
R.F.M. Traifalgar
Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo 5023, Philippines
V.L. Corre
Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo 5023, Philippines
A.E. Serrano
Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo 5023, Philippines
Journal of Fisheries and Aquatic Science
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 340-354







ABSTRACT
Immunoprophylaxis is widely acknowledged as a potential approach in the control of disease outbreaks in shrimp aquaculture. The present study evaluates the efficacy of dietary immunostimulants such as Fucus vesiculosus fucoidan (FCD), Eucheuma cottonii κ-carrageenan (CAR), heat-killed Vibrio harveyi cells (VHK), Vibrio harveyi lipopolysaccharide (VLP) and yeast β-glucan (BGN) in enhancing the immunological response and the resistance of juvenile Penaeus monodon against Vibrio harveyi infection. Experimental diets were incorporated with the test immunostimulants at 2.0 g kg-1 and a two-week feeding trial was conducted. Following the feeding trial, the experimental shrimp were challenged with pathogenic Vibrio harveyi through intramuscular injection and immunological responses were measured. Results showed significant enhancement of shrimp survival after Vibrio harveyi infection in the FCD (82.2%), BGN (84.4%) and VLP (84.4%) dietary treatments as compared to the control (51.1%), CAR (55.6%) and VHK (42.2%) treatments. The high survival against Vibrio harveyi infection in these treatments is associated with the prominent enhancement of hemocyte phagocytosis (VLP: 118.63±4.42; BGN: 120.01±0.60; FCD: 127.41±1.52) and serum antibacterial activities (VLP: 9.02±0.21; BGN: 9.19±0.012; FCD: 9.61±0.31) as compared to the hemocyte phagocytosis (CAR: 103.10±0.59; VHK: 108.01±2.02; Control: 79.170±1.06) and serum antibacterial activities(CAR: 7.61±0.01; VHK: 7.90±0.16; Control: 5.01±0.09) of the control and the other treatments. These results suggest that dietary supplementation of VLP, BGN and FCD at a dose of 2.0 g kg-1 can be used to boost the immunological response and enhance the resistance of P. monodon against Vibrio harveyi infection.
PDF Abstract XML References Citation
Received: March 17, 2012;
Accepted: October 24, 2012;
Published: January 15, 2013
How to cite this article
R.F.M. Traifalgar, V.L. Corre and A.E. Serrano, 2013. Efficacy of Dietary Immunostimulants to Enhance the Immunological Responses and Vibriosis Resistance of Juvenile Penaeus monodon. Journal of Fisheries and Aquatic Science, 8: 340-354.
DOI: 10.3923/jfas.2013.340.354
URL: https://scialert.net/abstract/?doi=jfas.2013.340.354
DOI: 10.3923/jfas.2013.340.354
URL: https://scialert.net/abstract/?doi=jfas.2013.340.354
INTRODUCTION
At present, outbreaks of microbiological diseases including viral, fungal and bacterial infections that threaten the sustainability and profitability of crustacean aquaculture have become an issue of global concern (Raa et al., 1992; Afsharnasab et al., 2009; Fouzi et al., 2012). Luminous Vibriosis, caused by Vibrio harveyi is a serious disease problem in Penaeus monodon aquaculture. This disease has been implicated in considerable financial losses in shrimp farming activities of several countries (Pizzutto and Hirst, 1995; Robertson et al., 1998; Pasharawipas et al., 2011) including the Philippines (Lavilla-Pitogo et al., 1990). The Philippine isolate of Vibrio harveyi is highly pathogenic, can infect shrimp at various life stages and can cause up to 100% mortality even at bacterial cell densities as low as 102 colony forming units (CFU) mL-1 (Lavilla-Pitogo et al., 1990). Disease symptoms are characterized by lethargy, anorexia, muscle opacity, atrophied hepatopancreatic tubules and luminescence of the host (Jiravanichpaisal et al., 1994; Lavilla-Pitogo et al., 1998).
The use of antibiotics was found to be a possible means of control (Baticados and Paclibare, 1992) but its use was eventually restricted due to the increased risk of developing antibiotic resistant microbial strains and being an environmental hazard (Karunasagar et al., 1996). Consequently, alternative approach including the use of probiotics (Soundarapandian and Sankar, 2008), bacteriophage (Srinivasan et al., 2007) and the use of immunostimulants have been advocated (Anderson, 1992; Raa et al., 1992). To date, a number of substances have been reported as effective immunostimulants for different species of cultured shrimp. Inactivated microbes and their cellular components, seaweed polysaccharides, herbal extract and synthetic chemicals are some of the many substances reported to enhance disease resistance of cultured aquatic animals (Sakai, 1999; John et al., 2011; Balasundaram et al., 2012). In spite of these promising results, reports of substances that can improve resistance of P. monodon against V. harveyi have been relatively few. In addition, the differences in experimental animals (species, stage), purity, dose of immunostimulant and administration methods used in previous studies make selection of an appropriate immunostimulant for P. monodon difficult.
It is evident that different immunostimulants activate different aspects of the immune system and their efficacies vary with different host species, delivery methods and types of pathogens (Sakai, 1999; Smith et al., 2003; Agrawal et al., 2010; Agouz and Anwer, 2011). Hence, the present work was conducted to evaluate the efficacy of dietary administered immunostimulants such as, Eucheuma cottonii κ-carrageenan (CAR), Focus vesiculosus fucoidan (FCD), V. harveyi lipopolysaccharide (VLP), yeast β-glucan (BGN) and heat-killed V. harveyi cells (VHK) to strengthen the immune response of juvenile P. monodon and to enhance its survival against V. harveyi infection. These compounds have been known as potent activators of the non-specific immune response of terrestrial animals, fish and some crustacean species (Fujiki et al., 1997; Alabi et al., 1999; Crosbie and Nowak, 2004; Yuan and Song, 2005; Qin et al., 2007; Cheng et al., 2008; Parmentier et al., 2010) but their efficacy on P. monodon by dietary administration have not been fully evaluated.
Dietary supplementation was used because it has been suggested as the most economical and feasible way of delivery in large-scale culture systems (Siwicki et al., 1994; Sakai, 1999; Singh et al., 2011). Furthermore, in order to understand the aspect of the immune system acted on by these substances, immune indices representing both the humoral and cellular immune responses were also evaluated.
MATERIALS AND METHODS
Immunostimulants: Fucoidan from Fucus vesiculosus, κ-carrageenan from Eucheuma cottonii and yeast β-glucan were obtained from Sigma Chemical Co. (St. Louis, Mo, USA). Pathogenic V. harveyi strain PN9801 isolated from diseased P. monodon was obtained from the bacterial collection of SEAFDEC Aquaculture Department in the Philippines. Heat-killed bacteria were prepared by growing V. harveyi on trypticase soy broth (BBL) supplemented with 2% NaCl for 24 h at 28°C with mild shaking. Bacterial cells were collected by centrifugation at 10, 000 rpm for 20 min, washed three times with phosphate buffered saline (PBS; pH 7.2), heat-killed at 80°C for 30 min, lyophilized and stored at 4°C until use. Sterility was confirmed by inoculating the heat-killed bacteria in thiosulfate citrate bile salt sucrose agar (TCBS, Difco) media, incubated at 28°C for 3 days and no bacterial growth was observed. Crude lipopolysaccharide (LPS) was isolated from log-phase culture of V. harveyi by hot phenol extraction following the procedure described by Schille et al. (1989).
Test diets: The basal diet was prepared following the formulation of Deshimaru et al. (1984), containing 46.7% protein, 8.2% fat, 15.2% ash 12% moisture and has been demonstrated to be nutritionally adequate for the optimal growth of juvenile P. monodon. The basal diet was used as control diet and the test immunostimulants were separately incorporated to the basal diet at 2 g kg-1 to formulate the other five experimental diets. This concentration was based on a previous study showing that immunostimulant supplementation at 2 g kg-1 provided optimal immune enhancing effects in P. monodon (Chang et al., 1999, 2003; Lopez et al., 2003).
Experimental animals and feeding: This study was conducted in 2004 (January to April) at the research facilities of the Institute of Aquaculture, College of Fisheries and Ocean Sciences University of the Philippines in Visayas. Juvenile Penaeus monodon were obtained from a population reared in 10 ton-capacity outdoor tanks and apparently healthy shrimp weighing 6.4±1.2 g were selected, acclimatized in laboratory conditions and fed with control diet for 10 days. Following acclimatization, shrimp were randomly distributed into eighteen 35 L aquaria at a density of 25 shrimp aquarium-1, constituting the six dietary treatments including a control, all in triplicate.
Treatment groups were fed the test diets at 5% of shrimp biomass given twice daily at 08:00 and 16:00 h for a period of two weeks. Removal of wastes and replenishment of 40% rearing water were done daily to maintain optimum water quality. During the feeding trial, seawater temperature ranged from 27-28°C, pH from 7.8-8.2, salinity from 32-340/00 and dissolved oxygen from 6.5-7.1 mg L-1. At the termination of the feeding trial, surviving shrimp on each tank were counted, weighed and growth was calculated as:
Challenge test: After the feeding trial, 45 shrimp from each dietary group were separated, maintained in separate aquaria and used for the challenge test. Individual shrimp were intramuscularly injected with 30 μL of V. harveyi suspension in Phosphate Buffered Saline (PBS) at a dose of 1x106 CFU shrimp. This challenge dose was previously standardized to effect a 50-60% mortality in shrimp fed the control diet. Non-infected shrimp group, injected with PBS were also included. Mortalities were recorded daily for 12 days and the cause of death was verified by re-isolation of the bacteria from the hepatopancreas of moribund shrimp.
Immune assays
Extraction of haemolymph and total hemocyte counting: Haemolymph were collected from the ventral sinus cavity of shrimp using 1 mL tuberculin syringe (26 gauge) containing chilled (4°C) anticoagulant solution (10 mM EDTA-Na2, 45 mM NaCl, 10 mM KCl, 10 mM HEPES, pH 7.3) at a proportion of one part hemolymph to three parts anticoagulant solution. Haemolymph was centrifuged at 3 000 rpm for 10 min at 4°C, supernatant was discarded, hemocyte pellet were rinsed twice and re-suspended in shrimp salt solution (45 mM NaCl, 10 mM KCl, 10 mM HEPES, pH 7.3) (Vargas-Albores et al., 1993). Hemocytes were counted using a hemacytometer and viability was assessed by trypan blue exclusion (Weeks-Perkins et al., 1995). Serum was prepared by collecting haemolymph without anticoagulant, allowed to clot and subjected to a freeze thaw cycle 5 times to induce cell lysis and serum collected by centrifugation at 10 000 rpm for 10 min at 4°C (Sritunyalucksana et al., 1999). Seven shrimp from each treatment were used for the assessment of each immunological parameter. All assays were conducted in triplicate.
Hemocyte phagocytosis activity: Phagocytic activity was measured following the method described previously (Weeks-Perkins et al., 1995). Hemocyte suspension (100 μL) was allowed to adhere on a microscope slide, added with 50 μL of phloxine red-stained yeast cell suspension and incubated in a humidified chamber for 30 min at 25°C. The cells were fixed with 2.5% glutaraldehyde, stained with Wright stain and phagocytic hemocytes were observed using an epifluorescence microscope (Hund Wetzlar, Germany). Phagocytic hemocytes and number of ingested yeast cells were counted from 200 hemocytes and Phagocytic Index (PI) was calculated as described by Itami et al. (1994).
Hemocyte respiratory burst activity (NBT reductase assay): The reduction of nitro blue tetrazolium (NBT) to formazan was quantified as a measure of hemocyte superoxide anion production (Song and Hsieh, 1994). Hemocyte suspension (100 μL) was deposited in a 96-well microtiter plates to promote cell adhesion, washed with Leibovitz L-15 medium (Sigma) and stained with 100 μL of 0.1% Nitro blue tetrazolium (NBT) and 0.01% Phorbol Myristate Acetate (PMA) in Leibovitz L-15 at 25°C for 30 min. The supernatant was removed, hemocytes fixed with 70% methanol and air-dried. The formazan formed was dissolved in 120 μL of 2 M KOH and 140 μL of DMSO and quantified by reading the optical density at 630 nm. Control reactions were conducted without the stimulation of hemocytes with PMA. The ratio of the optical density reading of the stimulated hemocytes to the optical density of the control hemocytes was expressed as relative respiratory burst activity (Campa-Cordova et al., 2000).
Serum antibacterial activity: One hundred microliter of V. harveyi suspension (1x105 CFU mL-1) in PBS was added with equal volume of shrimp serum and incubated with shaking at 28°C for 1 h. Then, 50 μL of the sample samples was removed, serially diluted with sterile saline and a 50 μL aliquot of each dilution was spread on TCBS agar plates. Plates were incubated at 28°C for 24 h and bacterial colonies counted. Bacterial suspension added with PBS instead of serum served as the control. Serum antibacterial activity was calculated as described by Alabi et al. (2000). One unit of serum antibacterial activity was defined as a difference of 0.01 between R (control) and R (treatment), where R represents the ratio of change in bacterial counts from the beginning and end of the reactions and calculated as:
where, A0 and At are the bacterial counts at the beginning and end of the reactions, respectively.
Serum hemolytic activity: Serum hemolytic assays were performed following a modification of the method described by Li et al. (2006). Shrimp serum (100 μL) was added to 900 μL of 3% chicken erythrocytes in saline (0.15 M NaCl), incubated at 25°C for 1 h, centrifuged at 500 rpm for 1 min and the released hemoglobin were collected and optical density was read at 540 nm. Control reactions were conducted using saline instead of serum. The 100% hemolytic reaction was carried out by adding deionized water to erythrocytes instead of saline. The results are expressed as percentage of hemolysis (Guzman et al., 1993).
Statistical analyses: Chi-square test (χ2) was used to compare the protective efficacy of the test substances. Data on growth and immunological responses were analyzed by One way analysis of variance (ANOVA) and Tukey’s honest significant difference test was used to compare differences among treatment means (SPSS 10, SPSS, Inc., USA). All probability values were set at 0.05 level of significance.
RESULTS
Growth: At the end of the two-week feeding trial, all dietary treatments promoted positive growth but were not statistically different. Highest growth value was observed in VLP treatment with 33.50±0.8% followed by VHK with 33.23±1.7%. The control treatment exhibited a 32.97±1.3% growth, CAR with 32.77±1.1%, BGN with 32.14±0.5% and fucoidan treatment has the lowest growth value of 32.06±1.1% (Fig. 1). This indicates that the test immunostimulants were acceptable, not toxic and did not promote stress to the test animals at the concentration tested. Further, no mortalities were observed among the treatments throughout the experimental period.
Challenge test: Protective effects of the test compounds were evaluated by challenge test against V. harveyi and surviving shrimp were recorded until 12 days post-challenge (Table 1).
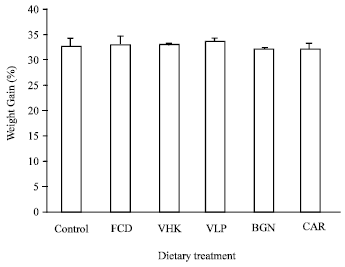 | |
| Fig. 1: | Growth of shrimp fed diets supplemented with the test compounds for 14 days. Value are Mean±SE of three replicate groups per treatment. Control, diet without immunostimulant (32.97±1.3); VHK, heat-killed Vibrio harveyi cells (33.23±1.7); CAR, κ-carrageenan (32.77±1.1); VLP, Crude Vibrio harveyi lipopolysaccharide (33.50±0.8); BGN, β-glucan (32.14±0.5); FCD, fucoidan (32.06±1.1) |
| Table 1: | Survival of shrimp in different treatment groups 12 days after the challenge test with V. harveyi |
 | |
| *Significant difference from the control treatment (χ2 test; p<0.05). Control, diet without immunostimulant; VHK, heat-killed Vibrio harveyi; CAR, κ-carrageenan; VLP, crude Vibrio harveyi lipopolysaccharide; FCD, fucoidan; BGN, β-glucan; Buffer control, shrimps injected with phosphate buffered saline instead of V. harveyi, n = 45 | |
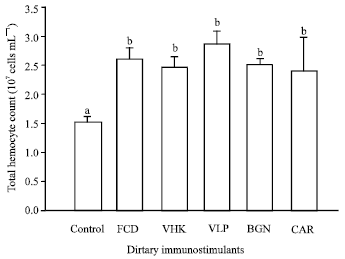 | |
| Fig. 2: | Total hemocyte count of shrimp fed diets supplemented with the test compounds for 14 days. Value are Mean±SE of triplicate assays per shrimp (N = 7 shrimp) on each treatment group. Bars bearing similar superscript letters are not statistically different (p<0.05). Control, diet without immunostimulant (1.50±0.12x107); VHK, heat-killed Vibrio harveyi (2.46±0.19x107); CAR, κ-carrageenan (2.40±0.58x107); VLP, Crude Vibrio harveyi lipopolysaccharide (2.86±0.23x107); BGN, β-glucan (2.50±0.12 x107); FCD, fucoidan (2.60±0.20x107) |
No mortalities were observed in PBS-injected control group. Significantly higher survival values were recorded in BGN (84.4%), FCD (82.2%) and VLP (84.4%) treatments in comparison with CAR (55.6%), VHK (42.2%) and control treatments (51.1%). Percentage survival of shrimp in CAR and VHK treatments were not significantly different from that of the control group. Luminescent colonies of Vibrio harveyi were isolated from the hepatopancreas of deceased shrimp.
Total hemocyte count: Shrimp in treatments receiving the test compounds exhibited a significantly higher total hemocyte counts as compared to the control (Fig. 2). However, no significant differences were observed in the total hemocyte counts of shrimp receiving the different immunostimulants.
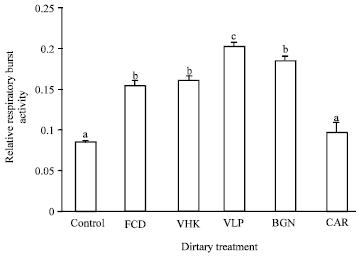 | |
| Fig. 3: | Hemocyte respiratory burst activity of shrimp fed diets supplemented with the test compounds for 14 days. Values are Mean±SE of triplicate assays per shrimp (N= 7 shrimp) on each treatment group. Bars bearing similar superscript letters are not statistically different (p<0.05). Control, diet without immunostimulant (0.08±0.002); VHK, heat-killed Vibrio harveyi (0.16±0.006); CAR, κ-carrageenan (0.10±0.012); VLP, Crude Vibrio harveyi lipopolysaccharide (0.20±0.006); BGN, β-glucan (0.18±0.007); FCD, fucoidan (0.15±0.007) |
Highest value of total hemocyte count was observed in VLP (2.86±0.23x107), followed by FCD (2.60±0.20x107), BGN (2.50±0.12x107), VHK (2.46±0.19x107) and CAR (2.40±0.58x107), respectively. The control treatment (1.50±0.12x107) has the lowest total hemocyte count.
Hemocyte respiratory burst activity: Respiratory burst activity of shrimp in VLP treatment (0.20±0.006) was significantly higher than those of other treatments (Fig. 3). While respiratory burst activities of shrimp in BGN (0.18±0.007), VHK (0.16±0.006) and FCD (0.15±0.007) treatments were not significantly different but were significantly higher than those of CAR (0.10±0.012) and the control treatment (0.08±0.002).
Hemocyte phagocytosis activity (PI): All treatments receiving the test compounds elicited a significant enhancement of the hemocyte phagocytic activity as compared to the control (Fig. 4). Among the substances tested, FCD treatment elicited the highest PI (127.41±1.52) but not significantly different from BGN (120.01±0.60) and VLP (118.63±4.42) treatments. CAR (103.10±0.59) and VHK (108.01±2.02) treatments elicited a moderate response, which were significantly higher than the control but lower compared to FCD, BGN and VLP treatments. The control group (79.170±1.06) showed the lowest phagocytic response.
Serum antibacterial activity: FCD treatment (9.61±0.31) exhibited the highest serum antibacterial activity followed by BGN (9.19±0.012) and VLP (9.02±0.21) treatments. No significant differences in serum antibacterial activity were noted among these treatments.
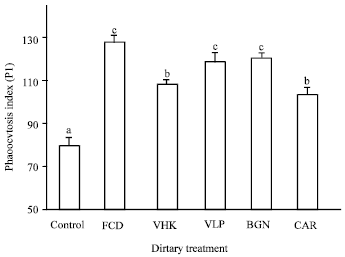 | |
| Fig. 4: | Hemocyte phagocytosis activity of shrimp fed diets supplemented with the test compounds for 14 days. Values are Man±SE of triplicate assays per shrimp (N = 7 shrimp) on each treatment group. Bars bearing similar superscript letters are not statistically different (p<0.05). Control, diet without immunostimulant (79.170±1.06); VHK, heat-killed Vibrio harveyi (108.01±2.02); CAR, κ-carrageenan (103.10±0.59); VLP, Crude Vibrio harveyi lipopolysaccharide (118.63±4.42); BGN, β-glucan (120.01±0.60); FCD, fucoidan (127.41±1.52) |
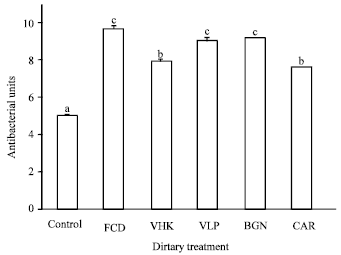 | |
| Fig. 5: | Serum antibacterial activity of shrimp fed diets supplemented with the test compounds for 14 days. Values are Mean±SE of triplicate assays per shrimp (N = 7 shrimp) on each treatment group. Bars bearing similar superscript letters are not statistically different. Control, diet without immunostimulant (5.01±0.09); VHK, heat-killed Vibrio harveyi (7.90±0.16); CAR, κ-carrageenan (7.61±0.01); VLP, Crude Vibrio harveyi lipopolysaccharide (9.02±0.21); BGN, β-glucan (9.19±0.012); FCD, fucoidan (9.61±0.31) |
Groups receiving supplemented CAR (7.61±0.01) and VHK (7.90±0.16) showed significantly higher serum dietary antibacterial activities than groups fed the control diet, but significantly lower compared to those of shrimp fed diets supplemented with FCD, BGN and VLP. The control group (5.01±0.09) showed the lowest serum antibacterial activity (Fig. 5).
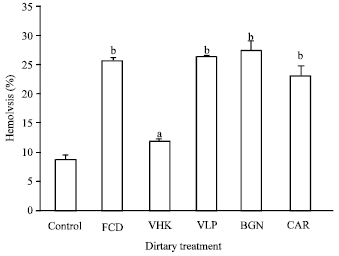 | |
| Fig. 6: | Serum hemolytic activity of shrimp fed diets supplemented with the test compounds for 14 days. Values are Mean±SE of triplicate assays per shrimp (N = 7 shrimp) on each treatment group. Bars bearing similar superscript letters are not statistically different. Control, diet without immunostimulant (8.73_}0.93); VHK, heat-killed Vibrio harveyi (11.68±0.59); CAR, κ-carrageenan (11.68±0.59); VLP, Crude Vibrio harveyi lipopolysaccharide (25.98±0.17); BGN, β-glucan (27.23±1.52); FCD, fucoidan (25.50±0.55) |
Serum hemolytic activity: Hemolytic activity was the highest in shrimp fed BGN (27.23±1.52) supplemented diets but not significantly different from those fed with FCD (25.50±0.55), CAR (11.68±0.59) and LPS (25.98±0.17) supplemented diets. The control (8.73±0.93) and VHK (11.68±0.59) groups exhibited significantly lower levels of serum hemolytic activity (Fig. 6).
DISCUSSION
The efficacy of dietary supplemented BGN, FCD, VLP, VHK and CAR to promote resistance against V. harveyi infection in P. monodon were evaluated in this study. Among these substances, BGN, FCD and VLP were found effective in promoting a protective immune response as measured in terms of percentage survival following a challenge test. CAR and VHK treatments exhibited lower protective effects that were comparable with the control treatment. In this study, all treatment groups exhibited a statistically similar growth response and no mortalities occurred throughout the experimental period. This suggests that the immunostimulants were acceptable and probably not inhibitory to the overall metabolism of the test animals at the given dose and duration of the experiment.
The enhanced protection conferred by BGN is associated with the enhancement of both the humoral and cellular immune responses. Relative to the control, hemocyte count, hemolytic, phagocytosis, respiratory burst and serum antibacterial activities were significantly elevated in BGN treatment, indicating the importance of these responses in resistance against V. harveyi infection. The present results corroborate with previous in- vitro and in-vivo findings, elucidating the potency of BGN in activating crustacean humoral and cellular immune responses (Sritunyalucksana et al., 1999; Vitvicka and Sima, 2004). In-vitro incubation of crustacean hemocyte with soluble β-glucan resulted to increased hemocyte phagocyte activity (Smith and Soderhall, 1983) and increased hemocyte respiratory burst activity of P. vannamei (Campa-Cordova et al., 2000). Also in P. monodon β-glucan treatment was reported to activate the production of hemolymph antibacterial proteins (Sritunyalucksana et al., 1999). Moreover, similar to the present findings Sung et al. (1994) reported enhanced survival of P. monodon post-larvae immersed in β-glucan and challenged with Vibrio vulnificus. Immersion with β-glucan was also documented to increase survival of Macrobrachium rosenbergii post larvae exposed to pathogenic Vibrio alginolyticus (Misra et al., 2004) Enhanced protection against vibrio strain NU1 infection was also documented in P. japonicus fed with diets supplemented with β-glucan (Itami et al., 1994). Additionally, dietary administration of β-glucan in juvenile P. monodon promoted increased resistance and improves survival against white spot syndrome virus infection (Chang et al., 2003). All of these earlier findings indicate that the improvement of shrimp survival against microbial infection due to β-glucan treatment could be attributed to the enhancement of elevated immune responses similar to that observed in the present study.
Lipopolysaccharide (LPS) constitutes a part of the cell membrane of gram-negative bacteria and has been used as an important tool in defining vertebrate and crustacean immune functions (Sritunyalucksana et al., 2002; Van de Braak et al., 2002; Pakzad et al., 2010). In spite of this, studies regarding the practical application of LPS as an immunostimulant for shrimp have been limited. In the present work, dietary supplementation of VLP resulted in higher protection against V. harveyi infection due to the up-regulation of immune responses. These results are consistent with earlier reports elucidating the role of bacterial LPS in immune activation. Injection of LPS has been shown to promote proliferation of hemocytes in P. monodon (Van de Braak et al., 2002) and in M. japonicus (Sequira et al., 1996), up-regulates gene transcription of antimicrobial peptides in P. vannamei (Okumura, 2007) and in-vitro incubation with hemocytes activated prophenol oxidase cascade (Cardenas et al., 2004). Similar to the present findings, enhancement of hemocyte phagocytic activity and activation of hemolymph virus neutralizing factor resulting in improved survival against penaeid acute viremia virus was also demonstrated in M. japonicus fed diets supplemented with Pantoea agglomerans LPS (Takahashi et al., 2000). Furthermore, it has been documented that the mechanism by which LPS promotes shrimp resistance against microbial infection is due to the activation of an anti-LPS peptide (Nagoshi et al., 2006). This peptide has been characterized as having, a strong affinity to bacterial LPS, broad spectrum antibacterial, anti-fungal and antiviral activities (Somboonwiwat et al., 2005; Liu et al., 2006; De la Vega et al., 2008). Activation of this peptide could probably be involved in the enhancement of P. monodon resistance against V. harveyi infection as observed in the present study.
Fucoidan is an acid-soluble polysaccharide of sulfated L-fucose mainly produced by brown algae (Eluvakkal et al., 2010). This compound has been known to be a potent activator of phagocytosis and inducer of cytokine secretion in vertebrate immune cells (Heinzelmann et al., 1998) but information on its effects on shrimp has been scarce. The present findings show that dietary fucoidan supplementation significantly elevated phagocytosis and serum antibacterial activities resulting to increased survival of P. monodon juveniles against V. harveyi infection and is the first time to be documented in shrimp. The enhanced resistance seen in FCD treatment could be attributed to the significant enhancement of immunological responses, specifically the hemocyte phagocytic and serum antibacterial activities. Although, detailed studies on the effects of fucoidan on crustacean immune functions are limited, but its effects on vertebrate immune system in promoting phagocytosis and release of cytokines are well documented (Heinzelmann et al., 1998). Similar effect has been documented in P. monodon wherein fucoidan injection induced the synthesis of a phagocytosis activating protein, known to activate hemocyte phagocytic response that eliminates invading viral or microbial pathogens (Deachamag et al., 2006). Dietary supplemented fucoidan has also been shown to increase the resistance of M. japonicus (Takahashi et al., 1998) and P. monodon (Chotigeat et al., 2004) against white spot syndrome virus infection. In these reports, significant enhancement of hemocyte phagocytic activity were also observed, suggesting a strong functionality of fucoidan towards the activation of cellular immune response in a manner similar to that observed in the present study and in the activation of macrophages in vertebrate organisms.
The low percentage survival observed in VHK and CAR treatments was associated with lower immunological responses in comparison with BGN, FCD and VLP supplementation. This suggests that the magnitude of immunological responses elicited by CAR and VHK was inadequate to render an effective protection against V. harveyi. The low immunological responses observed in these treatments could have been caused by the digestive degradation of the functional components of these substances, rendering it unrecognizable by the immune system. Decrease in vaccine efficacy due to digestive degradation has also been documented in fish (Jenkins et al., 1994). Similar results indicating the ineffectiveness of Vibrio bacterin supplementation in eliciting a protective immune response were also reported in Penaeus indicus larvae (Alabi et al., 1999). Also, the present findings is in agreement with the results of Goarant and Boglio (2000) in Litopenaeus stylirostris fed diets supplemented with Vibrio bacterin that did not improve the immunological responses of this shrimp. In contrast to the present findings, Heidarieh et al. (2010) reported enhanced survival of P. vannamei post larvae challenged with V. harveyi. The larvae were maintained with diets supplemented with a bacterin consisting of a mixture of different Vibrio species (Vibriomax) and an algin based immunostimulant (Ergosan). The combination of mixed Vibrio bacterin and alginate immunostimulant may account for the difference in the infection survival response observed in that study.
Contrary to the present findings, carrageenan administered by intramuscular injection has been reported to enhance the non-specific immune response and increase disease resistance of Cyprinus carpio (Fujiki et al., 1997) and Epinephelus coioides (Cheng et al., 2008). However, the differences in immune physiology between shrimp and fishes and the different route of administration may account for these contrasting results.
Overall, the present results suggest that dietary supplementation of BGN, FCD and VLP at 2 g kg-1 diet could enhance the immunological responses and improve resistance of juvenile P. monodon against V. harveyi. The potential use of these substances for practical application in P. monodon is promising but concerns regarding their effects on shrimp metabolism and growth on a long-term administration await further confirmations.
ACKNOWLEDGMENT
The authors gratefully acknowledge the financial support and assistance of the Department of Agriculture-Bureau of Agricultural Research, Philippines for the completion of this work.
REFERENCES
- Afsharnasab, M., R. Mortezaei, V. Yegane and B. Kazemi, 2009. Gross sign, histopathology and polymerase chain reaction observations of white spot syndrome virus in shrimp specific pathogen free Litopeneaus vannamei in Iran. Asian J. Anim. Vet. Adv., 4: 297-305.
CrossRefDirect Link - Agouz, H.M. and W. Anwer, 2011. Effect of biogen® and Myco-ad® on the growth performance of common carp (Cyprinus carpio) fed a mycotoxin contaminated aquafeed. J. Fish. Aquat. Sci., 6: 334-345.
CrossRefDirect Link - Agrawal, S., S. Khadase and G. Talele, 2010. Bioactive immunomodulatory fraction from Tridax procumbens. Asian J. Biol. Sci., 3: 120-127.
CrossRefDirect Link - Alabi, A.O., J.W. Latchford and D.A. Jones, 2000. Demonstration of residual antibacterial activity in plasma of vaccinated Penaeus vannamei. Aquaculture, 187: 15-34.
CrossRefDirect Link - Alabi, A.O., D.A. Jones and J.W. Latchford, 1999. The efficacy of immersion as opposed to oral vaccination of Penaeus indicus larvae against Vibrio harveyi. Aquaculture, 178: 1-11.
CrossRefDirect Link - Anderson, D.P., 1992. Immunostimulants, adjuvants and vaccine carriers in fish: Applications to aquaculture. Ann. Rev. Fish Dis., 2: 281-307.
CrossRefDirect Link - Balasundaram, A., P.R. Kumari, A. Stalin, V. Masilamani and G. John, 2012. Effect of a commercial probiotic and Cassia auriculata leaf powder on vibriosis induced freshwater prawn, macrobrachium rosenbergii. Asian J. Anim. Vet. Adv., 7: 542-555.
CrossRefDirect Link - Cardenas, W., J.R. Dankert and J.A. Jenkins, 2004. Flow cytometric analysis of crayfish haemocytes activated by lipopolysaccharides. Fish. Shellfish Immunol., 17: 223-233.
CrossRefDirect Link - Chang, C.F., M.S. Su, H.Y. Chen and I.C. Liao, 2003. Dietary β-1,3-glucan effectively improves immunity and survival of Penaeus monodon challenged with white spot syndrome virus. Fish Shellfish Immunol., 15: 297-310.
CrossRefPubMedDirect Link - Cheng, A.C., Y.Y. Chen and J.C. Chen, 2008. Dietary administration of sodium alginate and kappa-carrageenan enhances the innate immune response of brown-marbled grouper Epinephelus fuscoguttatus and its resistance against Vibrio alginolyticus. Vet. Immunol. Immunopathol., 121: 206-215.
PubMed - Chotigeat, W., S. Tongsupa, K. Supamataya and A. Phongdara, 2004. Effect of fucoidan on disease resistance of black tiger shrimp. Aquaculture, 233: 23-30.
CrossRef - Campa-Cordova, A.I., N.Y. Hernandez-Saavedra, R. de Philippis and F. Ascencio, 2000. Generation of superoxide anion and SOD activity in haemocytes and muscle of American white shrimp (Litopenaeus vannamei) as a response to β-glucans and sulphated polysaccharide. Fish Shellfish Immunol., 12: 353-366.
PubMed - Crosbie, P.B.B. and B.F. Nowak, 2004. Immune responses of barramundi, Lates calcarifer (Bloch), after administration of an experimental Vibrio harveyi bacterin by intraperitoneal injection, anal intubation and immersion. J. Fish Dis., 27: 623-632.
Direct Link - Deachamag, P., U. Intaraphad, A. Phongdara and W. Chotigeat, 2006. Expression of a Phagocytosis Activating Protein (PAP) gene in immunized black tiger shrimp. Aquaculture, 255: 165-172.
CrossRefDirect Link - De la Vega, E., N.A. O'Leary, J.E. Shockey, J. Robalino and C. Payne et al., 2008. Anti-lipopolysaccharide factor in Litopenaeus vannamei (LvALF): A broad spectrum antimicrobial peptide essential for shrimp immunity against bacterial and fungal infection. Mol. Immunol., 45: 1916-1925.
CrossRefPubMedDirect Link - Deshimaru, O., K. Kuroki and M.A. Mazid, 1984. Nutritional quality of compounded diets for prawn Penaeus monodon. Bull. Jpn. Soc. Sci. Fish., 51: 1037-1044.
Direct Link - Eluvakkal, T., S.R. Sivakumar and K. Arunkumar, 2010. Fucoidan in some Indian brown seaweeds found along the coast gulf of mannar. Int. J. Bot., 6: 176-181.
CrossRefDirect Link - Fouzi, M.N.M., M. Shariff and F.M. Yusoff, 2012. Stress quantification in Penaeus monodon exposed to different levels of ammonia and subsequent infection to WSSV. Res. J. Vet. Sci., 5: 12-24.
CrossRefDirect Link - Goarant, C. and E. Boglio, 2000. Changes in hemocyte counts in Litopenaeus stylirostris subjected to sublethal infection and to vaccination. J. World Aquacult. Soc., 31: 123-129.
CrossRefDirect Link - Guzman, M.A., J.L. Ochoa and F. Vargas-Albores, 1993. Hemolytic activity in the brown shrimp (Penaeus californiensis Holmes) haemolymph. Comp. Biochem. Physiol. A: Physiol., 106: 271-275.
CrossRefDirect Link - Heidarieh, M., M. Afsharnasab, M. Soltani, A. Dashtyannasab, S. Rajabifar, N. Sheikhzadeh and A.H. Tamimi, 2010. Effects of ergosan and vibromax to prevent vibriosis and WSSV in Litopeaneus vannamei. J. Fish. Aquat. Sci., 5: 120-125.
CrossRefDirect Link - Heinzelmann, M., H.C. Polk Jr. and F.N. Miller, 1998. Modulation of lipopolysaccharide-induced monocyte activation by heparin-binding protein and fucoidan. Infect. Immunity, 66: 5842-5847.
Direct Link - Itami, T., Y. Takahashi, E. Tsuchihira, H. Igusa and M. Kondo, 1994. Enhancement of disease resistance of kuruma prawn Penaeus japonicus and increase in phagocytic activity of prawn hemocytes after oral administration of β1,3-glucan (Schizophyllan). Proceedings of the 3rd Asian Fisheries Forum, October 26-30, 1992, Asian Fisheries Society, Manila, Philippines, pp: 375-378.
- Jenkins, P.G., A.B. Wrathmell, J.E. Harris and A.L. Pulsford, 1994. Systemic and mucosal immune responses to enterically delivered antigen in Oreochromis mossambicus. Fish Shellfish Immunol., 4: 255-271.
CrossRefDirect Link - Jiravanichpaisal, P., T. Miyazaki and C. Limsuwan, 1994. Histopathology, biochemistry and pathogenicity of Vibrio harveyi infecting black tiger prawn Penaeus mondon. J. Aquat. Ann. Health, 6: 27-35.
CrossRefDirect Link - John, G., P.R. Kumari and A. Balasundaram, 2011. Health promoting biochemical effects of three medicinal plants on normal and Aeromonas hydrophila infected Labeo rohita. J. Fish. Aquat. Sci., 6: 633-641.
CrossRef - Karunasagar, I., S.K. Otta and I. Karunasagar, 1996. Biofilm formation by Vibrio harveyi on surfaces. Aquaculture, 140: 241-245.
CrossRef - Lavilla-Pitogo, C.R., E.M. Leano and M.G. Paner, 1998. Mortalities of pond-cultured juvenile shrimp, Penaeus monodon, associated with dominance of luminescent vibrios in the rearing environment. Aquaculture, 164: 337-349.
CrossRefDirect Link - Lavilla-Pitogo, C.R., M.C. L. Baticados, E.R. Cruz-Lacierda and L.D. de la Pena, 1990. Occurrence of luminous bacterial disease of Penaeus monodon larvae in the Philippines. Aquaculture, 91: 1-13.
CrossRef - Li, Y.Q., J. Li and Q.Y. Wang, 2006. The effects of dissolved oxygen concentration and stocking density on growth and non-specific immunity factors in Chinese shrimp, Fenneropenaeus chinensis. Aquaculture, 256: 608-616.
CrossRef - Liu, H., P. Jiravanichpaisal, I. Soderhall, L. Cerenius and K. Soderhall, 2006. Antilipopolysaccharide factor interferes with white spot syndrome virus replication in vitro and in vivo in the crayfish Pacifastacus leniusculus. J. Virol., 80: 10365-10371.
Direct Link - Lopez, N., G. Cuzon, G. Gaxiola, G. Taboada and M. Valenzuela et al., 2003. Physiological, nutritional and immunologicalrole of dietary β 1-3 glucan and ascorbic acid 2-monophosphate in Litopenaeus vannamei juveniles. Aquaculture, 224: 223-224.
CrossRefDirect Link - Misra, C.K., B.K. Das, J. Pradhan, P. Pattnaik, S. Sethi and S.C. Mukherjee, 2004. Changes in lysosomal enzyme activity and protection against Vibrio infection in Macrobrachium rosenbergii (De Man) post larvae after bath immunostimulation with β-glucan. Fish Shellfish Immunol., 17: 389-395.
CrossRefPubMedDirect Link - Nagoshi, H., H. Inagawa, K. Morii, H. Harada and C. Kohchi et al., 2006. Cloning and characterization of a LPS-regulatory gene having an LPS binding domain in kuruma prawn Marsupenaeus japonicus. Mol. Immunol., 43: 2061-2069.
CrossRefDirect Link - Okumura, T., 2007. Effects of lipopolysaccharide on gene expression of antimicrobial peptides (penaeidins and crustin), serine proteinase and prophenoloxidase in haemocytes of the Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol., 22: 68-76.
CrossRefPubMedDirect Link - Pakzad, I., A. Rezaee, M.J. Rasaee, A.Z. Hosseini and B. Tabbaraee et al., 2010. Novel approach of vaccination against Brucella abortus 544 based on a combination of fusion proteins, human serum albumin and Brucella abortus lipopolysaccharides. J. Biol. Sci., 10: 767-772.
CrossRefDirect Link - Parmentier, H.K., G. de Vries Reilingh, P. Freke, R.E. Koopmanschap and A. Lammers, 2010. Immunological and physiological differences between layer- and broiler chickens after concurrent intratracheal administration of lipopolysaccharide and human serum albumin. Int. J. Poult. Sci., 9: 574-583.
CrossRefDirect Link - Pasharawipas, T., J. Manopvisetcharean and T.W. Flegel, 2011. Phage treatment of Vibrio harveyi: A general concept of protection against bacterial infection. Res. J. Microbiol., 6: 560-567.
CrossRef - Qin, Y., J. Wang, S. Wang and Q. Yan, 2007. Antigenicity analysis of Vibrio harveyi TS-628 strain. Front. Biol. China, 2: 263-267.
CrossRefDirect Link - Robertson, P.A.W., J. Calderon, L. Carrera, J.R. Stark, M. Zherdmant and B. Austin, 1998. Experimental Vibrio harveyi infections in Penaeus vannamei larvae. Dis. Aquat. Organ., 32: 151-155.
Direct Link - Sakai, M., 1999. Current research status of fish immunostimulants. Aquaculture, 172: 63-92.
CrossRefDirect Link - Sequira, T., D. Tavares and M. Arala-Chaves, 1996. Evidence for circulating hemocyte proliferation in the shrimp Penaeus japonicus. Dev. Comp. Immunol., 20: 97-104.
CrossRefDirect Link - Singh, G.P., A.K. Sinha, K.N. Madhusudhan, P.K. Kumar, B.C. Prasad, D.K. Roy and A. Sahay, 2011. Attenuation of Antheraea mylitta Cytoplasmic Polyhedrosis Virus (AmCPV) and its potential as an oral vaccine against virus diseases in tasar silkworm, Antheraea mylitta D. Int. J. Virol., 7: 33-38.
CrossRef - Siwicki, A.K., D.P. Anderson and G.L. Rumsey, 1994. Dietary intake of immunostimulants by rainbow trout affects non-specific immunity and protection against furunculosis. Vet. Immunol. Immunopathol., 41: 125-139.
PubMedDirect Link - Smith, V.J., J.H. Brown and C. Hauton, 2003. Immunostimulation in crustaceans: Does it really protect against infection? Fish Shellfish Immunol., 15: 71-90.
CrossRefDirect Link - Smith, V.J. and K. Soderhall, 1983. β1, 3 glucan activation of crustacean hemocytes in vitro and in vivo. Biol. Bull., 164: 299-314.
Direct Link - Somboonwiwat, K., M. Marcos, A. Tassanakajon, S. Klinbunga and A. Aumelas et al., 2005. Recombinant expression and anti-microbial activity of Anti-Lipopolysaccharide Factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol., 29: 841-851.
CrossRefPubMedDirect Link - Song, Y.L. and Y.T. Hsieh, 1994. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation of microbicidal substances: Analysis of reactive oxygen species. Dev. Comp. Immunol., 18: 201-209.
PubMedDirect Link - Soundarapandian, P. and S. Sankar, 2008. Effect of probiotics on the survival and production of black tiger shrimp Penaeus monodon (Fabricius). Int. J. Zool. Res., 4: 35-41.
CrossRefDirect Link - Srinivasan, P., P. Ramasamy, G.P. Brennan and R.E.B. Hanna, 2007. Inhibitory effects of bacteriophages on the growth of Vibrio sp., pathogens of shrimp in the Indian aquaculture environment. Asian J. Anim. Vet. Adv., 2: 166-183.
CrossRefDirect Link - Sritunyalucksana, K., P. Sithisarn, B. Withayachumnarnkul and T.W. Flegel, 1999. Activation of prophenoloxidase, agglutinin and antibacterial activity in haemolymph of the black tiger prawn, Penaeus monodon, by immunostimulants. Fish Shellfish Immunol., 9: 21-30.
CrossRefDirect Link - Sritunyalucksana, K., S.Y. Lee and K. Soderhall, 2002. A β1, 3-glucan binding protein from black tiger shrimp, Penaeus monodon. Dev. Comp. Immunol., 26: 237-245.
CrossRefDirect Link - Takahashi Y., K. Uehara, R. Watanabe, T. Okumura and T. Yamashita et al., 1998. Efficacy of Oral Administration of Fucoidan a Sulfated Polysaccharide in Controlling White Spot Syndrome in Kuruma Shrimp in Japan. In: Advances in Shrimp Biotechnology, Flegel, T.W. (Ed.). National Center for Genetic Engineering and Biotechnology, Bangkok, Thailand, pp: 171-173.
- Takahashi, Y., M. Kondo, T. Itami, T. Honda and H. Inagawa et al., 2000. Enhancement of disease resistance against penaeid acute viraemia and induction of virus-inactivating activity in haemolymph of kuruma shrimp, Penaeus japonicus, by oral administration of Pantoea agglomerans lipopolysaccharide (LPS). Fish. Shellfish. Immunol., 10: 555-558.
CrossRefPubMedDirect Link - Van de Braak, C.B.T., M.H.A. Botterblom, W. Liu, N. Taverne, W.P.W. Van der Knaap and J.H.W.M. Rombout, 2002. The role of the haematopoietic tissue in haemocyte production and maturation in the black tiger shrimp (Penaeus monodon). Fish Shellfish Immunol., 12: 253-272.
CrossRefDirect Link - Vargas-Albores, F., M.A. Guzman-Murillo and J.L. Ochoa, 1993. An anticoagulant solution for haemolymph collection and prophenoloxidase studies of penaeid shrimp (Penaeus californiensis). Comp. Biochem. Physiol. Part A: Physiol., 106: 299-303.
CrossRefDirect Link - Vitvicka, V. and P. Sima, 2004. β-glucan in invertebrates. Invert. Survival J., 1: 60-65.
Direct Link - Weeks-Perkins, B.A., N. Chansue and D. Wong-Verele, 1995. Assays of Immune Function in Shrimp Phagocytes: Techniques used as Indicators of Pesticide Exposure. In: Techniques in Fish Immunology 4: Immunology and Pathology of Aquatic Invertebrates, Stolen, J.S., T.C. Fletcher, S.A. Smith, J.T. Zelikoff and S.L. Kaatari et al. (Eds.). SOS Publications, Fair Haven, NJ., USA., pp: 173-179.
- Yuan, H. and J. Song, 2005. Preparation, structural characterization and in vitro antitumor activity of kappa-carrageenan oligosaccharide fraction from Kappaphycus striatum. J. Applied Phycol., 17: 7-13.
CrossRefDirect Link