
M. Gallardo-Cabello
Instituto de Ciencias del Mary Limnologia, Universidad Nacional Autonoma de Mexico, Av. Ciudad Universitaria 3000, Col. Copilco, D.F. (C.P. 04360), Mexico
E. Espino-Barr
Instituto Nacional de Pesca, Centro Regional de Investigacion Pesquera, Playa Ventanas s/n, Manzanillo, Colima (C.P. 28200), Mexico
E.G. Cabral-Solis
Instituto Nacional de Pesca, Centro Regional de Investigacion Pesquera, Playa Ventanas s/n, Manzanillo, Colima (C.P. 28200), Mexico
M. Puente-Gomez
Instituto Nacional de Pesca, Centro Regional de Investigacion Pesquera, Playa Ventanas s/n, Manzanillo, Colima (C.P. 28200), Mexico
A. Garcia-Boa
Instituto Nacional de Pesca, Centro Regional de Investigacion Pesquera, Playa Ventanas s/n, Manzanillo, Colima (C.P. 28200), Mexico
Journal of Fisheries and Aquatic Science
Year: 2012 | Volume: 7 | Issue: 6 | Page No.: 346-363







ABSTRACT
Morphology, morphometry and growth rings of the otoliths: sagitta, asteriscus and lapillus of Mugil cephalus from Central Mexican Pacific were analyzed. The three pairs of otoliths (sagitta, asteriscus and lapillus), right and left and sex differences were studied. The samples were obtained from the commercial catch in the months of August to December 2007, January to March 2008 and July 2011. In all cases it was observed that growth in otoliths is eccentric with respect to the core. The distance from the core to each age growth ring was measured. Relations between total length of the fish and length and width of the otolith showed that these structures can be used to determine the age in fish. Ten growth rings were identified in sagittae and asterisci. Due to its thickness, these marks could not be seen in lapilli. During the first stages of development, female sagittae and lapilli have larger size but once they reach sexual maturity, males’ otoliths reach larger sizes.
PDF Abstract XML References Citation
Received: December 06, 2011;
Accepted: June 19, 2012;
Published: August 06, 2012
How to cite this article
M. Gallardo-Cabello, E. Espino-Barr, E.G. Cabral-Solis, M. Puente-Gomez and A. Garcia-Boa, 2012. Study of the Otoliths of Stripped Mullet Mugil cephalus Linnaeus, 1758 in Mexican Central Pacific. Journal of Fisheries and Aquatic Science, 7: 346-363.
DOI: 10.3923/jfas.2012.346.363
URL: https://scialert.net/abstract/?doi=jfas.2012.346.363
DOI: 10.3923/jfas.2012.346.363
URL: https://scialert.net/abstract/?doi=jfas.2012.346.363
INTRODUCTION
Stripped mullet Mugil cephalus Linnaeus, 1758 is a circumtropical distribution species that also can be found in temperate areas, limited by a 15°C isotherm. In the American Pacific coast it lives from California to Chile, including the Gulf of California and Galapagos Islands. It is found in mud and sandy bottoms but also rocky shores, from the coast line to 120 m deep. It can tolerate high salinity variations, from hipersaline to fresh water; it swims sometimes in rivers but is more abundant in bays, brackish coastal lagoons and estuaries. Usually it forms big schools and swims away from shore to spawn, mainly during fall and winter months (Allen and Robertson, 1994; Ibañez-Aguirre and Gallardo-Cabello, 1996).
The striped mullet is intensively fished with gill nets, cast nets and shore nets. Its meat is consumed and appreciated, but the gonad “roe” has higher demand and price. This fish is also used as bait (Fischer et al., 1995).
Capture of M. cephalus in México during 2010 was of 9 079 metric tons (mt), of which 4 656 mt (51.3%) were obtained in the Atlantic Ocean and 4 423 mt (48.7%) in the Pacific. In the coastal lagoons of Jalisco and Colima, area where samplings of the present paper were done, 53 mt were obtained that is 1.20% of the Pacific production. The fishery of M. cephalus is part of the coastal artisanal fishery (small scale fishery), together with Mugil curema (Cabral-Solís et al., 2007) and both are socially very important, because they generates a large number of employments and offer high protein food (Ravichandran et al., 2011) at a low price for the lower classes which is $18.00 kg-1 ($1.28 US dollars). This species has also been used in aquaculture with important results (Shofiquzzoha et al., 2001) and has proven to be a good indicator in metal accumulation in lagoons (Authman and Abbas, 2007).
Studies of these three pairs of otoliths (sagitta, asteriscus and lapillus) have been done in larvae of different species by Victor and Brothers (1982), Brothers et al. (1983), Solomon et al. (1985), Suthers et al. (1989), Barkman (1978), Bolz and Lough (1983, 1988), Lagardere (1989) and David et al. (1994) and in adult fish by Gallardo-Cabello et al. (2006), Espino-Barr et al. (2005), Santana-Hernandez et al. (2008), Granados-Flores et al. (2010) and Gallardo-Cabello et al. (2011). Study of otoliths can be for diverse reasons. They have proven useful in the determination of anthropogenic contamination (Kamaruzzaman et al., 2011).
Otoliths also show age groups of the stock and growth analysis, one of the fundamental parameters to formulate models on maximum sustainable captures, yield per recruit, predictions and catch simulations (Campana and Neilson, 1985; Sparre and Venema, 1995; Mascarena-Osorio et al., 2003; Gallardo-Cabello et al., 2003; Begg et al., 2005; Espino-Barr et al., 2005; Gallardo-Cabello et al., 2007; Espino-Barr et al., 2010). These models are to regulate the fishery administration to avoid overproduction, capturing organisms that have reproduced at least once and limiting young specimens still sexually immature (Francis et al., 2005; Berg et al., 2005; Stevensen and Campana, 1992; Ehrhardt, 1981). Studies on fisheries are of great importance and should be thoroughly assessed for their sustainability (Eide and Wikan, 2010). This is why the analysis of otoliths in age determination is one of the most important bases in population dynamics studies. Therefore, this study presents, for the first time, a study on morphology and morphometry and growth ring identification in the otoliths: sagittae, asterisci and lapilli of M. cephalus.
MATERIALS AND METHODS
Specimens were obtained from the commercial fishery in the Cuyutlán Lagoon, Colima, México (103°57'-104°19'W and 18°57'-19°50'N) and in Cruz de Loreto Lagoon, Jalisco, México (105°27'-105°33'W and 19°58'-20°05'N). The fishing gears were gill-nets of 2.0, 2.5, 3, 3.5 and 4 inches mesh size (5.08, 6.35, 7.62, 8.89, 10.16 cm). Samples were obtained from August to December 2007, January to March 2008 and July 2011 (months in which this species was available). Total length was measured to the nearest mm (from the snout tip to the caudal fin extreme for 144 organisms. The size of the sample was determined according to Daniel (1991).
Sex and gonad maturation were determined in visu on fresh organisms taken to the laboratory the same day they were caught. The stages of sexual maturity were determined using the key by Diaz-Pardo and Hernandez-Vazquez (1980) where stage I is juvenile fish; in stage II gonads fill 1/3 of the abdominal cavity; in stage III gonads occupy ½ to ¾ of the abdominal cavity and oocytes are not individualized; in stage IV, pre-spawning stage, gonads occupy ¾ of the entire abdominal cavity and oocytes are individualized; stage V is the spawning stage where gonads occupy the total abdominal cavity and oocytes and semen can be released by squeezing the body; stage VI is post-spawning.Otoliths, sagittae, asterisci and lapilli were obtained through a transverse cut in the ventral cranial cavity, the brain was removed and the left and right semicircular canals were extracted. The otoliths were rinsed with water and stored dry in Eppendorf tubes labeled with number, date, total length and sex.
The structure of otoliths was studied with a dissection microscope. A description of the labyrinth system and the sagittae was made with the terminology of the glossary of Secor et al. (1992). The same concepts were applied to the description of the asterisci and lapilli according to Gallardo-Cabello et al. (2006, 2011) and Espino-Barr et al. (2006).
Data on length and width were registered for each otolith through their observation in a stereoscopic microscope with a graduated ocular lens. Measures were taken on the right and left aspect for the three pairs.
Constants of the relationships of the sagittae, asterisci and lapilli were calculated for Rostrum Length (RL) and Width (Wi). The relationships between the total length of the fish and all the measures of the three otoliths were also recorded. Regressions were done by the least squares. For the evaluation of the relations and the analysis of the possible morphometric differences between the otoliths of males and females, a one way variance analysis (ANOVA) was carried out (Zar, 1996).
The identification of growth rings was done observing the sagittae and the asterisci in the stereoscopic microscope with transmitted light and the average length of the fish was calculated.
The structure and microstructure of the otoliths were studied with a scanning electronic microscope, from the Institute of Physics of the Universidad Nacional Autónoma de México.
Statistical analysis: Constants of the relationships of otoliths and with total length of the fish were calculated by the least squares. To evaluate differences of these regressions and morphometric differences between males and females, a one way variance analysis (ANOVA) was carried out (Mendenhall, 1987; Zar, 1996).
RESULTS
Morphologic description of otoliths of Mugil cephalus
Labyrinth system of Mugil cephalus: The membranous labyrinth is formed by a tubular system of semicircular canals named: anterior vertical canal (AVSC), posterior vertical canal (PVSC) (Fig. 1) and horizontal canal. The semicircular canals form chambers where the otoliths are contained, the sagitta is included in the sacculus, the asteriscus is included in the lagena (Fig. 2) and the lapillus is included in the utriculus (Fig. 1). The chambers where the otoliths are included are full of a liquid called endolymph (Lagler et al., 1962).
The otoliths are in contact with the acoustic macula that enters in the acoustic canal of the sagitta (Mugiya, 1964, 1966a, b) and the sulcus of the asteriscus and the lapillus (Gallardo-Cabello et al., 2006; Espino-Barr et al., 2006; Santana-Hernandez et al., 2008; Granados-Flores et al., 2010; Gallardo-Cabello et al., 2011). Otolith growth is possible because nutrient deposition occurs through acoustic macula (Gallardo-Cabello et al., 2006, 2011; Espino-Barr et al., 2006). Vibrations are transmitted from the otoliths through the acoustic macula to the eighth cranial nerve and to the brain (Mugiya, 1964, 1966a, b). Sagittae and asterisci are responsible for sound perception, gravity and angular acceleration. On the other hand balance is related with lapilli (Holst et al., 1950; Lowenstein, 1957).
The sagittae are made of calcic carbonate crystals (Lagler et al., 1962) in the form of aragonite (Hickling, 1931; Sasaki and Miyata, 1955; Carlstrom, 1963; Gallardo-Cabello, 1986) and a high molecular weight protein named otoline (Degens et al., 1969).
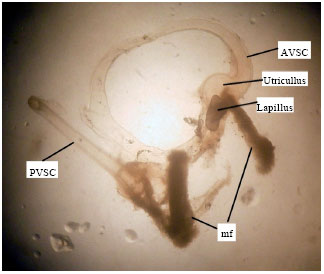 | |
| Fig. 1: | Lapillus contained in utriculus in an individual of Mugil cephalus (25 cm of total length) (increased 6.5 times). AVSC: Anterior vertical semicircular canal, PVSC: Posterior vertical semicircular canal and mf: Macula fragments |
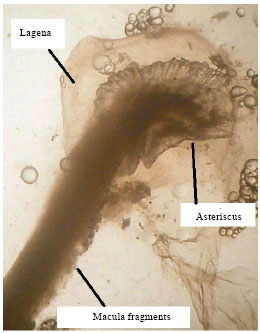 | |
| Fig. 2: | Asteriscus in lagena of the membranous labyrinth in an individual of Mugil cephalus (25 cm total length, increased 19 times) showing the asteriscus with the macula in its acoustic canal |
The largest otolith of Mugil cephalus is sagitta reaching a total length of 7 mm while asteriscus measures 1.28 mm and lapillus 1.23 mm in individuals of 25 cm total length (Fig. 3).
Description of the sagitta: The sagitta shows in its anterior margin a rostrum in the form of a pointed arrow, without antirostrum and therefore no excisura major. The shape of the rostrum varies as the fish ages, becoming more rounded in older fish (Fig. 4).
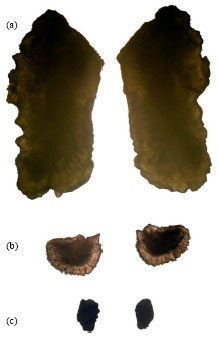 | |
| Fig. 3(a-c): | Left and right otoliths of Mugil cephalus: (a) Sagittae, (b) Asterisci and (c) Lapilli (increased 6 times) |
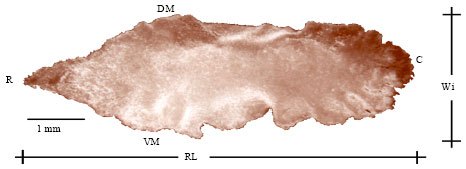 | |
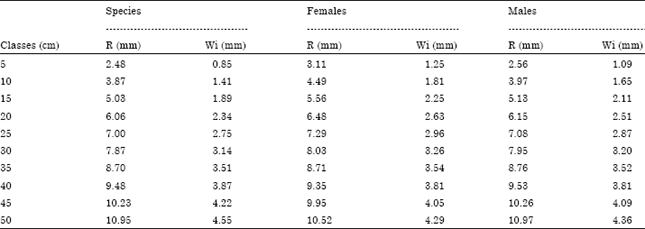
| Fig. 4: | Left sagitta, external aspect of Mugil cephalus of 25 cm RL: Rostrum length, R: Rostrum, C: Postrostrum, DM: Dorsal margin, VM: Ventral margin and Wi: Width |
These otoliths vary in their form between individuals and also in right and left sagittae of the same specimen. A rounded postrostrum is observed at the rear section of the sagitta without excisura minor and therefore no pararostrum. The dorsal margin shows a pointed rostrum and a round postrostrum with a profile that tends to be rectilinear in the middle section of the otolith. The ventral side shows big irregularities which are bigger in the middle section of sagitta (Fig. 4).
The external aspect of the sagitta is concave a feature which increases with age. The internal aspect of the sagitta is convex without variations in its thickness, its surface is smooth and it is totally traversed by the acoustic channel (Fig. 5); its width from the anterior to the posterior part of the otolith is the same, therefore no differentiation between ostium and cauda was observed.
The sulcus base shows calcium carbonate crystals arranged epitaxially with different growth patterns, these characteristics could increase the impulse transmission through the acoustic macula to the brain (Fig. 6a, b). Mean width of the sagitta is 2.54 times its average length.
| Table 1: | ANOVA statistical results for sagitta |
 | |
 | |
| Fig. 5: | Scanning photograph of the internal aspect of the right sagitta of M. cephalus showing the acoustic canal |
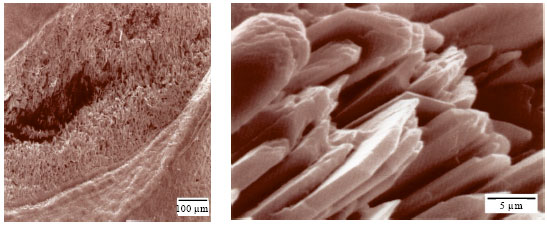 | |
| Fig. 6: | Scanning photograph on the base of the acoustic canal, in the right sagitta internal aspect of M. cephalus, showing the calcic carbonate crystals |
Even though there is visual difference in the shapes and details of individual sagitta, there was no statistical difference between left and right sagittae (F’0.05 (2, 86 = 3.952) = 0.061). Also, there were no statistical difference between male’s and female’s sagittae shapes (F’0.05 (2, 59 = 4.010) = 0.495) (Table 1).
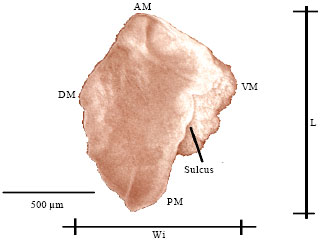
Description of the asteriscus: This otolith is divided in two parts in its anterior margin by a blunt projection showing a dorsal part with a larger surface than the ventral one.
| Table 2: | ANOVA statistical results for asteriscus |
 | |
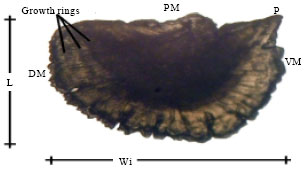 | |
| Fig. 7: | Scanning photograph of the left asteriscus, external aspect of M. cephalus, AM: Anterior margin, PM: Posterior margin, P: Projection, DM: Dorsal margin, VM: Ventral margin, L: Length, Wi: Width and GR: Growth rings |
 | |
| Fig. 8: | Scanning photograph of the right asteriscus, internal aspect of M. cephalus, AM: Anterior margin, PM: Posterior margin, DM: Dorsal margin and VM: ventral margin |
Some sections of the anterior margin can be rectilinear from the dorsal to the ventral margins. Growth rings can be seen in the external aspect of the asteriscus (Fig. 7).
The posterior margin is rounded showing irregularities in the form of notches. The posterior margin presents grooves all around the dorsal and ventral margins which divide the otolith in two parts. The first part side has larger sized radius in the external aspect (posterior external margin) than in the internal aspect (Fig. 8).
The space between the smaller radius in the internal aspect of the otolith (posterior internal margin) and the larger radius in the external aspect (posterior external margin) forms a sulcus which is in contact with the acoustic macula (Fig. 9). The external aspect of the asteriscus is convex, its curvature increases as the fish ages, the internal aspect is concave.
Asteriscus mean width is 0.621 times its average length.
The shape of the asterisci varies between specimens and between right and left, as in the sagittae but there were no statistical difference (F’0.05 (2, 63 = 4.913) = 1.066). Also between males and females, no statistical difference was found (F’0.05 (2, 52 = 4.079) = 2.555) (Table 2).
| Table 3: | ANOVA statistical results for lapillus |
 | |
 | |
| Fig. 9(a-b): | Scanning photograph of the right asteriscus, internal aspect of M. cephalus, showing details of the sulcus, (a) IF: Internal aspect and EF: External aspect, (b) IPM: Internal posterior margin and EPM: External posterior margin |
Description of the lapillus: The anterior margin of the otolith which is oriented toward the front of the fish is rounded.
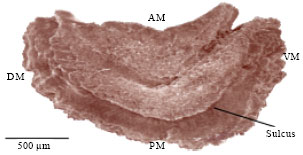
The dorsal margin is rectilinear from the anterior margin to the posterior margin and shows a smooth surface. The ventral margin shows some parts of the sulcus surface just in the middle part of the otolith. The external aspect is convex (Fig. 10).
The inner surface is concave, its curvature increases as fish ages. The posterior border shows the sulcus which extends along the dorsal and ventral margins and enters in contact with the acoustic macula (Fig. 11).
A large number of calcium carbonate crystals in the sulcus are arranged epitaxially showing different growth patterns in shape and size that could increase the impulse transmission from the lapillus to the brain through the eighth cranial nerve (Fig. 12).
Mean width of the lapillus is 1.89 times its average length.
As in the other two otoliths, visual details were noticed in lapilli, although statistical differences could not be found between right and left (F’0.05 (2, 74 = 3.974) = 0.017) and between males and females (F’0.05 (2, 51 = 4.047) = 0.565) (Table 3).
Morphometric analysis of otoliths of Mugil cephalus
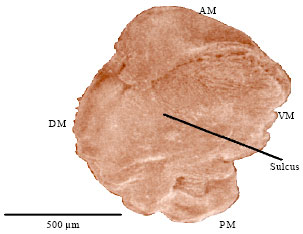
Growth of the sagitta: Table 4 shows the relation between rostrum and width of sagitta and the length classes of fish, for the species and sexes. Growth of the rostrum and width of sagitta are smaller in males during the first and middle stages that is, lengths 5-30 cm; thereafter 35-50 cm growth rate decreases in females and it is higher in males’ sagittae.
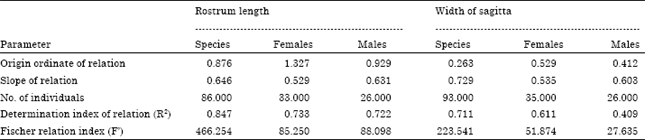
The relationship between length and width of sagitta is expressed by the exponent value b = 0.943 which shows a tendency to an isometric growth (Table 5) indicating that the sagitta tends to keep the same shape as the fish ages.
| Table 4: | Measures of rostrum (R) and width (Wi) at different size classes of the sagitta of Mugil cephalus (n = 86) |
 | |
 | |
| Fig. 10: | Scanning photography of the left lapillus external aspect of M. cephalus, AM: Anterior margin, PM: Posterior margin, DM: Dorsal margin, VM: Ventral margin, L: Length and Wi: Width |
 | |
| Fig. 11: | Scanning photograph of the right lapillus, internal aspect of M. cephalus, AM: Anterior margin, PM: Posterior margin, DM: Dorsal margin, VM: ventral margin, L: length and Wi: width |
| Table 5: | Relationship between rostrum length and width of sagitta of M. cephalus |
 | |
| Table 6: | Relationship between the total length of the fish and the measures of sagitta of M. cephalus |
 | |
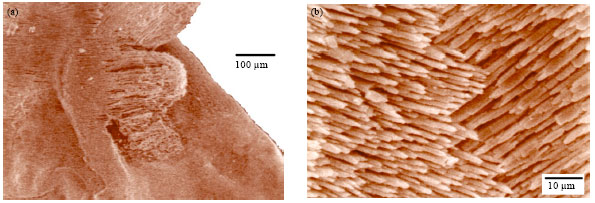 | |
| Fig. 12(a-b): | Scanning photograph of the right lapillus, internal aspect of M. cephalus, showing the calcic carbonate crystals in the base of the sulcus |
In the case of sexes results vary and the relations of males are b = 0.804 and females b = 0.603 which show a negative allometric growth. These results indicate that the growth of sagitta is eccentric to the core which means that when the fish ages the postrostrum grows more than the rostrum. The dorsal edge grows more than the ventral edge and a larger amount of material accumulates on the inner than the external aspect of the sagitta.
The relationship between fish length and length and width of sagitta is shown in Table 6. The higher value of the allometric index relating fish length to rostrum’s width is for the species with a value of b = 0.729, smaller values are found for the males and females, b = 0.603 and b = 0.535, respectively. Similarly, the relation between fish length and rostrum length for the species has a value of b = 0.646 (0.631 for males and 0.529 for females). In all cases, the values show that there is a direct proportionality between sagitta length, width and fish length; this structure is adequate to describe the growth of the organism. High values of the ANOVA (F’) show a strong correlation between the structures analyzed in each case. Values of allometric growth rates are below one, due to the difficulty of relating very small structures (mm) with values of total fish length, expressed in centimeters.
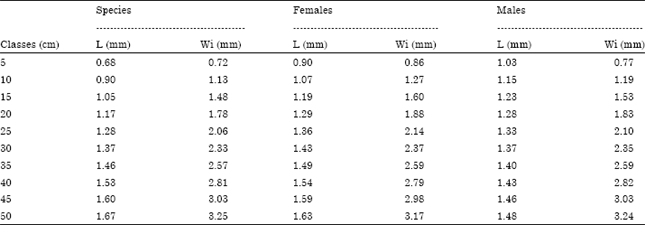
| Table 7: | Measures of length (L) and width (Wi) of asteriscus at different length classes of M. cephalus (n = 63) |
 | |
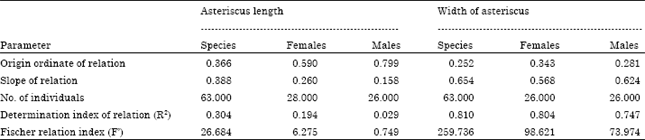
| Table 8: | Relationship between the length and width of the asteriscus of M. cephalus |
 | |
| Table 9: | Relationship between total length of the fish and measures of the asteriscus of M. cephalus |
 | |
Growth of the asteriscus: The relationship between fish length and length and width of asteriscus is shown in Table 7. In this case growth of asteriscus is higher in males during the early stages.
The relationship between the length and width of the otolith (Table 8) is described by the allometric index b = 0.608 (R2 = 0.360 and F’ = 32.665) with similar values for females and males, the latter having a small value of the allometric index of 0.175. These results show a negative allometric growth in which the increase in width is greater than length, so, asteriscus growth is eccentric to the core; therefore its anterior border grows more than the posterior border and the dorsal margin grows more than the ventral margin.
Table 9 shows the relationship between fish total length and length and width of asteriscus. The allometric index value closest to one is for species b = 0.654, for fish total length and asteriscus width. Values of the ANOVA (F’) show a correlation between the structures analyzed in each case, with a tendency to a direct proportionality between fish length and asteriscus width and length which allows age group determination based on the number of growth rings in asteriscus be valid for M. cephalus.
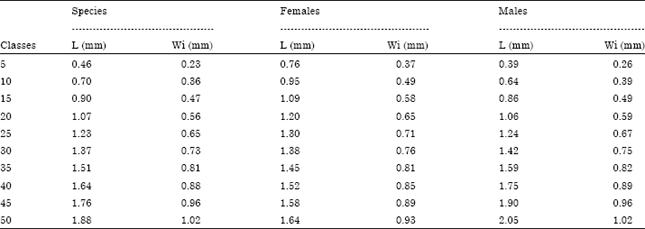
| Table 10: | Measures of length (L, mm) and width (Wi, mm) of lapillus at different length classes of M. cephalus (n = 74) |
 | |
| Table 11: | Relationship between length and width of lapillus of M. cephalus |
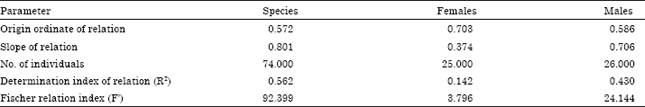 | |
| Table 12: | Relationship between fish length and other measures of lapillus of M. cephalus |
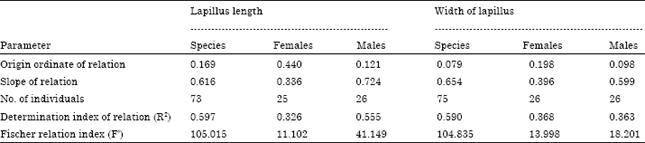 | |
Growth of the lapillus: Table 10 shows the relationship between fish length and length and width of the lapillus. As in the sagitta and asteriscus, the fastest growth of lapillus occurs in females at 30 cm in length.
The relationship between the length and width of the lapillus (Table 11) is described by the allometric index b = 0.801, smaller indexes were found for males: b = 0.706 and females b = 0.374. These values represent a negative allometric growth, in which lapillus grows more in length than in width. Lapillus growth is eccentric to the core which means, the anterior and ventral margins show a higher deposition of growth materials than the posterior and dorsal margins.
The relationship between fish length and the length and width of the lapillus is shown in Table 12. The higher value of the allometric index for fish total length and lapillus length was found for males with a value of b = 0.724, smaller indexes were found for the species and females: b = 0.616 and b = 0.336, respectively. These values suggest a trend toward direct proportionality between fish length and lapillus that validates identification of growth rings in this structure of the M. cephalus. This phenomenon is also observed in other species (Aydin et al., 2004).
| Table 13: | Number of rings on sagittae and scales from several authors and mean length (Lt ±SD) of M. cephalus |
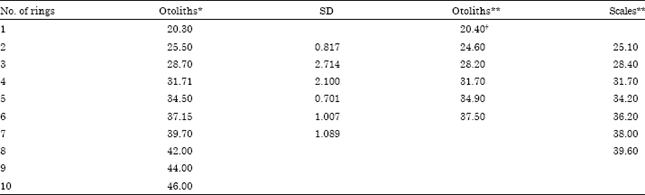 | |
| *Present study, **Ibañez-Aguirre and Gallardo-Cabello (1996) and †Obtained by backcalculation method | |
Identification of growth rings: The identification of ten age groups was possible through growth ring analysis in sagittae of the Mugil cephalus (Table 13). One hundred percent of sagittae showed clearly defined growth rings. Growth rings can be seen in a region on the dorsal margin that goes from the middle of the otolith to the postrostrum; in this region is higher the deposition of growth mater.
In 64% of asterisci appeared the same number of rings as sagittae, in the remaining 36%, growth rings were not clearly identified. Growth rings in asterisci can be seen on the outer side of the otolith as dark concentric lines which run from the dorsal to the ventral margin (Fig. 7).
Growth rings in lapilli are very difficult to see because this structure is very thick which avoids the observation of growth rings by transmitted light transparency.
DISCUSSION
Otoliths extraction is not easy, most of the time they can be broken easily, especially sagittae because of its large size and for this reason the number of items (n) is different for sagittae, asterisci and lapilli.
Although, right and left otoliths of the three pairs of otoliths can show morphologic differences in the same organism, morphometric differences are not statistically significant.
In the sagittae and lapilli, growth during early stages is higher in females than in males but once gonadal maturity starts at 30 cm total length growth decreases in females and is higher in males.
The allometric index values between length and width of sagitta show a high tendency to isometry, because many of the data used for calculations are young and sexually undifferentiated organisms that is that have not yet reached gonadic maturation. Most of their energy is used to grow in length to avoid depredation and also a higher development of structures related with sense organs to maintain alert against depredators attacks and therefore increment survival. Once gonadic maturation begins in M. cephalus (at two years of age, 20 cm of total length), most of the energy is used in sexual organs production that is, in gonadic maturation and also to store fatty acids and growth in weight. That is why in mature organisms sagittae show a tendency to a negative allometry (Gallardo-Cabello, 1986; Gallardo-Cabello et al., 2011). A similar phenomenon occurs in asterisci and lapilli, the higher values of the allometric growth indexes are those in which calculations were made including data on immature individuals.
| Table 14: | Mean length (mm) from nucleus to each fast growth band of the sagitta of M. cephalus |
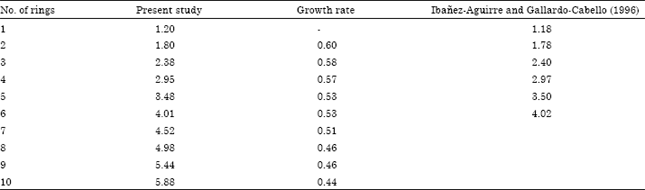 | |
Values of mean size for each growth ring in the present study are very similar to age groups reported from analysis of otoliths of the same species in Tamiahua Lagoon, Veracruz, México in the Gulf of México (Ibañez-Aguirre and Gallardo-Cabello, 1996) (Table 13). Similarly, the mean lengths from the nucleus to each fast growth band on the sagittae of M. cephalus were similar to those in Tamiahua Lagoon (Table 14). In this table it is also observed the existence of a progressive diminution of the extent of the growth rings as the fish grows old. Nevertheless, in this present study larger organisms with 10 growth rings were found. On the other hand, Ibañez-Aguirre and Gallardo-Cabello (1996) reported a higher number of age groups (eight age groups) for this species using scale reading method which can be related with the fact that when individuals grow, the sagittae becomes more curved, making it very difficult to observe the last growth ring on the border of the older specimens which means that the number of age groups could be underestimated in M. cephalus of Tamiahua Lagoon. Another phenomenon that could explain the difference between the number of growth bands in organisms of Tamiahua and those from this study is the fishery pressure. It is likely that the fishing pressure on M. cephalus stocks in Tamiahua Lagoon is higher because of the great demand on the female gonads in northern parts of the Gulf of México and therefore prevents the organisms to grow more than six to eight years. There are previous analysis of overfishing as reported by Garcia (1978a, b and 1980), who showed that during the 70’s the capture included young organisms and mature adults that were fished throughout the year, without considering reproduction seasons or minimum length for extraction. Gomez-Ortiz et al. (2006) reported that in Tamiahua lagoon fishers don’t always use authorized gears to fish M. cephalus, they only use one type of net to fish all kinds of fish; they even use illegal gears, such as troll nets and fixed gears called palisade that affect young organisms of many species.
CONCLUSION
The analysis of sagittae and asterisci allowed a clear identification of growth rings, contrary to lapilli, whose thickness doesn’t allow the transmittance of light and the determination of growth rings by simple view. Morphologic differences of the three pairs were not statistically significant. Dorsal border of sagittae grows more than the ventral and a larger deposit of material goes to the postrostrum than to the rostrum. Relation between length and width of the three pairs of otoliths versus the fish length showed that these structures can be used to determine age in this species. Ten growth rings were identified in sagittae and asterisci, periodicity has to be evaluated. Growth of sagittae and lapilli is higher in males than females once the sexual maturity begins. After the second growth ring of age the three pair of otoliths show a negative allometric growth. A larger number of age groups of M. cephalus are found in Central Mexican Pacific than in Tamiahua Lagoon in the Gulf of México.
ACKNOWLEDGMENTS
The authors wish to thank M. S. Jacqueline Cañetas, technician from the Institute of Physics of the Universidad Nacional Autónoma de México, for her help in the scanning photographs and the fishers who kindly proportionate the samples and always teach us something new on each species.
REFERENCES
- Aydin, R., M. Calta, D. Sen and M.Z. Coban, 2004. Relationships between fish lengths and otolith length in the population of Chondrostoma regium (Heckel, 1843) inhabiting Keban dam lake. Pak. J. Biol. Sci., 7: 1550-1553.
CrossRefDirect Link - Barkman, R.C., 1978. The use of otolith growth rings to age young Atlantic silversides, Menidia menidia. Trans. Am. Fish. Soc., 107: 790-792.
CrossRef - Begg, G.A., S.E. Campana, A.J. Fowler and I.M. Suthers, 2005. Otolith research and application: Current directions in innovation and implementation. Mar. Freshwater Res., 56: 477-483.
Direct Link - Berg, E., T.H. Sarvas, A. Harbitz, S.E. Fevolden and A.B. Salberg, 2005. Accuracy and precision in stock separation of North-East Arctic and Norwegian coastal cod by otoliths-comparing readings, image analyses and a genetic method. Mar. Freshwater Res., 56: 753-762.
Direct Link - Bolz, G.R. and R.G. Lough, 1988. Growth through the first six months of Atlantic cod, Gadus morhua and haddock, Melanogrammus aeglefinus, based on daily otolith increments. Fish. Bull., 86: 223-236.
Direct Link - Cabral-Solis, E., E. Espino-Barr, M. Gallardo-Cabello and A. Ibanez-Aguirre, 2007. Fishing impact on Mugil curema Stock of multi-species gill net fishery in a tropical lagoon, Colima, Mexico. J. Fish. Aquatic Sci., 2: 235-242.
CrossRefDirect Link - Campana, S.E. and J.D. Neilson, 1985. Microestructure of fish otoliths. Can. J. Fish. Aquat. Sci., 44: 1014-1032.
Direct Link - Carlstrom, D., 1963. A crystallographic study of vertebrate otoliths. Biol. Bull., 125: 441-463.
Direct Link - David, A.W., J.J. Isely and C.B. Grimes, 1994. Differences between the sagitta, lapillus, and asteriscus in estimating age and growth in juvenile red drum, Sciaenops ocellatus. Fish. Bull., 92: 509-515.
Direct Link - Degens, E.T., W.G. Deuser and R.L. Haedrich, 1969. Molecular structure and composition of fish otoliths. Mar. Biol., 2: 105-113.
CrossRef - Eide, A. and A. Wikan, 2010. Optimal selection and effort in a fishery on a stock with cannibalistic behaviour the case of the Northeast arctic cod fisheries. J. Fish. Aquat. Sci., 5: 454-468.
CrossRefDirect Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the Coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Espino-Barr, E., M. Gallardo-Cabello, K. Granados-Flores, E.G. Cabral-Solis, A. Garcia-Boa and M. Puente-Gomez, 2010. Growth analysis of Microlepidotus brevipinnis from the pacific coast of Jalisco, Mexico. J. Fish. Aquat. Sci., 5: 293-303.
CrossRefDirect Link - Francis, C., S.J. Harley, S.E. Campana and P. Doering-Arjes, 2005. Use of otolith weight in length-mediated estimation of proportions at age. Mar. Freshwater Res., 56: 735-743.
Direct Link - Gallardo-Cabello, M., E. Espino-Barr, F. Gonzalez-Orozco and A. Garcia-Boa, 2003. Age determination of Anisotremus interruptus (Perciformes: Haemulidae) by scale reading, in the coast of Colima, Mexico. Rev. Biol. Trop., 51: 519-528.
PubMedDirect Link - Espino-Barr, E., M. Gallardo-Cabello, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2006. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of Caranx caninus (Pisces: Carangidae) in the coast of Colima, Mexico. J. Fish. Aquat. Sci., 1: 157-170.
CrossRefDirect Link - Gallardo-Cabello, M., E. Espino-Barr, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2007. Study of the growth of the green jack Caranx caballus Günther 1868, in the Coast of Colima, Mexico. J. Fish. Aquat. Sci., 2: 131-139.
CrossRefDirect Link - Gallardo-Cabello, M., E. Espino-Barr, R.A. Nava-Ortega, A. Garcia-Boa, E.G. Cabral-Solis and M. Puente-Gomez, 2011. Analysis of the otoliths of sagitta, asteriscus and lapillus of Pacific sierra Scomberomorus sierra (Pisces: Scombridae) in the coast of Colima Mexico. J. Fish. Aquat. Sci., 6: 390-403.
CrossRefDirect Link - Granados-Flores, K., M. Gallardo-Cabello, E. Espino-Barr and E.G. Cabral-Solis, 2010. Age determination of Microlepidotus brevipinnis (Steindachner, 1869) (Pisces: Haemulidae) in the coast of Jalisco, Mexico, by reading otoliths and scales. Int. J. Zool. Res., 6: 1-12.
CrossRefDirect Link - Hickling, C.F., 1931. The structure of the otolith of the hake. Q. J. Microsc. Sci., 74: 547-562.
Direct Link - Ibanez-Aguirre, A.L. and M. Gallardo-Cabello, 1996. Age determination of the grey mullet Mugil cephalus L. and the white mullet Mugil curema V. (Pisces: Mugilidae) in Tamiahua lagoon, Veracruz. Ciencias Marinas, 22: 329-345.
Direct Link - Kamaruzzaman, B.Y., Z. Rina, B.A. John and K.C.A Jalal, 2011. Heavy metals accumulation in commercially important fishes of South West of Malaysian coast. Res. J. Environ. Sci., 5: 595-602.
CrossRefDirect Link - Lagardere, F., 1989. Influence of feeding conditions and temperature on the growth rate and otolith-increment deposition of larval Dover sole (Solea solea) (L.). Rapp. P. Bv. Reun. Cons. Int. Explor. Mer., 191: 390-399.
Direct Link - Authman, M.M.N. and H.H.H. Abbas, 2007. Accumulation and distribution of copper and zinc in both water and some vital tissues of two fish species (Tilapia zillii and Mugil cephalus) of Lake Qarun, Fayoum Province, Egypt. Pak. J. Biol. Sci., 10: 2106-2122.
CrossRefPubMedDirect Link - Mugiya, Y., 1964. Calcification in fish and shell-fish seasonal occurrence of a pre-albumin fraction in the otolith fluid of some fish corresponding to the period of opaque zone formation in their otoliths. Bull. Jap. Soc. Scient. Fish., 30: 445-467.
Direct Link - Ravichandran, S., K. Kumaravel and E.P. Florence, 2011. Nutritive composition of some edible fin fishes. Int. J. Zool. Res., 7: 241-251.
CrossRefDirect Link - Santana-Hernandez, H., E. Espino-Barr, M. Gallardo-Cabello and A. Garcia-Boa, 2008. Morphologic and morphometric analysis and growth rings identification of otoliths: Sagitta, asteriscus and lapillus of yellowfin tuna Thunnus albacares (Bonaterre, 1788) (Pisces: Scombridae) in the Eastern Pacific. Int. J. Zool. Res., 4: 138-151.
CrossRefDirect Link - Shofiquzzoha, A.F.M., M.L. Islam and S.U. Ahmed, 2001. Optimization of stocking rates of shrimp (P. monodon) with brackishwater finfish in a polyculture system. J. Biological Sci., 1: 694-697.
CrossRefDirect Link - Solomon, G., K. Matsushita, M. Shimizu and Y. Nose, 1985. ge and growth of rose bitterling in Shin Tone river. Bull. Jap. Soc. Sci. Fish., 51: 55-62.
Direct Link - Suthers, I.M., K.T. Frank and S.E. Campana, 1989. Spatial comparison of recent growth in postlarval Atlantic cod (Gadus morhua) off southwestern Nova Scotia: Inferior growth in a presumed nursery area. Can. J. Fish. Aquat. Sci., 46: S113-S124.
CrossRefDirect Link - Victor, B.C. and E.B. Brothers, 1982. Age and growth of the fallfish Semotilus corporalis with daily otolith increments as a method of annulus verification. Can. J. Zool., 60: 2543-2550.
CrossRefDirect Link - Holst, E., H. Kaiser, L. Schoen, G. Roebig und G. Goldner, 1950. Otolith functioning in fish. Review of comparative Physiology. (Die Arbeitsweise de Statolithenapparates bei Fischen. Zeitschrift fur Verglaichende Physiologie, 32: 60-120.
CrossRef - Elaine, E.B., C.S.E. Guadalupe, G.C. Manuel and I.A.A. Laura, 2005. Age determination of Mugil curema Valenciennes, 1836 (Pisces: Mugilidae) in the Cuyutlan Lagoon, Colima, Mexico. Int. J. Zool. Res., 1: 21-25.
CrossRefDirect Link