
Aquaculture Research and Development Centre Kajjansi, P.O. Box 530, Kampala, Uganda
Matthew Tenywa Mwanja
Aquaculture Research and Development Centre Kajjansi, P.O. Box 530, Kampala, Uganda
Egulance Ganda
Aquaculture Research and Development Centre Kajjansi, P.O. Box 530, Kampala, Uganda
Godfrey Kawooya Kubiriza
Department of Zoology, Entomology and Fisheries Sciences, Makerere University, P.O. Box 7062, Kampala, Uganda
Jackson Efitre
Department of Zoology, Entomology and Fisheries Sciences, Makerere University, P.O. Box 7062, Kampala, Uganda
Charles Masembe
Department of Zoology, Entomology and Fisheries Sciences, Makerere University, P.O. Box 7062, Kampala, Uganda
LiveDNA: 256.30565
Journal of Fisheries and Aquatic Science
Year: 2020 | Volume: 15 | Issue: 2 | Page No.: 59-68







ABSTRACT
Background and Objective: Human-induced physical and environmental barriers especially hydropower dams and dense anoxic vegetation along riverine-lacustrine connectivity are known to isolate and fragment fish populations, leading to variation. This study explored possible morphological variations among populations of Barbus altianalis from Uganda with the intent to provide a more cost-effective and reliable platform for elucidating the stocks under combined environmental and anthropogenic influence as a tool for improved fisheries management and conservation. Materials and Methods: Multivariate analysis of morphometric variables was performed to determine the morphological variation and discreteness among three populations of B. altianalis (N = 251) from the lacustrine (Lake Edward) and riverine (River Nile and Kazinga channel) systems in Uganda. Morphological variation was determined using a multivariate analysis of advanced truss and non-truss network systems on all individuals collected from commercial catches. Results: There was a significantly (p<0.001) higher level of heterogeneity among populations. Group means on the DFA1 showed that River Nile (-0.89) separated furthest from Lake Edward (0.79) and the Kazinga channel (0.68). River Nile had the highest number (80.5%) of individuals correctly classified while Lake Edward (67.2%) and Kazinga channel (60.5%) had more misclassifications between them, an indication that they could be much harder to separate. Conclusion: The observed patterns of variation were consistent with the geographical locations of the different water bodies, that the isolating mechanisms are subjecting the different populations to different selection pressure hence different phenotypic responses by the populations.
PDF Abstract XML References Citation
How to cite this article
Constantine Chobet Ondhoro, Gregory E. Maes, Nalukenge Winnie Nkalubo, John K. Walakira, Matthew Tenywa Mwanja, Egulance Ganda, Godfrey Kawooya Kubiriza, Jackson Efitre and Charles Masembe, 2020. Multivariate Morphometric Analysis of Riverine and Lacustrine Barbus altianalis populations in Uganda. Journal of Fisheries and Aquatic Science, 15: 59-68.
DOI: 10.3923/jfas.2020.59.68
URL: https://scialert.net/abstract/?doi=jfas.2020.59.68
DOI: 10.3923/jfas.2020.59.68
URL: https://scialert.net/abstract/?doi=jfas.2020.59.68
INTRODUCTION
The barbel fish Barbus altianalis, locally known as “Kisinja” in Uganda, is a commercially important freshwater fish that grows to a large size (maximum reported size about 90 cm total length) and is considered a culinary delicacy among riparian communities in East Africa1,2. This cyprinid is an omnivore, widely distributed in Lakes Kyoga, Edward, Albert and George, as well as in the River Nile and others that drain into Lake Victoria3. Although the taxonomic status of B. altianalis is debatable, we used the traditional nomenclature which recognizes the species as B. altianalis4-6.
The populations of B. altianalis are under threat from over fishing, erecting hydropower dam barriers along migration routes and degraded aquatic ecosystems7. However, various conservation aspects such as population structure, variability and resilience to harvest pressure have not been satisfactorily evaluated8. Prior to the construction of the Owen Falls dam on the River Nile in 1954, large congregations of B. altianalis were common at the foot of Ripon falls1-8. However, due to over fishing and environmental changes, coupled with vulnerable reproductive and feeding behavior, the species is under threat of extinction5-7. Being a partial batch spawner, the fish migrates up and downstream throughout the year to spawn. However, this behavior exposes it to overexploitation along the migration routes9,10. While off-shore trawl surveys through Lake Victoria in 1975 and 1985 showed fluctuations in Catch per Unit Effort (CPUE) of between 0.2 to 0.5 kg/h5,8, the current CPUE in the Victoria and Edward basins is estimated to average 0.06 and 1.50 kg/h respectively11-13.
In addition, physical barriers including hydropower dams along the Nile River and anoxic papyrus swamps along the riverine-lacustrine borders between Lakes Victoria and Albert have isolated populations of B. altianalis4,2.
Examination of systemic morphometric variation, therefore, gives prospects for population structure of B. altianalis stocks from different water bodies. This is premised on the affirmation that variations among fish populations provide a basis for fish population structure and a foundation for documenting environmentally induced variance14. Furthermore, morphological divergence is important in fish stock management as a marker of changes in environmental quality associated with evolution. Knowledge of fish morphometric variations therefore, facilitates the development of optimal strategies for efficient management of stocks, reacting independently to fishing pressure and other ecosystem dynamics14,15. Changes in body forms of fish populations exposed to different environmental conditions could be used to improve fisheries management and conservation16. However, phenotypic features, more so the morphology of fishes are interlinked and therefore, use of single parameter as had been before, could be misleading16. For example, the shortcoming with single morphological parameters is related to the existence of temporary morphotypes such as deformities originating from either early ontogenetic development such as juvenile pigmentation in some tilapia species, commonly known as “Tilapia mark”, or those inflicted by type II cannibalism, common with African catfish17. Therefore, unless a given morphometric feature is known to be genetically linked, multivariate morphometric approach remains the most reliable technique for inferring population morphometric variations18-23. The method has been widely applied in a number of recent studies to study taxonomic contradictions on fish stocks20,15,16-24. Application of multivariate morphometric analysis on the current stocks of B. altianalis is a cost-effective alternative to generate valuable management information for monitoring and designing appropriate conservation strategies for different water bodies23.
The aim of this study was therefore, to examine the morphometric variation among populations of B. altianalis from River Nile, Lake Edward and Kazinga channel Uganda using multiple morphometric variables to inform conservation and management of the species.
MATERIALS AND METHODS
Study area: The study was carried out in selected fish landing sites along River Nile, Lake Edward and the Kazinga channel in 2013.
Fish specimens used in this study were obtained from four landing sites in the River Nile (upstream - Ripon and downstream -Kiira, Kalange and Bukeka) of the Owen Falls dam. On Lake, Edward samples were obtained from Katwe and Kayanja landing sites. Fish were also sampled from two landing sites of Katunguru and Kazinga in the Kazinga channel, upstream Lake Edward, Uganda. Given its location, the fishers at the Ripon landing site conduct their fishing activities both in the lacustrine and the riverine systems. The fisherfolk, fishing vessels and gears in Kiira, Kalange and Bukeka landing sites are restricted to fishing within the riverine system between the dams along River Nile. Katwe, Kayanja, Katunguru and Kazinga are situated in water bodies located within Queen Elizabeth National Park, where fishing activities have some level of restriction given their location in a protected area (Fig. 1).
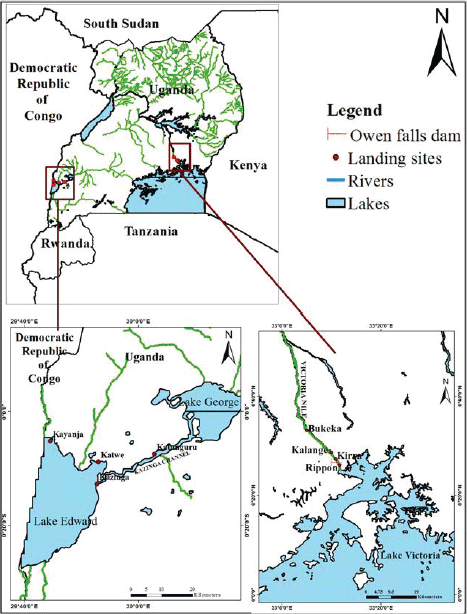 | |
| Fig. 1: | Map indicating the location of the sampled landing sites in River Nile, Lake Edward and Kazinga channel. Source: personal data |
Sample and morphometric data collection: Fish specimens were obtained from commercial catches in December 2013. Samples from all the landing sites from the same water body were pooled into one unit to generate three discrete populations of the River Nile- Population one, Lake Edward- population two and Kazinga channel- population three. Pooling of samples was done mainly to limit the study to population level that was designated by water bodies under investigation. Fish samples were collected within the same period (December 2013) to minimize variation contributed by seasonal variation in food, water quality, reproduction and other allochthonous influence25.
A truss dimension of each fish was constructed from the left side of the fish, using Vernier callipers, following landmarks adopted from the previous studies26,27,15. A total of 13 trusses and 6 non-truss morphometric variables were measured on each specimen to the nearest 0.1 cm. Truss and non-truss network systems were adopted because they are able to discriminate phenotypic stocks by the configuration of the constructed landmarks that cover the entire fish body with no loss of information and being more sensitive to changes26-29 (Table 1).
Construction of truss and non-truss dimensions: A truss dimension is series of measured distances calculated between landmarks that form a regular pattern of connected quadrilaterals or cells across the body form as described by Strauss and Bookstein27; Turan15. The truss procedure used for this study was based on 10 landmarks, whereby, the truss dimensions were constructed by interconnecting landmarks to form a total of 13 measurements.
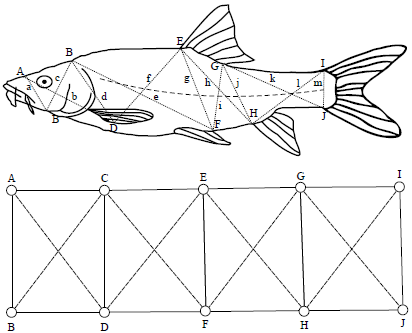 | |
| Fig. 2: | Data recording procedure, Top 19 distance parameters recorded on vernier calipers, Top marked in lower case letters are distances measured between landmarks and marked in upper case letters are the morphometric land marks in B. altianalis |
a: Snout, b: Ventral junction of operculum junction, c: Supra orbital, d: Lower pectoral fin insertion, e: Anterior dorsal fin insertion, f: Posterior end of Pelvic Fin insertion, g: Posterior end of dorsal fin insertion, h: Anterior end of anal fin insertion, i: Dorsal caudal fin Insertion, j: Ventral caudal fin insertion, k: Posterior dorsal fin insertion to ventral caudal fin insertion, l: Dorsal caudal fin insertion to ventral caudal fin insertion, m: Anterior anal fin insertion to dorsal caudal fin insertion, Bottom: Geometric representation of the data recording procedure, Each single cell is determined by four sides and two diagonals, Modified from Corti et al.30 | |
| Table 1: | Truss and non-truss dimensions measured for the morphometric analysis of B. altianalis collected from River Nile, Lake Edward and Kazinga channel as in Fig. 2 |
 | |
The measurements were conducted by inserting a pin on an insertion of the first landmark (A-snout) followed by a similar insertion on the second landmark (B-Ventral junction of operculum). The readings were then taken by vernier callipers along a straight edge. The same procedure were repeated for all other measurements along different landmarks. The measurements of non-truss (measured distances along the length of a single landmark i.e. fin length), were done by inserting a pin on a point of attachment of the fin to the trunk and a vernier calliper then run through a straight edge to the tip and a record of the readings was taken (Fig. 2)26,27.
Data analysis: Size-dependent morphometric variables were log-transformed to normalize the dataset. The log-transformed data were each correlated to fish total length to establish the relationship between total length and each morphometric variables. After establishing the relationship between total length and morphometric variables, the log-transformed variables were then corrected to eliminate the correlation of the variables with size. This was done by adopting an allometric method suggested by Elliot et al.31, Turan15 and Mir et al.16, using the formula:
Madj = M (Ls/Lo) b
where, M is the original truss and non-truss measurement of the morphometric variable, Madj is the size adjusted truss and non-truss measurement of the morphometric variable, Lo is the total length of the fish, Ls is the overall mean total length and b was estimated for each variable from the observed data as the slope of the regression of log M on log Lo using the fish from every population.
The results derived from the allometric method were confirmed by testing the correlation between transformed variables and total length (Lo).
The differentiation among the populations was based on the percentage of correctly and incorrectly classified fish. The degree of intermingling among the populations as indicated by the number of misclassified individuals. The first and the second F-ratios that test both canonical correlations were explored for significance to describe the differences among the three populations. The discriminant function for dimensions one and two was explored.
The separation among the populations was explored using a discriminant function analysis score plot.
In order to identify the contribution of each morphometric variable to the observed variation, loading plot that provides the graphical way of looking at the standardized discriminant function coefficient was used. Log transformed morphometric variables were used for the production of the loading plot and the correlation table. The transformed variables are therefore, denoted by capital letters to differentiate it from ‘adj’ (adjusted).
Statistical analysis: Morphometric truss and non-truss parameters were subjected to Discriminant Function Analysis (DFA) to explore possible separation among the three populations using the software STATA, version 12. The percentage of correctly classified individuals was used as a measure of the morphological distinctness of the samples pooled within groups. Correlations between variables and discriminant scores were used to interpret canonical variants among populations16. Significant (p<0.001) morphometric differences and the separation of the three populations away from each other were confirmed by Wilk’s lambda (λ) test31.
RESULTS
Fish population characteristics: A total of 251 freshly caught fish samples were obtained; 113 specimens from River Nile, (only four from the lake-river interface upstream from the Owen Falls hydropower dam and 109 downstream the dam); 61 were from Lake Edward while 77 from Kazinga channel (Table 2).
All the morphometric characters were linearly correlated to total length, indicating significant relationship between respective variables with size before allometric transformation (Table 3).
The size correlated morphometric variables showed no relationship with size after allometric transformation. None of the 19 morphometric variables showed a significant correlation with total length, indicating the size effect was successfully removed from the dataset as showed by standardized discriminant function coefficients below. None of the variables showed a significant correlation of >0.9 in both function one (DFA 1) and two (DFA 2) Table 4.
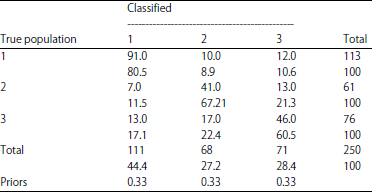
Morphometric variation: The re-substitution classification table (confusion matrix) indicated population one had the highest number (80.53%) of individuals classified correctly while populations two (67.2%) and three (60.5%) had more misclassifications between them indicating that the latter groups could be harder to separate (Table 5).
| Table 2: | Sampling sites, fishing effort and size (±SD) of B. altianalis from River Nile, Lake Edward and Kazinga |
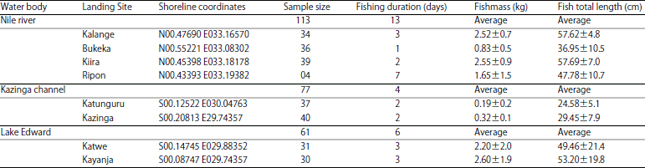 | |
Among the 19 variables, orbital distance, pelvic fin and caudal fin lengths (OBDadj, PVFLadj and CFLadj) respectively, were not significantly (p>0.001) different in population one. Wilk’s lambda (λ) test of DFA showed significant (p<0.001) differences in morphometric variables and confirmed significant separation of the three populations from each other (Table 6).
The results of the factor analysis between the three populations indicated that the two factors (DFA1 and DFA2) collectively explained 100% of the total morphometric variation. With exception of OBDadj, PVFLadj and CFLadj, all other variables had higher loadings. Those with higher loadings accounted for 84% of the variance than the second-factor loading accounted for 16% of the variance (Table 7).
The group means on the first function showed that population one (River Nile (-0.8869) separated farthest from populations two (Lake Edward (0.796) and three (Kazinga channel (0.6796). The score plot illustrates the populations being reasonably well separated from each other with minimum intermingling.
| Table 3: | Regression models of TL against morphometric variables of B. altianalis |
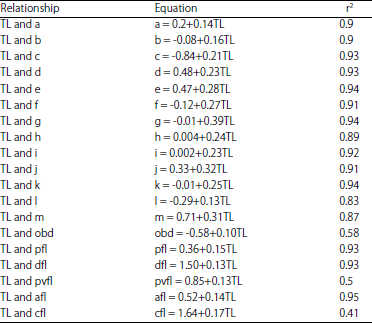 | |
| Regression model of TL, truss and non-truss variables (p<0.0 5), TL is total length of each fish sample obtained from respective landing sites, The different letters marked in lower case represent respective truss and non-truss variables measured from each fish sample as elaborated in Table 1 above | |
There is clear separation in the second dimension between the lacustrine (Lake Edward) from riverine (River Nile and Kazinga channel) but with insubstantial overlap (Fig. 3).
Observations of the loading plot indicated variables OBDadj, PVFLadj and CFLadj near the origin, implying that they provide almost no discriminating ability in comparison to the other discriminating variables (Fig. 4).
| Table 4: | Coefficients of correlation of adjusted morphometric variables with discriminant functions of B. altianali |
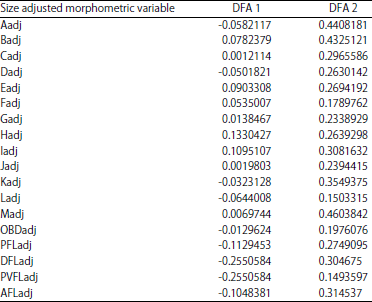 | |
The different letters marked in upper case represent respective size adjusted/corrected truss and non-truss variables measured from each fish sample as elaborated in Table 1 above | |
| Table 5: | Re-substitution classification summary of three populations of B. altianalis |
 | |
| Table 6: | Wilk’s lambda tests (function 1 and 2) for the verifying and classifying differences among three populations of B altianalis |
| Table 7: | Canonical linear discriminant analysis of morphometric variables of B. altianalis from River Nile, Lake Edward and Kazinga channel |
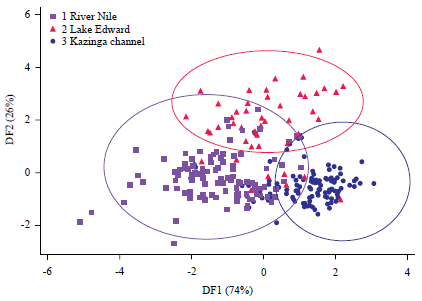 | |
| Fig. 3: | DFA graph of 19 morphometric variables used for B. altianalis from River Nile, Lake Edward and Kazinga channel. Labels 1, 2 and 3 represent River Nile, Lake Edward and Kazinga Channel populations respectively |
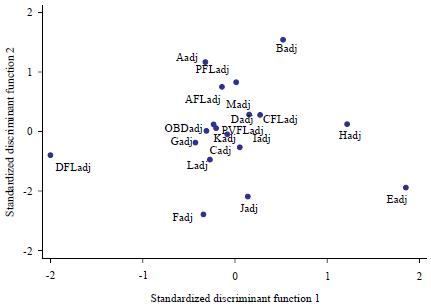 | |
| Fig. 4: | Standardized discriminant factor loading plot (source; personal data) |
| a: Snout, b: Ventral junction of operculum junction, c: Supra orbital, d: Lower pectoral fin insertion, e: Anterior dorsal fin insertion, f: Posterior end of pelvic fin insertion, g: Posterior end of dorsal fin insertion, h: Anterior end of anal fin insertion, i: Dorsal caudal fin Insertion, j: Ventral caudal fin insertion, k: Posterior dorsal fin insertion to ventral caudal fin insertion, l: Dorsal caudal fin insertion to ventral caudal fin insertion: m: Anterior anal fin insertion to dorsal caudal fin insertion, Here variables are denoted by uppercase letters to differentiate them from ‘adj’ (adjusted) | |
DISCUSSION
There was no significant overlap among the River Nile, Lake Edward and the Kazinga channel populations. Intersect among the populations as revealed by the score plot, collectively showed no intermingling, implying significant heterogeneity among populations (Fig. 3). The observed variations were largely explained by both truss and non-truss morphometric variables. Exceptions were with the distance between the eyes, referred to as orbital distance (OBDadj), pelvic fin (PVFLadj) and caudal fin (CFLadj) lengths.
Group means of the first function further indicated that the River Nile population (-0.89) was further separated from Lake Edward (0.79) and Kazinga channel (0.68). Similarly, the matrix also puts River Nile population much further morphologically from the Lake Edward and Kazinga channel counterparts. Over 80.53% of individuals were correctly classified in River Nile, contrary to 67.2 and 60.5% in Lake Edward and Kazinga channel populations respectively. The smaller margin of variance between Lake Edward and Kazinga channel populations implies the two populations could be much harder to separate. The variations were however, more concentrated on the anterior side of the specimens. As indicated in Table 4, the snout length and other parts of the head region contributed the largest variance in dimensions one (-0.06) and two (0.44).
Similar phenotypic variations among populations had earlier been observed with the variation in snout configuration in samples obtained from Lakes Edward and Victoria basins6,2. However, these earlier studies relied mainly on single parameter (snout), contrary to the current study which used multiple variables.
Morphological variation among fish populations due to deviation in head and middle regions of the fish are useful indicators of differentiation among respective populations, mainly as a result of geographical and topographical deviations that compel the different populations to adapt differently to physiological and morphological requirements of the habitat. Moreover, variation in the snout form reveals differences in the methods of obtaining food, following dietary shifts among populations. These forms therefore, induce evolution of body shape, influencing selection of food and prey catching mechanisms16,32.
Whereas the posterior part of the fish did not significantly contribute to the variation, the stability of the caudal (CFLadj) and the pelvic (PVFLadj) fin lengths across different water bodies shows the traits have been conserved across populations. Notably, in bony fishes, the caudal fins are mainly used for propulsion to move the fish forward, while the pelvic fin provides the ability during sharp turns, quick and vertical movement along the water column. As reported by Ogutu-Owhayo et al.33, the smothering effects of siltation of River Katonga that used to provide a direct connection between Lakes Victoria and Edward basins blocked upstream migration of aerophilic Barbus altianalis, rendering them more or less obligate lacustrine or riverine species.
Similarly, as observed, the low abundance of B. altianalis upstream Owen-falls dam is characteristic of the species that is extinct from the lacustrine side of River Nile. Owen-falls hydropower dam and the dense anoxic conditions along the riverine habitats have barred the fish from migrating; subjecting individuals blocked upstream the reservoir to high mortality rates due to predation and high fishing pressure. Recent study on B. altianalis fishery reveals deteriorated environmental quality and high fishing effort against the species across River Nile, Lake Edward and Kazinga channel34. Similarly, Ochumba and Manyala11 observed that, habitat alteration through anthropogenic processes caused the growth of dense papyrus at the mouth of River Sundo-Miru in Kenya, hampering up and downstream migration among potamodromous fish species. Likewise, such vegetation also limited the entry of newly hatched fry into the lake from the river, predisposing the fry to higher predation pressure and lack of suitable diet29.
These finding may be useful to fisheries management and aquaculture production by providing baseline information on inter-population morphological variation in B. altianalis. Coupled with the findings of Ondhoro et al.34 that presents data on fishing effort and environmental parameters, this information could assist in generating guidelines for location-specific management based on the prevailing environmental conditions and stock structure. A similar morphological study was applied to identify hatchery against wild stocks of Coho salmon (Oncorhynchus kisutch)17,18,20. Spawning experiments with B. altianalis concur with the current findings following survival variation in several spawning trials of wild specimens collected from the same study sites10. Individuals from the Lake Victoria basin had better spawning characteristics than their Lake Edward basin counterparts that experienced high broodstock mortality and very low larval survival rates10.
Despite these apparently good findings, the shortcoming with morphological characterization still rests in its inability to resolve variation due to genetic influence. The use of nuclear markers such as SNP analysis is recommended for future population studies.
CONCLUSION
This study shows B. altianalis stocks in riverine and lacustrine systems are morphologically distinct and is very rare in the Lake Victoria- River Nile interface mainly due to its stocks being locked upstream by the dam, facing high fishing pressure and low rate of recruitment. This information is therefore, relevant for the development of appropriate capture fisheries and ecological monitoring policy based on periodic integrated multivariate approaches.
SIGNIFICANCE STATEMENT
This study discovers and provides the first construction of truss and non-truss dimensions for B. altianalis multivariate morphological discrimination. The study generated baseline data for monitoring the effects of isolating mechanisms such as hydropower dams on migrating fish species. With the emergency of more hydropower dams along the River Nile, the information generated in this study will provide a baseline for future monitoring of Barbus populations in Uganda and beyond.
ACKNOWLEDGMENT
Financial support for this study was provided through the National Fisheries Resources Research Institute-Uganda, by the Agricultural Technology Agri-business Advisory Services (World Bank ATAAS) Project (P.109224), administered by the National Agricultural Research Organization (NARO). Director of Research Buginyanya ZARDI (Dr. Lawrence Owere) provided additional financial support in processing the manuscript. We acknowledge the efforts of EA. Nyboer of McGill University Canada, for proof-reading the manuscript. Special thanks to the field staff; R. Wasswa and P. Odongo for guidance and coordination with fish folk in Kasese and Jinja districts respectively. M. Turyashemererwa of Aquaculture Research and Development Centre (ARDC) Kajjansi offered information and communication technology (ICT) support.
REFERENCES
- Boulenger, G.A., 1900. Descriptions of new reptiles and batrachians from Borneo. Proc. Zool. Soc. London, 69: 182-187.
CrossRefDirect Link - Greenwood, P.H. and K.S. Thomson, 1960. The pectoral anatomy of Pantodon buchholzi peters (a freshwater flying fish) and the related osteoglossidae. Proc. Zool. Soc. London, 135: 283-301.
CrossRefDirect Link - Chemoiwa, E.J., R. Abila, A. Macdonald, J. Lamb, E. Njenga and J.E. Barasa, 2013. Genetic diversity and population structure of the endangered ripon barbel,Barbus altianalis (Boulenger, 1900) in Lake Victoria catchment, Kenya based on mitochondrial DNA sequences. J. Appl. Ichthyol., 29: 1225-1233.
CrossRefDirect Link - Greenwood, P.H. and K.S. Thomson, 1960. The pectoral anatomy of Pantodon buchholzi peters (a freshwater flying fish) and the related osteoglossidae. Proc. Zool. Soc. London, 135: 283-301.
CrossRefDirect Link - De Graaf, M., M.A.M. Machiels, T. Wudneh and F.A. Sibbing, 2004. Declining stocks of Lake Tana's endemic Barbus species flock (Pisces; Cyprinidae): Natural variation or human impact? Biol. Conservat., 116: 277-287.
CrossRefDirect Link - Masese, F.O., P.O. Raburu, B.N. Mwasi, and L. Etiegni, 2012. Effects of deforestation on water resources: integrating science and community perspectives in the Sondu-Miriu River Basin, Kenya. In: New advances and contributions to Forestry Research, Oteng-Amoako, A.A., pp: 1–18.
CrossRefDirect Link - Rutaisire, J., B. Levavi-Sivan, C. Aruho and C.C. Ondhoro, 2015. Gonadal recrudescence and induced spawning in Barbus altianalis. Aquacult. Res., 46: 669-678.
CrossRefDirect Link - Ochumba, P.B.O. and J.O.M. Ala, 1992. Distribution of fishes along the Sondu-Miriu River of Lake Victoria, Kenya with special reference to upstream migration, biology and yield. Aquacult. Fish. Manage., 23: 701-719.
CrossRefDirect Link - Ntiba, M.J., W.M. Kudoja and C.T. Mukasa, 2001. Management issues in the Lake Victoria watershed. Lakes Reservoirs: Res. Manage., 6: 211-216.
CrossRefDirect Link - Beger, R.D., J.F. Young and H. Fang, 2004. Discriminant function analyses of liver-specific carcinogens. J. Chem. Inf. Comput. Sci., 44: 1107-1110.
CrossRefDirect Link - Turan, C., 2004. Stock identification of mediterranean horse mackerel (Trachurus mediterraneus) using morphometric and meristic characters. ICES J. Mar. Sci., 61: 774-781.
CrossRefDirect Link - Mir, J.I., F.A. Mir, S. Chandra and R.S. Patiyal, 2013. Pattern of morphological variations in Alghad snowtrout, Schizopyge niger (Heckel 1838) from Kashmir Himalaya using truss network analysis. Ichthyol. Res., 60: 256-262.
CrossRefDirect Link - Swain, D.P. and C.J. Foote, 1999. Stocks and chameleons: the use of phenotypic variation in stock identification. Fish. Res., 43: 113-128.
CrossRefDirect Link - Klingenberg, C.P., 2010. Evolution and development of shape: integrating quantitative approaches. Nat. Rev. Genet., 11: 623-635.
CrossRefDirect Link - Cadrin, S.X., 2000. Advances in morphometric identification of fishery stocks. Rev. Fish Biol. Fish., 10: 91-112.
CrossRefDirect Link - Monet, G., A. Uyanik and A. Champigneulle, 2006. Geometric morphometrics reveals sexual and genotypic dimorphisms in the brown trout. Aquat. Living Resour., 19: 47-57.
CrossRef - Silva, A., 2003. Morphometric variation among sardine (Sardina pilchardus) populations from the northeastern Atlantic and the western Mediterranean. ICES J. Mar. Sci., 60: 1352-1360.
CrossRefDirect Link - Mekkawy, I.A.A. and A.S. Mohammad, 2011. Morphometrics and meristics of the three epinepheline species: Cephalopholis argus (Bloch and Schneider, 1801), Cephalopholis miniata (Forsskal, 1775) and Variola louti (Forsskal, 1775) from the red sea, Egypt. J. Biol. Sci., 11: 10-21.
CrossRefDirect Link - Finlay, S. and N. Cooper, 2015. Morphological diversity in tenrecs (Afrosoricida, Tenrecidae): comparing tenrec skull diversity to their closest relatives. PeerJ.
CrossRefDirect Link - Fleming, I.A., B. Jonsson and M.R. Gross, 1994. Phenotypic divergence of sea-ranched, farmed, and wild salmon. Can. J. Fish. Aquat. Sci., 51: 2808-2824.
CrossRefDirect Link - Strauss, R.E. and F.L. Bookstein, 1982. The truss: Body form reconstructions in morphometrics. Syst. Zool., 31: 113-135.
CrossRefDirect Link - Chemoiwa, E.J., R. Abila, A. Macdonald, J. Lamb, E. Njenga and J.E. Barasa, 2013. Genetic diversity and population structure of the endangered ripon barbel,Barbus altianalis (Boulenger, 1900) in Lake Victoria catchment, Kenya based on mitochondrial DNA sequences. J. Appl. Ichthyol., 29: 1225-1233.
CrossRefDirect Link - Greenwood, P.H. and K.S. Thomson, 1960. The pectoral anatomy of Pantodon buchholzi peters (a freshwater flying fish) and the related osteoglossidae. Proc. Zool. Soc. London, 135: 283-301.
CrossRefDirect Link - Corti, M., R.S. Thorpe, L. Sola, V. Sbordoni and S. Cataudella, 1988. Multivariate morphometrics in aquaculture: a case study of six stocks of the common carp (Cyprinus carpio) from Italy. Can. J. Fish. Aquat. Sci., 45: 1548-1554.
CrossRefDirect Link - Elliott, N.G., K. Haskard and J.A. Koslow, 1995. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of Southern Australia. J. Fish Biol., 46: 202-220.
CrossRefDirect Link - Noatch, M.R and C.D. Suski, 2012. Non-physical barriers to deter fish movements. Environ. Revision, 20: 11-12.
CrossRefDirect Link - Ogutu-Ohwayo, R., K. Odongkara, W. Okello, D. Mbabazi, S.B. Wandera, L.M. Ndawula and V. Natugonza, 2013. Variations and changes in habitat, productivity, composition of aquatic biota and fisheries of the Kyoga lake system: lessons for management. Afr. J. Aquatic Sci., 38: 1-14.
CrossRefDirect Link - Ondhoro, C.C., C. Masembe, G.E. Maes, N.W. Nkalubo and J.K. Walakira et al., 2017. Condition factor, length-weight relationship and the fishery of Barbus altianalis (Boulenger 1900) in Lakes Victoria and Edward basins of Uganda. Environ. Biol. Fishes, 100: 99-110.
CrossRefDirect Link