
R. Sanchez-Cardenas
Centro Interdisciplinario de Ciencias Marinas del IPN, Apartado Postal 592, La Paz, Baja California Sur, Mexico
F. Arreguin-Sanchez
Centro Interdisciplinario de Ciencias Marinas del IPN, Apartado Postal 592, La Paz, Baja California Sur, Mexico
Journal of Fisheries and Aquatic Science
Year: 2012 | Volume: 7 | Issue: 6 | Page No.: 379-391







ABSTRACT
This study contributes to the existing knowledge of temporal reproduction patterns in tropical fish species which is potentially relevant to regional fisheries management. The latitudinal variation in Spawning Season Length (SSL) and its relationship with spawning season temperature (SPST) was explored, as well as the latitudinal variation in Spawning Aggregation Duration (SAD), through Spearman’s rank coefficient. Additionally, the lunar synchrony of spawning and the time of day at which spawning occurs were described. We found variable trends for SSL with respect to latitude in the tropics; most patterns did not satisfactorily explain the variation in SSL. The minimum SPST consistently tended to be lower for longer spawning seasons, both within each species and for the entire dataset. We observed SSLs of one to six months across the entire range of latitudes (-30° to 35°) but most spawning seasons longer than six months were observed at latitudes lower than -4° or higher than 10°, where lower water temperatures can be found. The latitudinal tendencies of SAD were variable and poorly supported, but the data on Epinephelus striatus hinted that SSL may vary spatially. Furthermore, the studied species most likely follow lunar and semilunar spawning cycles which are likely to be related to specific daily spawning patterns.
PDF Abstract XML References Citation
Received: February 28, 2012;
Accepted: April 19, 2012;
Published: August 06, 2012
How to cite this article
R. Sanchez-Cardenas and F. Arreguin-Sanchez, 2012. Latitudinal Exploration of the Temporalities of Spawning for Some Tropical Fish Species (Epinephelidae: Plectropomus spp., Mycteroperca spp. and Epinephelus spp.). Journal of Fisheries and Aquatic Science, 7: 379-391.
DOI: 10.3923/jfas.2012.379.391
URL: https://scialert.net/abstract/?doi=jfas.2012.379.391
DOI: 10.3923/jfas.2012.379.391
URL: https://scialert.net/abstract/?doi=jfas.2012.379.391
INTRODUCTION
Reproduction of fish is a relevant issue regarding fish management and conservation and its knowledge is of importance for sustainable exploitation and conservation. Studies has been focused on different ways, general aspect on reproductive biology (Ramadan and El-Halfawy, 2007; Sivashanthini et al., 2008; Chelemal et al., 2009; Kerdgari et al., 2009; Sadeghi et al., 2009), metabolism (Bouriga et al., 2011; Sutharshiny and Sivashanthini, 2011) and physiology related to gonadal maturation (Khalil et al., 2007), or reproductive behaviour (Hosseini et al., 2009; Salcedo-Bojorquez and Arreguin-Sanchez, 2011), among others. However, of particular interest are the spawning aggregations where fishes are more vulnerable to fishing.
The spawning patterns of tropical fishes can occur on daily, lunar and seasonal time scales (Colin, 2012) and their interspecific and intraspecific geographic variations can be complex (Robertson, 1991). There are many tropical marine environments where fish breeding occurs and the temporality of breeding may be associated with environmental factors, the quality and availability of food, water chemistry and day length, among other variables. However, the processes that influence optimal breeding conditions are still poorly understood for most tropical fishes (Johannes, 1978; Lowe-McConnell, 1987; Wootton, 1998). Thus, the geographical variability of reproduction timing is a complex issue. Some tropical fishes exhibit longer spawning seasons nearer to the equator (Johannes, 1978; Craig, 1998) but such behaviour has not been explored for many species (Craig, 1998). Some questions that concern managers remain unanswered; e.g., what is the effect of fishing on those fish resources that form spawning aggregations and sustain brief spawning seasons at low latitudes? The timing of the spawning season for some epinephelid groupers is related to temperature and tends to occur during the summer in cooler latitudes and when temperatures are below the annual maximum in warmer, lower latitudes (Thresher, 1984; Samoilys, 1997). For example, there is evidence that the latitudinal shifts of Epinephelus striatus, whose spawning is restricted to a narrow range of temperatures, may be related to water temperatures (Colin, 1992) but other species may not respond in this way (Colin, 2012). The interest in this type of knowledge relates to the usage of spawning aggregations by fishermen and the need to regulate such exploitation to sustain fisheries. In this contribution, we explored the latitudinal variation of spawning season length and spawning aggregation duration and their relationships with water temperatures for fish species that form spawning aggregations in the Atlantic and Indo-Pacific Oceans. This study integrates information to find patterns that are useful for understanding the temporality of reproduction in tropical areas for regional fisheries management.
MATERIALS AND METHODS
Information on the temporality of grouper (Epinephelidae) reproduction, along with the geographical locations of temporal data, was collected from published scientific documents and FishBase (Froese and Pauly, 2009; www.fishbase.org). Google Earth version 5 was used for georeferencing locations whose geographical coordinates were not specified in the literature. The biological variables considered were spawning season length (SSL, in months), spawning aggregation duration (SAD, in days), the lunar phase during spawning, the time of day at which spawning occurred and the water temperature during the spawning season (minimum, maximum and average). The monthly temperatures during the spawning season were obtained from the POET database (http://poet.jpl.nasa.gov/) generated by the AVHRR Pathfinder algorithm version 5 using measurements with a resolution of 4 km of made by the AVHRR (Advanced Very-High-Resolution Radiometer) NOAA sensor.
The statistical relationships of SSL and SAD with latitude and the relationship of temperature with SSL were explored using Spearman’s rank correlation coefficient (Spearman R) by species and hemisphere (northern and southern). The aim of the analyses was to determine the latitudinal pattern in the available information on reproductive temporality. The Spearman R was used because the data in each analysis set was scarce and non-normal (Zar, 2010). The coefficient ranges from 1 for a perfect positive correlation to -1 for a perfect inverse correlation, with 0 indicating complete independence (Spearman, 1904). The Spearman R was evaluated using Student’s t test with n (number of data) and n-2 degrees of freedom with a significance threshold of p<0.05. Statistica 8.0 software (StatSoft Inc., USA) was used for statistical procedures. Finally, spawning temporality, the lunar synchrony of spawning events and the time of day of spawning were described.
RESULTS
Spawning season length: The Spawning Season Length (SSL) was correlated with latitude and spawning season temperature (SPST) for two Plectropomus spp., four Mycteroperca spp. and eight Epinephelus spp. (Table 1, Fig. 1) from areas of the southeast United States, the Gulf of Mexico, the Mexican Caribbean, the Bahamas, Cuba, the Cayman Islands, the Virgin Islands, Puerto Rico, Jamaica, Belize, the Netherlands Antilles, Colombia, Ascension Island and Brazil in the Atlantic Ocean and also Japan, the Marshall Islands, the Gulf of Mannar, the Federated States of Micronesia, Malaysia, Indonesia, Kenya, Seychelles, the Solomon Islands, the Torres Strait Islands, the Great Barrier Reef, Australia, Fiji, French Polynesia, New Caledonia and the Cook Islands in the Indo-Pacific Ocean.
The relationship between SSL data and latitude showed a patchy distribution between 10° Lat and 35° Lat for the species from the Atlantic Ocean (M. phenax, M. bonaci, M. venenosa, M. tigris, E. itajara, E. guttatus, E. striatus and E. adscensionis) and for one of the species from the Indo-Pacific Ocean (E. merra). These SSLs ranged from one to twelve months. The latitude-SSL correlations in the above-mentioned latitudes were negative in two cases and positive in seven cases, but none were statistically different from zero (Fig. 1). The minimum SPST-SSL correlations were negative for all mentioned species and were statistically significant for E. itajara (Spearman R = -0.71) and E. guttatus (-0.41), indicating that low values of SSL corresponded to high values of minimum SPST (Table 1). Some maximum SPST-SSL and average SPST-SSL correlations were negative whereas, others were positive and the maximum SPST-SSL correlations were only statistically significant for M. bonaci (Spearman R = 0.75), E. guttatus (Spearman R = 0.41) and E. striatus (Spearman R = 0.35), indicating that longer SSL values corresponded with higher maximum SPSTs (Table 1).
| Table 1: | Spearman rank correlation of the minimum, maximum and average temperature during spawning season respect to spawning season length of some tropical groupers |
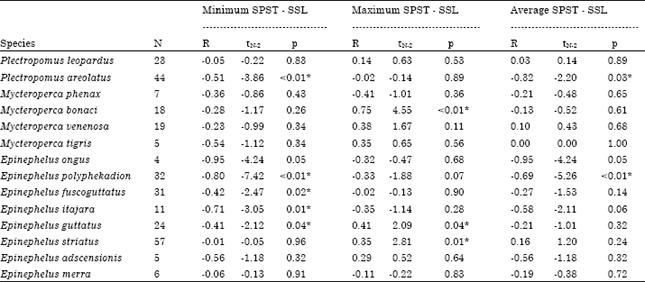 | |
| SSL: Spawning season length, SPST: Spawning season temperature, N: No. of data, R: Spearman rank correlation coefficient, tN-2: Student’s t-test with N (number of data) less two degrees of freedom, p: probability of significance, *Significant correlation | |
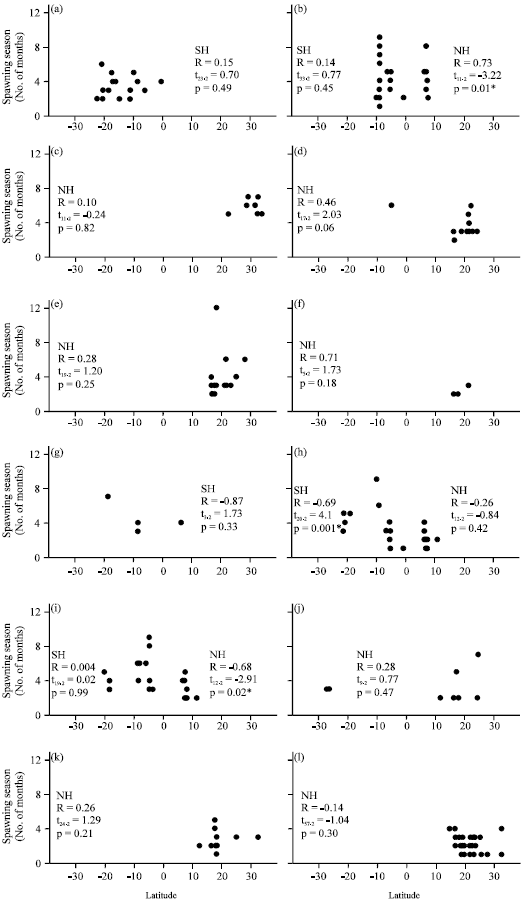 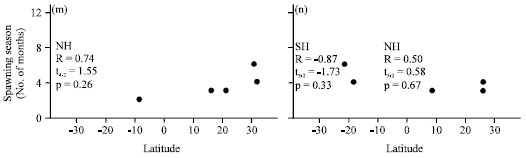 | |
| Fig. 1(a-n): | Relationships of spawning season lengths, showing distribution with respect to latitude. SH, southern, NH, northern, R, the Spearman rank correlation, t, the t-test and p, probability level where, *indicates a statistically significant relationship, (a) Plectropomus leopardus, (b) Plectropomus areolatus, (c) Mycteroperca phenax, (d) Mycteroperca bonaci, (e) Mycteroperca venenosa, (f) Mycteroperca tigris, (g) Epinephelus ongus, (h) Epinephelus polyphekadion, (i) Epinephelus fuscoguttatus, (j) Epinephelus itajara, (k) Epinephelus guttatus, (l) Epinephelus striatus, (m) Epinephelus adscensionis and (n) Epinephelus merra |
SSL data for other species from the Indo-Pacific Ocean (P. leopardus, P. areolatus, E. ongus, E. polyphekadion and E. fuscoguttatus) showed a patchy distribution mostly below 10° Lat, where SSL ranges from one to nine months. The latitude-SSL correlations were negative in the northern hemisphere between 0° Lat and 10° Lat and were statistically significant for P. areolatus (Spearman R = -0.73) and P. fuscoguttatus (Spearman R = -0.68), indicating that the shorter SSLs occurred at higher latitudes. However, the latitude-SSL correlations between 0° Lat and -25° Lat in the southern hemisphere were both negative and positive and were statistically significant for E. polyphekadion (Spearman R = -0.69), indicating that SSLs were longer at more southerly latitudes (Fig. 1). The minimum SPST-SSL correlations were negative for all species from the Indo-Pacific Ocean and were statistically different from zero for P. areolatus (Spearman R = -0.51), E. polyphekadion (Spearman R = -0.80) and E. fuscoguttatus (Spearman R = -0.42), indicating that low values of SSL corresponded to high values of minimum SPST (Table 1). The maximum SPST-SSL and average SPST-SSL correlations were negative in most cases except for P. leopardus. The average SPST-SSL correlations were only significant for P. areolatus (Spearman R = -0.32) and E. polyphekadion (Spearman R = -0.69), indicating that lower SSL values corresponded to higher average SPSTs (Table 1).
The latitude-SSL correlations were statistically insignificant for all species in both the northern (Spearman R = 0.11, t179 = 1.54, p = 0.13) and the southern (Spearman R = -0.11, t103 = -1.1, p = 0.28) hemispheres. However, the minimum SPST had a negative and significant correlation with SSL in both hemispheres (northern: Spearman R = -0.23, t179 = -3.10, p = 0.002; southern: Spearman R = -0.22, t103 = -2.30, p = 0.02). The maximum SPST-SSL correlation was significant in the northern hemisphere (Spearman R = 0.35, t179 = 5, p<0.001), although the average SPST-SSL correlation was not (Spearman R = -0.06, t179 = -0.74, p = 0.46). The correlations of maximum SPST-SSL (Spearman R = 0.05, t103 = 0.54, p = 0.59) and average SPST-SSL (Spearman R = -0.11, t103, p = 0.27) were not statistically significant in the southern hemisphere. Figure 2 shows the distribution of all SSL and temperature data with respect to latitude, where SSLs from one to six months occurred across the entire range of latitudes (-30° to 35°), but most spawning seasons that were longer than six months were observed at latitudes lower than -4° or higher than 10°.
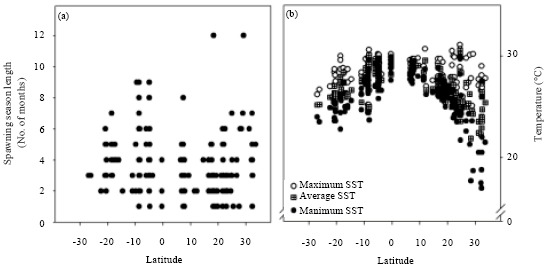 | |
| Fig. 2(a-b): | Distribution showing global trends for (a) Spawning season length and (b) Spawning season temperatures with respect to latitude |
Spawning aggregation duration: The Spawning Aggregation Duration (SAD) was correlated with latitude for two Plectropomus spp., two Mycteroperca spp. and four Epinephelus spp., (Fig. 3) from areas southeast of the Mexican Caribbean Sea and from the Bahamas, the Cayman Islands, the Virgin Islands, Puerto Rico and Belize in the Atlantic Ocean and from the Federated States of Micronesia, the Maldives Islands, Papua New Guinea and the Great Barrier Reef in the Indo-Pacific Ocean.
Most of the latitude-SAD correlations were negative except for P. areolatus, M. tigris and E. fuscoguttatus in the northern hemisphere. Only the correlation of SAD-latitude for E. striatus (Spearman R = -0.36) was statistically different from zero, indicating that lower values of SAD corresponded to higher latitudes (Fig. 3).
The latitude-SAD correlation was statistically insignificant for all species in both the northern (Spearman R = -0.03, t72 = 1.54, p = -0.28) and southern hemispheres (Spearman R = 0.25, t44 = 0.25, p = 0.09).
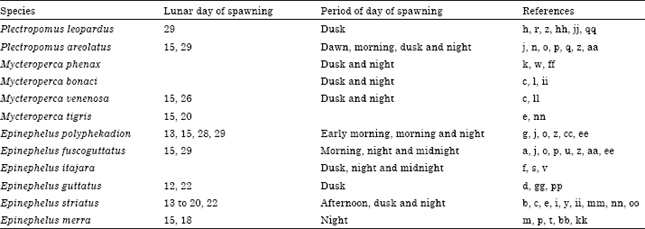
Lunar day of spawning: Few spawning events have been observed for the Plectropomus spp., Mycteroperca spp. and Epinephelus spp., included in this study. Spawnings have been observed near or within the full and third-quarter phases of the moon in species from the Atlantic Ocean and an occur from two days before the full moon to the third quarter in E. striatus. In the Indo-Pacific Ocean, spawnings for P. areolatus, E. polyphekadion and E. fuscoguttatus have been observed near or within the full and new phases of the moon, whereas spawning has been observed during the new moon for P. leopardus and around the full moon for E. merra (Table 2).
Time of day of spawning: Spawning activity has been recorded between afternoon and midnight in species from the Atlantic Ocean. In the Indo-Pacific Ocean, spawnings for P. areolatus, E. polyphekadion and E. fuscoguttatus can occur in the early morning to morning and from dusk to midnight. P. leopardus can spawn at dusk and E. merra can spawn during the night (Table 2).
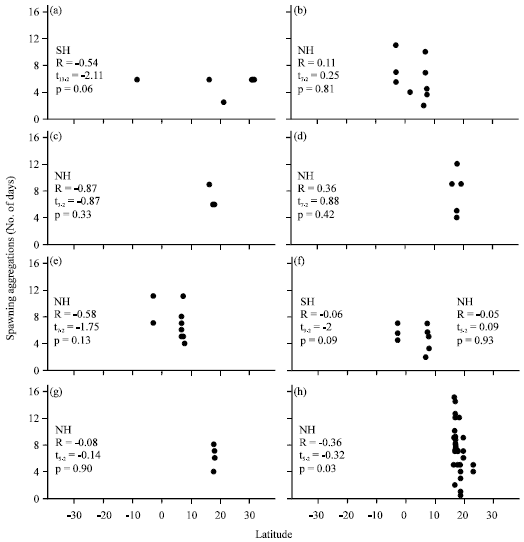 | |
| Fig. 3(a-h): | Relationships between spawning aggregation duration and latitude. SH, southern, NH, northern, R, the Spearman rank correlation, t: the t-test and p, probability level, (a) Plectropomus leopardus, (b) Plectropomus areolatus, (c) Mycteroperca venenosa, (d) Mycteroperca tigris, (e) Epinephelus polyphekadion, (f) Epinephelus fuscoguttatus, (g) Epinephelus guttatus and (h) Epinephelus striatus |
DISCUSSION
The spawning seasons of marine fishes are characteristically longer at latitudes nearer the equator (Johannes, 1978). Our analysis showed variable trends for SSL with respect to latitude in the tropics but only three cases were statistically significant, showing longer spawning periods nearer to the equator for P. areolatus and E. fuscoguttatus in the northern hemisphere and shorter spawning periods for E. polyphekadion in southern hemisphere. For most species, latitude was not a significant source of variation in SSL. This could be due to weak correlations, few records, or intraspecific inconsistencies, for example, E. polyphekadion, whose SSL increases closer to the equator in the northern hemisphere and decreases closer to the equator in the southern hemisphere.
Temperature is a common variable influencing the time at which fishes spawn in tropical areas (Colin, 2012; Craig, 1998; Pankhurst and Porter, 2003).
Consistently, the results showed that the minimum SPST tended to be lower for longer spawning seasons, in individual species as well as for the whole dataset. In fact, we observed SSLs from one to six months across the entire range of latitude (-30° to 35°), but most spawning seasons longer than six months were observed at latitudes lower than 4°S or higher than 10°N, where lower water temperatures can be found (Lowe-McConnell, 1987; Colin, 2012). Additionally, this suggests that the high intraspecific variability of SSLs in similar ranges of latitude could be associated with the presence of different temperature regimes (variation in temperature between locations) at similar latitudes due to differences in habitats, such as those cases where neritic waters are more dynamic with greater variation in temperatures than deeper waters (Lowe-McConnell, 1987).
The variability in SSL suggests that there is a need to scrutinise the reproductive ecology of fishes and compare different tropical habitats for a better understanding of the variability in reproductive tactics. In addition, the minimum SPST could be investigated as an indicator of SSL for fish resources. Hypothetically, we could predict that if the minimum SPST were to rise in an anomalously warm year with respect to a normal year, a certain reduction in SSL could be expected. This could be useful for decision making processes in fisheries.
We are not aware of any studies that relate latitude to SAD; we found variable tendencies between species, but only one such relationship was statistically significant. We found that SAD is lower at higher latitudes for E. striatus across a short latitude interval (15° to 25° Lat). For E. striatus, the relationship of SSL with latitude was not significant but was clearly negative, suggesting that a shorter SSL occurs at higher latitudes. Statistically, the error level could be high in the observation data and, consequently, in the distributions of the data, as field observations of this type are difficult; however, if the above tendency is assumed to be realistic, a shorter SAD could correspond to shorter SSLs in some areas between 10° and 30° Lat. This result suggests that reproductive output could be lower at higher latitudes in E. striatus and also that, hypothetically, some environmental conditions could be determining the SAD (Robertson, 1991). It is known that some spawning aggregations have disappeared in some areas of Mexico, the Bahamas, Belize, Puerto Rico, Bermuda and the Cayman Islands which is usually explained by over-fishing (Aguilar-Perera, 2006; Colin, 1992; Johannes et al., 1999; Sadovy and Eklund, 1999; Sala et al., 2001; Whaylen et al., 2004). If environmental conditions cause a low SSL in some areas, those spawning sites could be more vulnerable to fishing activities than other areas; although our results indicate that this could be possible, more research is needed.
Lunar cues influence the behaviour and reproductive activities of various species of fish (Johannes, 1978; Taylor, 1984; Robertson et al., 1990; Robertson, 1991), synchronising spawning activity at intervals of one month (lunar spawning cycle), two weeks (semi-lunar spawning cycle) or daily based on tidal changes (tidal spawning season) (Takemura et al., 2010). Present results suggest that some grouper species probably follow a lunar spawning cycle which may have a relationship with the timing of spawning behaviour between afternoon and midnight. Others may exhibit semilunar spawning, which could be related to spawning between the early morning and morning and from dusk to midnight. These spawning periodicities are based on the lunar days on which spawning has been observed (Robertson et al., 1990; Robertson, 1991; Takemura et al., 2010). For some species and localities, spawning is highly predictable during the spawning season because of lunar periodicities, making species vulnerable to over-fishing (Claro and Lindeman, 2003; Claydon, 2005; Domeier and Colin, 1997; Johannes et al., 1999).
CONCLUSION
The variability in SSL for tropical Epinephelidae fishes indicated that spawning seasons could be longer at cooler latitudes. Our analysis also suggests that the minimum SPST is probably a good indicator of SSL for several fish resources. We also found evidence that for E. striatus, SAD may vary with latitude with lower values towards higher latitudes. We propose two hypotheses: first, that a shorter SAD could correspond to a shorter SSL in E. striatus and second, that lunar and semilunar spawning behaviour could be associated with specific daily spawning patterns.
ACKNOWLEDGMENTS
The first author thanks CONACyT (scholarship) and the PIFI-Program of the National Polytechnic Institute, IPN for their support. Additional thanks for support through the SEP-CONACyT (104974) and SIP-IPN (20112444) projects. The second author is thankful for support through the COFAA and EDI programs of the IPN.
REFERENCES
- Aguilar-Perera, A., 2006. Disappearance of a Nassau grouper spawning aggregation off the Southern Mexican Caribbean coast. Mar. Ecol. Prog. Ser., 327: 289-296.
CrossRef - Bouriga, N., M. Cherif, G. Hajjej, S. Selmi, J.P. Quignard, E. Faure and M. Trabelsi, 2011. Growth, reproduction and seasonal variation in the fatty acid composition of the sand smelt Atherina sp. from kerkennah islands, Tunisia. J. Fish. Aquat. Sci., 6: 322-333.
CrossRefDirect Link - Chelemal, M., S. Jamili and I. Sharifpour, 2009. Reproductive biology and histological studies in Abu Mullet, Liza abu in the water of the Khozestan province. J. Fish. Aquat. Sci., 4: 1-11.
CrossRefDirect Link - Colin, P.L., 1992. Reproduction of the Nassau grouper, Epinephelus striatus (Pisces: Serranidae) and its relationship to environmental conditions. Environ. Biol. Fish., 34: 357-377.
CrossRef - Colin, P.L., 2010. Aggregation and spawning of the humphead wrasse Cheilinus undulatus (Pisces: Labridae): General aspects of spawning behaviour. J. Fish Biol., 76: 987-1007.
CrossRef - Colin, P.L., D.Y. Shapiro and D. Weiler, 1987. Aspects of the reproduction of two groupers, Epinephelus guttatus and E. striatus in the West Indies. Bull. Mar. Sci., 40: 220-230.
Direct Link - Craig, P.C., 1998. Temporal spawning patterns of several surgeonfishes and wrasses in American Samoa. Pac. Sci., 1: 35-39.
Direct Link - Doherty, P.J., A.J. Fowler, M.A. Samoilys and D.A. Harris, 1994. Monitoring the replenishment of coral trout (Pisces: Serranidae) populations. Bull. Mar. Sci., 54: 343-355.
Direct Link - Domeier, M.L. and P.L. Colin, 1997. Tropical reef fish spawning aggregations: Defined and reviewed. Bull. Mar. Sci., 60: 698-726.
Direct Link - Hamilton, R.J., M. Matawai and T. Potuku, 2004. Spawning aggregations of coral reef fish in New Ireland and Manus provinces, Papua New Guinea: Local knowledge field survey report. TNC Pacific Island Countries Report No. 4/04, The Nature Conservancy, Papua Nueva Guinea. http://pdf.usaid.gov/pdf_docs/PNADI905.pdf.
- Harris, P.J., D.M. Wyanski, D.B. White and J.L. Moore, 2002. Age, growth and reproduction of scamp, Mycteroperca phenax, in the Southwestern North Atlantic, 1979-1997. Bull. Mar. Sci., 70: 113-132.
Direct Link - Heyman W.D. and B. Kjerfve, 2008. Characterization of transient multi-species reef fish spawning aggregations at Gladden Spit, Belize. Bull. Mar. Sci., 83: 531-551.
Direct Link - Hosseini, S.A., S. Jamili, T. Valinassab, G. Vosoghi and S.M.R. Fatemi, 2009. Feeding and spawning of Sphyraena jello in the North-West of Persian Gulf. J. Fish. Aquat. Sci., 4: 57-62.
CrossRefDirect Link - Jagadis, I., B. Ignatius, D. Kandasami and M.A. Khan, 2007. Natural spawning of honeycomb grouper Epinephelus merra Bloch under captive conditions. J. Mar. Biol. Assoc. India, 49: 65-69.
Direct Link - Johannes, R.E., 1978. Reproductive strategies of coastal marine fishes in the tropics. Environ. Biol. Fish., 3: 65-84.
CrossRef - Johannes, R.E., L. Squire and T. Graham, 1994. Developing a protocol for monitoring spawning aggregations of palauan serranids to facilitate the formulation and evaluation of strategies for their management. Report No. 94/28, Forum Fisheries Agency, Solomon Islands. http://www.spc.int/digitallibrary/doc/fame/ffa/reports/ffa_1994_028.pdf.
- Johannes, R.E., L. Squire, T. Graham, Y. Sadovy and H. Renguul, 1999. Spawning aggregations of groupers (Serranidae) in Palau. Marine Conservation Research Series Publication No. 1, The Nature Conservancy, pp: 144. http://www.conservationgateway.org/file/spawning-aggregations-groupers-serranidae-palau.
- Kerdgari, M., T. Valinassab, S. Jamili, M.R. Fatemi and F. Kaymaram, 2009. Reproductive biology of the Japanese threadfin bream, Nemipteru japonicus, in the Northern of Persian Gulf. J. Fish. Aquat. Sci., 4: 143-149.
CrossRefDirect Link - Khalil, N.A., A.E.H.E. El-Gamal, S.A. Gaber and M.A. Mousa, 2007. Immunohistochemical localization of gonadotropin-releasing hormone and somatolactin during sexual maturation and spawning of Lates niloticus. J. Biol. Sci., 7: 1102-1111.
CrossRefDirect Link - Lee, Y.D., S.H. Park, A. Takemura and K. Takano, 2002. Histological observations of seasonal reproductive and lunar-related spawning cycles in the female honeycomb grouper Epinephelus merra in Okinawan waters. Fish. Sci., 68: 872-877.
CrossRef - Mann, D.A., J.V. Locascio, F.C. Coleman and C.C. Koening, 2009. Goliath grouper Epinephelus itajara sound production and movement patterns on aggregation sites. Endangered Species Res., 7: 229-236.
CrossRefDirect Link - Pankhurst, N.W. and M.J.R. Porter, 2003. Cold and dark or warm and light: Variations on the theme of environmental control of reproduction. Fish Physiol. Biochem., 28: 385-389.
CrossRef - Pet, J.S., P.J. Mous, A.H. Muljadi, Y.J. Sadovy and L. Squire, 2005. Aggregations of Plectropomus areolatus and Epinephelus fuscoguttatus (groupers, Serranidae) in the Komodo National Park, Indonesia: Monitoring and implications for management. Environ. Biol. Fish., 74: 209-218.
CrossRef - Ramadan, A.M. and M.M. El-Halfawy, 2007. Common forms of Atresia in the ovary of some red sea fishes during reproductive cycle. Pak. J. Biol. Sci., 10: 3120-3125.
CrossRefPubMedDirect Link - Rhodes, K.L. and Y. Sadovy, 2002. Temporal and spatial trends in spawning aggregations of camouflage grouper, Epinephelus polyphekadion, in Pohnpei, Micronesia. Environ. Biol. Fish., 63: 27-39.
CrossRef - Randall, J.E. and V.E. Brock, 1960. Observations on the ecology of Epinepheline and Lutjanid fishes of the Society Islands, with emphasis on food habits. Trans. Am. Fish. Soc., 89: 9-16.
CrossRef - Robertson D.R., C.W. Petersen and J.D. Brawn, 1990. Lunar reproductive cycles of benthic brooding reef fishes: Reflections of larval biology or adult biology? Ecol. Monogr., 60: 311-329.
CrossRef - Robinson, J., M. Samoilys and P. Kimani, 2008. Reef Fish Spawning Aggregations in the Western Indian Ocean: Current Knowledge and Implications for Management. In: Ten Years after Bleaching: Facing the Consequences of Climate Change in the Indian Ocean, Obura, D.O., J. Tamelander and O. Linden (Eds.). CORDIO, Mombasa, Kenya, pp: 263-276.
- Sadeghi, M.S., F. Kaymaram, S. Jamili, M.R. Fatemi and M.S. Mortazavi, 2009. Patterns of reproduction and spawning of the Scomberomorus commerson in the coastal waters of Iran. J. Fish. Aquatic Sci., 4: 32-40.
CrossRefDirect Link - Sadovy, Y., A. Rosario and A. Roman, 1994. Reproduction in an aggregating grouper, the red hind, Epinephelus guttatus. Environ. Biol. Fish., 41: 269-286.
CrossRef - Sala, E., E. Ballesteros and R.M. Starr, 2001. Rapid decline of Nassau grouper spawning aggregations in Belize: Fishery management and conservation needs. Fisheries, 26: 23-30.
CrossRef - Salcedo-Bojorquez, S. and F. Arreguin-Sanchez, 2011. An exploratory analysis to identify reproductive strategies of billfishes. J. Fish. Aquat. Sci., 6: 578-591.
CrossRefDirect Link - Samoilys, M.A., 1997. Periodicity of spawning aggregations of coral trout Plectropomus leopardus (Pisces: Serranidae) on the northern Great Barrier Reef. Mar. Ecol. Prog. Ser., 160: 149-159.
CrossRef - Samoilys, M.A. and L.C. Squire, 1994. Preliminary observations on the spawning behavior of coral trout, Plectropomus leopardus (Pisces: Serranidae), on the Great Barrier Reef. Bull. Mar. Sci., 54: 332-342.
Direct Link - Sedberry, G.R., C.L. Cooksey, S.F. Crowe, J. Hyland, P.C. Jutte, C.M. Ralph and L.R. Sautter, 2004. Characterization of deep reef habitat off the southeastern U.S., with particular emphasis on discovery, exploration and description of reef fish spawning sites. Report, project NA16RP2697. South Carolina: USA, Marine Resources Research Institute.
- Sivashanthini, K., G.A. Charles and S. Shutharshan, 2008. Fecundity studies of Gerres abbreviatus (BLEEKER, 1850) from the Jaffna lagoon, Sri Lanka. J. Fish. Aquatic Sci., 3: 320-327.
CrossRefDirect Link - Soyano, K., T. Masumoto, H. Tanaka, M. Takushima and M. Nakamura, 2003. Lunar related spawning in honeycomb grouper, Epinephelus merra. Fish Physiol. Biochem., 28: 447-448.
CrossRef - Spearman, C., 1904. The proof and measurement of association between two things. Am. J. Psychol., 15: 72-101.
Direct Link - Starr, R.M., E. Sala, E. Ballesteros and M. Zabala, 2007. Spatial dynamics of the nassau grouper (Epinephelus striatus) in a Caribbean atoll. Mar. Ecol. Prog. Ser., 343: 239-249.
CrossRef - Sutharshiny, S. and K. Sivashanthini, 2011. Lipid reserves of Scomberoides lysan (Pisces: Carangidae) from the Sri Lankan waters. Int. J. Biol. Chem., 5: 170-183.
CrossRefDirect Link - Takemura A., M.S. Rahman and Y.J. Park, 2010. External and internal controls of lunar related reproductive rhythms in fishes. J. Fish Biol., 76: 7-26.
CrossRef - Taylor, M.H., 1984. Lunar synchronization of fish reproduction. Trans. Am. Fish. Soc., 113: 484-493.
CrossRef - Tucker, J.W., P.G. Bush and S.T. Slaybaugh, 1993. Reproductive patterns of Cayman Islands nassau grouper (Epinephelus striatus) populations. Bull. Mar. Sci., 52: 961-969.
Direct Link - Whaylen, L., P. Bush, B. Johnson, K.E. Luke and C. Mccoy et al., 2006. Aggregation dynamics and lessons learned from five years of monitoring at a Nassau grouper (Epinephelus striatus) spawning aggregation in Little Cayman, Cayman Islands, BWI. Proceedings of the 59th Gulf and Caribbean Fisheries Institute, November 9-11, 2006, Belize, pp: 413-421.
- Whaylen, L., C.V. Pattengill-Semmens, B.X. Semmens, P.G. Bush and M.R. Boardman, 2004. Observations of a Nassau grouper, Epinephelus striatus, spawning aggregation site in Little Cayman, Cayman Islands, including multi-species spawning information. Environ. Biol. Fish., 70: 305-313.
CrossRef - Whiteman, E.A., C.A. Jennings and R.S. Nemeth, 2005. Sex structure and potential female fecundity in a Epinephelus guttatus spawning aggregation: Applying ultrasonic imaging. J. Fish Biol., 66: 983-995.
CrossRef - Zeller, D.C., 1998. Spawning aggregations: Patterns of movement of the coral trout Plectropomus leopardus (Serranidae) as determined by ultrasonic telemetry. Mar. Ecol. Prog. Ser., 162: 253-263.
CrossRef