
Dewi Nurhayati
Post Graduate School of Aquaculture Science, Bogor Agricultural University, Indonesia
Widanarni
Department of Aquaculture, Faculty of Fisheries and Marine Science, Bogor Agricultural University, Indonesia
Munti Yuhana
Department of Aquaculture, Faculty of Fisheries and Marine Science, Bogor Agricultural University, Indonesia
Journal of Fisheries and Aquatic Science
Year: 2015 | Volume: 10 | Issue: 1 | Page No.: 13-23







ABSTRACT
The experiment was conducted to examine the effects of dietary administration of probiotic SKT-b (Vibrio alginolyticus ) and oligosaccharide from sweet potato ( Ipomoea batatas L. ) on growth performance and immune responses of white shrimp (Litopenaeus vannamei). Shrimp (0.33±0.02 g) was fed with basal diet (control without challenge test (-) and control with challenge test (+)), supplemented with combination of probiotic and prebiotic (probiotic 0.5% + prebiotic 1% (A), probiotic 1% + prebiotic 2% (B), probiotic 2% + prebiotic 4% (C)) for 30 days of culture. After feeding experiment, five shrimp per tank were sampled for bacterial quantification and immunity determination. Then all the shrimp rest were challenged by infectious myonecrosis virus and V. harveyi. After 30 days, Daily Growth Rate (DGR) and Food Conversion Ratio (FCR) of the shrimp were significantly better in treatment B than the control but weren’t significantly different (p>0.05) in other treatment. Other than that, phenoloxidase (PO) activity and Total Haemocyte Count (THC) of the shrimp in all synbiotic treatments were higher from the control group. After co-infection challenged test, PO, THC and Respiratory Burst (RB) activity of the shrimp in all synbiotic treatments were higher from control (+). Besides that, lowest survival shrimp occurred in the control (+) was significantly different compared with all synbiotic treatments.
PDF Abstract XML References Citation
Received: November 25, 2014;
Accepted: January 07, 2015;
Published: March 16, 2015
How to cite this article
Dewi Nurhayati, Widanarni and Munti Yuhana, 2015. Dietary Synbiotic Influence on the Growth Performances and Immune Responses
to Co-Infection with Infectious Myonecrosis Virus and Vibrio harveyi in Litopenaeus
vannamei. Journal of Fisheries and Aquatic Science, 10: 13-23.
DOI: 10.3923/jfas.2015.13.23
URL: https://scialert.net/abstract/?doi=jfas.2015.13.23
DOI: 10.3923/jfas.2015.13.23
URL: https://scialert.net/abstract/?doi=jfas.2015.13.23
INTRODUCTION
White shrimp (Litopenaeus vannamei) is one of the most economically important commodities in fisheries. According to FAO, Indonesia is at number four in world shrimp production after China, Thailand and Vietnam. Indonesia in 2009 set a target to increase white shrimp production by 209-511% thousand tons in 2014 (Directorate General of Aquaculture, 2010).
One of the problems in white shrimp intensive culture is the outbreaks of infectious diseases, particularly those caused by viruses and bacteria. In the last few years, many shrimp farms in Indonesia have been infected by infectious myonecrosis virus (IMNV). That had been reported for the first time in Indonesia since the 2006s (Senapin et al., 2007). While bacterial disease that infected the white shrimp were caused by the bacterium Vibrio harveyi (Austin and Zhang, 2006). Vibrio harveyi has been recognized as a serious pathogen for various aquaculture organisms world wide (Soto-Rodriguez et al., 2012).
Previous studies showed that shrimp diseases not only caused by a single infection. The presence of co-infection or infection with multiple pathogens in white shrimp may accelerate and enhance shrimp mortality. Concurrent viral and bacterial infections in shrimp have been reported in White spot syndrome virus (WSSV) Vibrio campbelli (Phuoc et al., 2009), WSSV-V. harveyi (Phuoc et al., 2009), as well as IMNV-V. harveyi (Hasan, 2011). According to Hasan (2011), co-infection IMNV with different doses of V. harveyi could accelerate early mortality and increased mortality compared with IMNV single infection.
Several methods have been applied to control the disease. The application of antibiotic or other chemical in aquaculture may cause the risk of drug resistant pathogens (Balcazar et al., 2006). An alternative that is used to control the disease was by administration of synbiotic to prepare shrimp innate immunity by increasing the resistance of shrimp against pathogens. Synbiotic is a balanced combination of probiotics and prebiotics. Probiotics are considered as a live microbial food supplement that improves microbial balance and health of the host and their culture habitats (Verschuere et al., 2000). A prebiotic is defined as a non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or the activity of specific health promoting bacteria that can improve the host health (Ringo et al., 2010). Synbiotic applications of prebiotics and probiotics have been shown to significantly increase diseases resistance, modulate gastro-intestinal microbiota and stimulate the immune response in shrimp (Li et al., 2009), lobster (Daniels et al., 2010), sea cucumbers (Zhang et al., 2010), yellow croaker (Ai et al., 2011) and koi (Lin et al., 2012).
The present study aimed to determine the most optimum combination of probiotic SKT-b (V. alginolyticus) and oligosaccharide extracted from sweet potato (Ipomoea batatas L.) on the growth performance, immune response and disease resistance to co-infection with IMNV and V. harveyi of shrimp (L. vannamei).
MATERIALS AND METHODS
Probiotic and oligosaccharides: The SKT-b probiotic used in this study was previously isolated from the culture medium of Skeletonema sp. in shrimp hatchery environment of Labuan Banten, Indonesia (Widanarni et al., 2003). SKT-b bacteria was marked with resistant to the antibiotic rifampicin (SKT-bR) as a molecular marker at a dose of 50 μg mL-1 (Widanarni et al., 2003). Probiotic bacteria was cultured on sea water complete-agar (SWC, 1.7 bacto agar, 5 g bacto peptone, 1 g yeast extract, 3 mL glycerol, 750 mL sea water and 250 mL distilled water) and then transferred to a 25 mL SWC-broth (without bacto agar) with a shaking incubator at 1600 rpm for 18 h at 29°C. Cell culture was harvested by centrifuging at 5000 rpm for 15 min and washing two times with phosphate buffer saline (PBS, NaCl 0.8 g, KH2PO4 0.2 g, Na2HPO4 1.5 g, KCl 0.2 g and 1000 mL distilled water). Culture purity and identification were routinely checked during this investigation by spreading plate method (Madigan et al., 2003).
Oligosaccharides used in this study extracted from sweet potato (I. batatas L.) (Marlis, 2008; Muhtadi, 1989). The ingredients in Oligosaccharides (TDS, total dissolved solid 5%) were determined by High Performance Liquid Chromatography (HPLC) and expressed in percentage as follows: Inulin 1.115%, fructooligosaccharides (FOS) 1.015%, galactooligosaccharides (GOS) 1.488%.
Diet preparation: Basal diet was formulated to contain approximately 40% crude protein and 8% crude lipid which was suitable for the growth of this shrimp. Five experimental diets were prepared with different combinations of probiotic SKT-b and sweet potato extract: Control without challenge test (C-) and control with challenge test (C+) (basal diet used as the control), A (probiotic 0.5% + prebiotic 1%), B (probiotic 1% + prebiotic 2%), C (probiotic 2% + prebiotic 4%). The probiotic (109 CFU mL-1) and prebiotic with TDS 5% (Marlis, 2008) were provided in a liquid form. Doses of probiotic SKT-b and prebiotic were supplemented into the basal diet followed by manual mixing. The mixture was spread out and air dried by fan at room temperature. Diets were then stored in clean plastic bags at 4°C until use.
Culture condition: Experimental white shrimp at post-larval stadia (PL) 15 was obtained from a commercial hatchery (Banten, Indonesia) and acclimated in the laboratory for a 21 days before experimentation. Shrimp initial average body weight of 0.33±0.02 g was randomly assigned to triplicate tanks (30×35×40 cm; volume 0.3 m3) stocked with a density of 15 shrimp per tank. Shrimp fed four times daily at 10% biomass at-satiation. The water temperature in the culture tanks was ranged at 27-29°C, 29-32‰ in salinity, 7.4-7.5 in pH, 4.5-6.5 in Dissolved Oxygen (DO) and 0.005-0.016 mg L-1 of ammonia-nitrogen.
Growth and feed conversion ratio: After 30 days experiment, the total numbers of shrimp were counted and the mean of body weights were measured. Based on these weight of shrimp and counting the number of shrimp, Daily Growth Rate (DGR) and Feed Conversion Ratio (FCR) were calculated using the following equations:
where, DGR is the specific growth rate in weight (% g), Wt and Wo are shrimp weight at current time (t) and at the beginning of the experiment (o), respectively, whereas t is the number of rearing days (day). FCR is food conversion ratio, F is the total dry food consumed (g) and Bt, Bm and Bo is the total biomass of the shrimp (g) at current time (t), due to mortality of shrimp (m) and at the beginning of the experiment (o), respectively, whereas t is the number of rearing days (day).
Bacterial quantification: The intestines of shrimp (approximately 0.1 g in weight) collected from each tank and then homogenized in 0.9 mL sterile PBS. Bacterial quantifications for homogenized liquid were made using PBS, followed by plating on medium agar. Sea Water Complete (SWC) and vibrio selective medium thiosulphate citrate bile-salts sucrose (TCBS, HIMEDIA) agar were used for total viable bacterial counts (TBC), presumptive vibrio counts (VBC) and the SKT-bR counts. After 24 h of incubation at 27°C, the number of bacterial colonies was counted and the amount of bacteria was expressed as Colony Forming Unit (CFU g-1) (Madigan et al., 2003).
Challenged test: On day 30 of the feeding experiment, ten shrimp remained in the tanks were challenged with IMNV by injecting infected-shrimp (L. vannamei). IMNV was collected from BBAP Situbondo located in East Java, Indonesia. The V. harveyi (MR 5339) which has been tagged by rifampicin resistant was a collection of Fish Health Laboratory, Department of Aquaculture, Bogor Agricultural University, Indonesia.
Shrimp infected co-infection IMNV and V. harveyi MR 5339 by injection (Phuoc et al., 2009). Ten shrimp of each tanks were injected intramuscularly with 0.1 mL IMNV. After 72 h injection of IMNV, shrimp was injected intramuscularly with PBS containing 103 CFU live V. harveyi from a 24 h culture in SWC medium at 29°C. Shrimp mortality following challenge test was observed for 7 days. Confirmation of the presence of virus in the body IMNV shrimp performed by Polymerase Chain Reaction (PCR) analysis.
Immune parameter assay: Haemolymph was collected from the ventral-sinus with syringe containing anticoagulant solution. The same samples collected from the five shrimp in the same tank were mixed and used as one unit. The anticoagulant solution (10 mM of Ethylene Diamine Tetraacetic Acid (EDTA), 340 mM NaCl, 30 mM trisodium citrate and pH 7.0) for haemolymph. The haemolymph was thoroughly mixed with cooled (4°C) anticoagulant solution at 1:2 (v/v). Haemolymph samples from shrimp collected two times, those were after 30 days of feeding (before challenged test) and 7 days of challenge test with IMNV and V. harveyi (after challenged test).
The THCs were determined using a haemocytometer at 100 times magnification. Cells were counted on both sides of the grids. THCs were calculated using the following equation:
THC = Cells counted×dilution factor×1000 = Volume of grid (0.1 mm3)
Phenoloxidase activity was determined according to Liu and Chen (2004). Phenoloxidase (PO) activity was estimated spectrophotometrically using L-3, 4 dihydroxyphenylalanine (L-DOPA) and trypsin. Briefly, a total of 50 μL of haemolymph was incubated with 50 μL of 0.1% trypsin in CAC buffer at 25°C for 10 min and then 50 μL of L-DOPA (0.3% in CAC buffer) was added, mixed and optimum density was measured at 490 nm. One unit of enzyme activity was defined as an increase in absorbance of 0.001 min/mg protein.
The Respiratory Burst (RB) activity of haemocytes was measured by reduction of NBT (nitroblue tetrazolium) as a measure of superoxide anion (O2¯) following the method of Song and Hsieh (1994). A total of 300 mL haemolymph-anticoagulant mixture was incubated for 30 min at room temperature, subsequently centrifuged at 3000 rpm for 20 min and the supernatant was discarded. Then 100 mL NBT (HBSS solution with 0.3% NBT) was added and allowed to stand for 2 h at room temperature. Mixture was centrifuged at 3000 rpm for 10 min, the supernatant was removed and added to 100 mL of absolute methanol for further centrifuged at 3000 rpm for 10 min (supernatant discarded). Formed pellets were then rinsed 2 times with 70% methanol. The next 120 mL KOH (2 M) and 140 mL of dimethyl sulfoxide (DMSO) was added to dissolve the pellet. Insoluble pellet was then inserted into the microplate to measure Optical Density (OD) using a microplate reader at a wave length of 630 nm. Respiratory burst expressed as NBT reduction per 10 μL haemolymph.
Statistical analyses: Results are presented as Means±SD (standard deviation of means). Data analysis was done with statistical analysis methods at a 95% confidence interval (α = 0.05). Statistical analyses were performed using a statistic program SPSS (version, 17.0). One-way analysis of variance (One-way ANOVA) was used to determine significant differences between the treatments followed by Duncan’s Multiple Range Test (DMRT).
RESULTS
Growth and feed conversion ratio: Supplementation of synbiotic appear to increase the DGR and decrease the FCR as compared to the controls. The highest DGR (7.45±0.16%) and the lowest FCR (1.14±0.05), respectively observed in treatment B (probiotic 1% + prebiotic 2%) and was significantly different than control groups (p<0.05; Fig. 1). The other treatments had no significant difference in growth performances compared with the control groups (p>0.05).
Immune response: Total Haemocyte Count (THC) of shrimp fed on different doses of synbiotic are shown in Fig. 2. After 30 days of culture, THC increased significantly in the group treated with probiotic 2% + prebiotic 4% (C) compared with control groups. THC of shrimp from the groups fed synbiotic supplemented with combination probiotic 0.5% + prebiotic 1% (A), probiotic 1% + prebiotic 2% (B), probiotic 2% + prebiotic 4% (C) were higher (p<0.05; Fig. 3) compared control (+). Shrimp fed with the control positive diet produced the lowest THC.
Phenoloxidase (PO) activities in haemolymph showed increasing tendency with the increasing doses of synbiotic in diets (Fig. 4). The shrimp in treatment B (probiotic 1% + prebiotic 2%) and C (probiotic 2% + prebiotic 4%) produced the highest PO activities with, followed by shrimp fed with treatment A (probiotic 0.5% + prebiotic 1%) and they were significantly higher than the control groups (Fig. 3; p<0.05). After being given the challenge test with co-infection IMNV and V. harveyi, PO activities in control (-), control (+) and all treatments showed an increase.
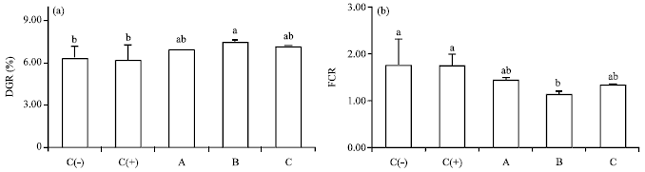 | |
| Fig. 1(a-b): | (a) Daily growth rate and (b) Feed conversion ratio of L. vannamei, Data is expressed as Mean±SD. Means in the same column sharing the same superscript letter are not significantly different determined by Duncan’s test (p>0.05) |
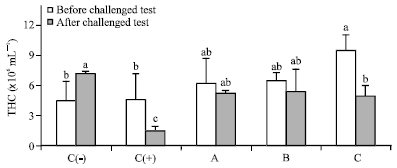 | |
| Fig. 2: | Total haemocyte count of L. vannamei, Data is expressed as Mean±SD. Means in the same column sharing the same superscript letter are not significantly different determined by Duncan’s test (p>0.05) |
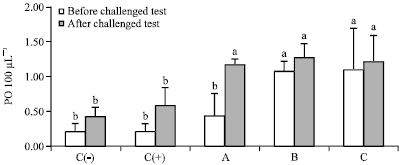 | |
| Fig. 3: | Phenoloxidase activities (PO) of L. vannamei, Data is expressed as Mean±SD. Means in the same column sharing the same superscript letter are not significantly different determined by Duncan’s test (p>0.05) |
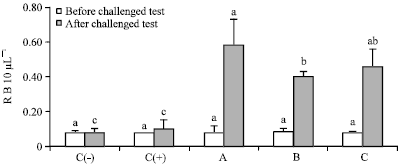 | |
| Fig. 4: | Respiratory burst of L. vannamei, Data is expressed as Mean±SD. Means in the same column sharing the same superscript letter are not significantly different determined by Duncan’s test (p>0.05) |
PO activities were lowest in control (-) and control (+) was significantly different with all treatments diet with synbiotic supplementation. In this study, respiratory burst activities (RB) for all treatments were presented in Fig. 4. After feeding trial for 30 days, RB activity in Haemolymph was not significantly different between synbiotic treatments and controls (Fig. 4; p>0.05). In contrast, RB activity after challenged with co-infection IMNV and V. harveyi significantly increased and was significantly different with controls (Fig. 4; p>0.05).
Survival: Shrimp survival after the 30 days of experimental period was similar in all experimental groups. Shrimp survivals after challenged with co-infection IMNV and V. harveyi significantly increase as the dose of dietary probiotic SKT-b and prebiotic in diet increased (Fig. 5; p<0.05). Shrimp fed with the control positive diet produced the lowest survival and were significantly different (p<0.05) from the groups fed synbiotic supplemented with combination of probiotic 0.5% + prebiotic 1% (A), probiotic 1% + prebiotic 2% (B) and probiotic 2% + prebiotic 4% (C).
Confirmation of IMNV with PCR analysis: Confirmation of the presence of IMNV in the shrimp performed by Polymerase Chain Reaction (PCR) analysis at the Laboratory of Fish Disease and Environment Diagnostic, Serang. PCR test performed on three samples of shrimp that showed clinical symptoms of infection IMNV (samples 1, 2 and 3). PCR test results showed that the samples 1, 2 and 3 were positive with IMNV (Fig. 6).
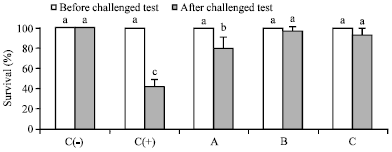 | |
| Fig. 5: | Survival of L. vannamei, Data is expressed as Mean±SD. Means in the same column sharing the same superscript letter are not significantly different determined by Duncan’s test (p>0.05) |
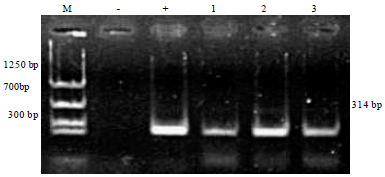 | |
| Fig. 6: | PCR analysis of IMNV infected L. vannamei, agarose gel of amplicons with 3 shrimp samples showing gross signs of white muscle tissue. M, DNA marker, -: Negative control, +: Positive control. The band at 314 bp for sample 4 indicates a light IMNV infection according to the kit instructions |
DISCUSSION
In this present study, combination of probiotic 1% and prebiotic 2% supplemented with the feed to the shrimp culture for 30 days has improved the Daily Growth Rate (DGR) and decreased Feed Conversion Ratio (FCR) of the shrimps. It was indicated that diet B is the optimal dose concentration for supplementing diet of synbiotic. A similar result was found in the European lobster (Homarus gammarus L.) fed a diet containing synbiotic with combination of Bacillus spp. and MOS had significantly increased Weight Gain (WG), SGR and FCR (Daniels et al., 2010). Ai et al. (2011) also showed that the combined use of Bacillus subtilis and fructooligosaccharide (FOS) enhanced growth performance in juvenile large yellow croaker (Larimichthys crocea).
Results suggested that bacterium probiotic strain may participate in digestion processes of shrimp by production exogenous enzymes as well as providing necessary growth factors. Data of counting bacteria in the shrimp intestines showed that the shrimp fed B (probiotic 1% + prebiotic 2%) was the highest total SKT-bR count (1.02×103 CFU g-1) (Table 1). Meanwhile, in the controls of bacteria were not found SKT-bR. In the previous study, bacteria SKT-b bacteria had been found producing the enzyme protease and amylase (Widanarni et al., 2003). Previous study showed that bacteria SKT-b in the shrimp with synbiotic applications increased the activity of digestive enzymes, protease and amylase, in shrimp as compared to control (Lesmanawati, 2013). The increase in digestive enzyme activities would allow the host to digest and absorb more nutrients, thus improve and promote a higher growth and/or feed efficiency (Cerezuela et al., 2011).
| Table 1: | TBC, TVC, SKT-bR Count of Litopenaeus vannamei |
 | |
An improved growth in synbiotic treatment could also related to the prebiotic administration. Previous study showed that prebiotic extracted from sweet potatoes could effectively support the growth of probiotic bacteria (Putra, 2010). Ringo et al. (2010) said that prebiotic can selectively support the growth of specific species of bacteria in the digestive tract of shrimp. Another study reported that, incorporating prebiotic could also increase the length of the intestinal microvilli in L. vannamei (Zhang et al., 2012) and European lobster (Homarus Gammarus L.) (Daniels et al., 2010). The length of the intestinal microvilli may improve the absorption of nutrients which in turn improve the growth performance of the host.
The haemocyte generate inflammatory-type reaction such as phagocytosis, haemocyte clumping, production of reactive oxygen metabolites and the release of microbial proteins (Rodriguez and Le Moullac, 2000; Smith et al., 2003; Hauton, 2012). After challenge test, THC of shrimp in all treatments showed a decline. The decrease of cells hemocytes after challenge test was the effect of the passage of the body’s defense mechanisms such as phagocytosis activity, encapsulation, nodule formation and the degranulation process for system prophenoloxidase activation (ProPO) (Smith et al., 2003).
According to data, PO activity of shrimp before challenge test known to be positively correlated with the value of THC. Hemocytes of shrimp function within the production and release of PO into hemolin in the form of inactive pro-enzyme called ProPO (Smith et al., 2003). After being given a challenge test all treatments showed an increase. Increased activity of PO indicates a high immune response of shrimp. Lesmanawati (2013), stated that an increase in PO activity occurs on the fifth day post-infection IMNV and shrimp on synbiotic treatment resulted in an increase in PO was higher than the control.
Respiratory Burst (RB) is a mechanism for elimination of particles by phagocytic cells which involves the release of degradative enzymes to the phagosome (oxygen-dependent killing mechanism) (Rodriguez and Le Moullac, 2000). The results showed that the activity of RB is not increased by treatment of synbiotic but the increase is due to co-infection of IMNV and V. harveyi. Several studies have shown that the application of synbiotic was not a significant influence on the activity of RB (Ai et al., 2011), gilth head seabream (Cerezuela et al., 2012) while in some other studies, RB activity in synbiotic significantly higher showed in Koi (Cyprinus carpio) (Lin et al., 2012).
Increasing the levels of THC, PO, RB activity on shrimp along with the increasing dose given synbiotic showed that administration of synbiotic in feed can improve the immune system and the health status of the shrimp. Cell wall components of probiotic bacteria, such as β-glucan, lipopolysaccharide, might also contribute to the immunostimulatory effects (Smith et al., 2003; Gullian et al., 2004). Prebiotic in application of synbiotic act like growth factor to particular commensal bacteria which inhibit the adherence and invasion of pathogens in the colonic epithelia by competing for the same glycoconjugates present on the surface of epithelial cells, altering the colonic pH, favoring the barrier function, improving the mucus production, production short-chain fatty acids and inducing cytokine production (Delgado et al., 2011). The Gut-associated Lymphoid Tissue (GALT) is the biggest tissue in the immune system comprising 60% of all lymphocytes in the body (Delgado et al., 2011). The similar effect also confirmed the synergistic effects of synbiotic on the immunostimulation and disease protection in shrimp. Shrimp immune responses increased with synbiotic treatment as shown in some previous studies (Li et al., 2009; Van Hai and Fotedar, 2009; Arisa, 2011; Lesmanawati, 2013). Li et al. (2009) reported that the shrimp fed with synbiotic (PB at dose of 108 CFU g-1 feed in combination with 0.2% IMO) produced the highest RB activities) and was significantly different from the control.
This synbiotic had significantly reduced mortality of the shrimp challenged by co-infection IMNV and V. harveyi and also stimulate immunity of the shrimp. The isolation of probiotic bacterium, SKT-b had been successfully used as probiotic in shrimp. In the previous study, probiotic SKT-b had been reported that synbiotic may enhance resistance of shrimp (L. vannamei) to V. harveyi (Arisa, 2011) and IMNV (Lesmanawati, 2013) with single infection. The result have been reported that SKT-b is known to inhibit the growth of V. harveyi pathogens effectively both in vitro and in vivo on tiger shrimp (Penaeus monodon) larva (Widanarni et al., 2003). The high resistance of survival shrimp might be caused by enhanced of innate immunity.
CONCLUSION
Results of this study showed that probiotic bacterium SKT-b with oligosaccharides from sweet potato (I. batatas L.) in shrimp diets can significantly improve growth and disease resistance by enhancing immunity, as well as presumably modulating microflora in the shrimp's gut. The results showed that the most optimum dose of dietary synbiotic in this study was shrimp SKT-b 1% and prebiotic 2%.
ACKNOWLEDGMENTS
This study was part of the master thesis of the first author financially supported by strategies national research (STRANAS, 2013) Directorate General of Higher Education (DGHE) through Dr. Widanarni.
REFERENCES
- Ai, Q., H. Xu, K. Mai, W. Xu, J. Wang and W. Zhang, 2011. Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture, 317: 155-161.
CrossRefDirect Link - Austin, B. and X.H. Zhang, 2006. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Applied Microbiol., 43: 119-124.
CrossRefDirect Link - Balcazar, J.L., I. de Blas, I. Ruiz-Zarzuela, D. Cunningham, D. Vendrell and J.L. Muzquiz, 2006. The role of probiotics in aquaculture. Vet. Microbiol., 114: 173-186.
CrossRefPubMedDirect Link - Cerezuela, R., J. Meseguer and M.A. Esteban, 2011. Current knowledge in synbiotic use for fish aquaculture: A review. J. Aquacult. Res. Dev.
CrossRefDirect Link - Cerezuela, R., F.A. Guardiola, J. Meseguer and M.A. Esteban, 2012. Increases in immune parameters by inulin and Bacillus subtilis dietary administration to gilthead seabream (Sparus aurata L.) did not correlate with disease resistance to Photobacterium damselae. Fish Shellfish Immunol., 32: 1032-1040.
CrossRefDirect Link - Daniels, C.L., D.L. Merrifield, D.P. Boothroyd, S.J. Davies, J.R. Factor and K.E. Arnold, 2010. Effect of dietary Bacillus spp. and mannan oligosaccharides (MOS) on European lobster (Homarus gammarus L.) larvae growth performance, gut morphology and gut microbiota. Aquaculture, 304: 49-57.
CrossRefDirect Link - Delgado, G.T.C., W.M. da Silva Cunha Tamashiro, M.R. Marostica Jr., Y.M.F. Moreno and G.M. Pastore, 2011. The putative effects of prebiotics as immunomodulatory agents. Food Res. Int., 44: 3167-3173.
CrossRefDirect Link - Gullian, M., F. Thompson and J. Rodriguez, 2004. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture, 233: 1-14.
CrossRefDirect Link - Van Hai, N. and R. Fotedar, 2009. Comparison of the effects of the prebiotics (Bio-Mos® and β-1,3-D-glucan) and the customised probiotics (Pseudomonas synxantha and P. aeruginosa) on the culture of juvenile western king prawns (Penaeus latisulcatus Kishinouye, 1896). Aquaculture, 289: 310-316.
CrossRefDirect Link - Hauton, C., 2012. The scope of the crustacean immune system for disease control. J. Invertebr. Pathol., 110: 251-260.
CrossRefDirect Link - Li, J., B. Tan and K. Mai, 2009. Dietary probiotic Bacillus OJ and isomaltooligosaccharides influence the intestine microbial populations, immune responses and resistance to white spot syndrome virus in shrimp (Litopenaeus vannamei). Aquaculture, 291: 35-40.
CrossRefDirect Link - Liu, C.H. and J.C. Chen, 2004. Effect of ammonia on the immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus. Fish Shellfish Immunol., 16: 321-334.
CrossRefDirect Link - Lin, S., S. Mao, Y. Guan, L. Luo, L. Luo and Y. Pan, 2012. Effects of dietary chitosan oligosaccharides and Bacillus coagulans on the growth, innate immunity and resistance of koi (Cyprinus carpio koi). Aquaculture, 342-343: 36-41.
CrossRefDirect Link - Phuoc, L.H., M. Corteel, N.C. Thanh, H. Nauwynck and M. Pensaert et al., 2009. Effect of dose and challenge routes of Vibrio spp. on co-infection with white spot syndrome virus in Penaeus vannamei. Aquaculture, 290: 61-68.
CrossRefDirect Link - Ringo, E., R.E. Olsen, T.O. Gifstad, R.A. Dalmo, H. Amlund, G.I. Hemre and A.M. Bakke, 2010. Prebiotics in aquaculture: A review. Aquacult. Nutr., 16: 117-136.
CrossRefDirect Link - Rodriguez, J. and G. Le Moullac, 2000. State of the art of immunological tools and health control of penaeid shrimp. Aquaculture, 191: 109-119.
CrossRefDirect Link - Senapin, S., K. Phewsaiya, M. Briggs and T.W. Flegel, 2007. Outbreaks of Infectious Myonecrosis Virus (IMNV) in Indonesia confirmed by genome sequencing and use of an alternative RT-PCR detection method. Aquaculture, 266: 32-38.
CrossRefDirect Link - Smith, V.J., J.H. Brown and C. Hauton, 2003. Immunostimulation in crustaceans: Does it really protect against infection? Fish Shellfish Immunol., 15: 71-90.
CrossRefDirect Link - Song, Y.L. and Y.T. Hsieh, 1994. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation of microbicidal substances: Analysis of reactive oxygen species. Dev. Comp. Immunol., 18: 201-209.
PubMedDirect Link - Soto-Rodriguez, S.A., B. Gomez-Gil, R. Lozano, R. del Rio-Rodriguez, A.L. Dieguez and J.L. Romalde, 2012. Virulence of Vibrio harveyi responsible for the Bright-red syndrome in the pacific white shrimp Litopenaeus vannamei. J. Invertebr. Pathol., 109: 307-317.
CrossRefDirect Link - Verschuere, L., G. Rombaut, P. Sorgeloos and W. Verstraete, 2000. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev., 64: 655-671.
CrossRefPubMedDirect Link - Widanarni, W., A. Suwanto, S. Sukenda and B.W. Lay, 2003. Potency of vibrio isolates for biocontrol of vibriosis in tiger shrimp (Penaeus monodon) larvae. J. Biotropia, 20: 11-23.
Direct Link - Zhang, Q., H. Ma, K. Mai, W. Zhang, Z. Liufu and W. Xu, 2010. Interaction of dietary Bacillus subtilis and fructooligosaccharide on the growth performance, non-specific immunity of sea cucumber, Apostichopus japonicus. Fish Shellfish Immunol., 29: 204-211.
CrossRefDirect Link - Zhang, J., Y. Liu, L. Tian, H. Yang, G. Liang and D. Xu, 2012. Effects of dietary mannan oligosaccharide on growth performance, gut morphology and stress tolerance of juvenile Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol., 33: 1027-1032.
CrossRefPubMedDirect Link