
S.R. Kazemi Nezhad
Department of Genetics, Faculty of Science, Shahid Chamran University of Ahvaz, Ahvaz, Islamic Republic of Iran
E. Modheji
Department of Genetics, Faculty of Science, Shahid Chamran University of Ahvaz, Ahvaz, Islamic Republic of Iran
H. Zolgharnein
Department of Marine Biology, Faculty of Marine Science, Khorramshahr University of Marine Science and Technology, Khorramshahr, Islamic Republic of Iran
Journal of Fisheries and Aquatic Science
Year: 2012 | Volume: 7 | Issue: 5 | Page No.: 339-345







ABSTRACT
In Iranian coastal waters of the Persian Gulf as a result of both direct and indirect human impacts such as uncontrolled or illegal hunting, sea turtle populations are drastically declining. Information on the genetic structure of marine species is essential for stock enhancement programs. To estimate the genetic diversity of the hawksbill turtle populations, the mitochondrial DNA control region was used as a matrilineal marker. The captured turtles (69 samples) were obtained from three different islands in the Persian Gulf. From all the samples DNA was isolated. The primers were selected based on a specific sequence in control region of mitochondrial genome. PCR products were restricted by various restriction enzymes and four different haplotypes observed. Measurement of inter-population genetic diversity and evolutionary distance between genotypes showed a low diversity in mitochondrial genome of hawksbill turtle in the studied regions. Therefore, the similarity between these three populations was significant. Present results provide evidences showing that significant genetic variation was not observed between these distinct populations and there is not even enough evidence to show the separation and diversion in the studied populations in haplotype level.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: February 20, 2012;
Published: March 26, 2012
How to cite this article
S.R. Kazemi Nezhad, E. Modheji and H. Zolgharnein, 2012. Polymorphism Analysis of Mitochondrial DNA Control Region of Hawksbill Turtles (Eretmochelys imbricata) in the Persian Gulf. Journal of Fisheries and Aquatic Science, 7: 339-345.
DOI: 10.3923/jfas.2012.339.345
URL: https://scialert.net/abstract/?doi=jfas.2012.339.345
DOI: 10.3923/jfas.2012.339.345
URL: https://scialert.net/abstract/?doi=jfas.2012.339.345
INTRODUCTION
A successful conservation planning depends on understanding key aspects of hawksbill biology together with the degree of isolation among nesting colonies and migratory pathways of juveniles and adults and also the source for foraging populations. The hawksbill turtle (Eretmochelys imbricata) is a species of endangered sea turtle belonging to the family Cheloniidae (Meylan and Donnelly, 1999). Marine turtles are specific group of vertebrates and creepers who are living in the Atlantic, Pacific and Indian oceans and the warm seas (Venkataraman and Wafar, 2005). There are two species of marine turtles in the Persian Gulf that include the green and hawksbill turtles (Mobaraki, 2004a; Ross and Barwani, 1995). According to the IUCN report (Cheraghi et al., 2008), the first species is at risk of extinction and the second one is in critical condition (Baillie and Groombridge, 1996).
Molecular markers such as mitochondrial DNA (mtDNA) show uniparental inheritance which approximately bears the lack of recombination so they can be used as a beneficial tool to study phylogeographic (Norazila and Patimah, 2002) and speciation (Sanna et al., 2008). All organisms due to cellular reaction or interaction with the environment are prone to different mutations which lead to polymorphism and finally genetic diversity (Gharibkhani et al., 2009). Two important environmental factors (Rahman et al., 2008) include natural selection and genetic drift in one individual or between different individuals or higher level of organization stabilizes the genetic diversity (Atashin et al., 2010). Some differences are usually detected in the nucleotide sequence of one gene or a specific part of a gene which create intra or inter species variation (Sultana and Khan, 2007). Although these variations are distinguishable by two methods based on molecular study, in case of first method sequences fragments are directly identified and the other method is based on PCR-RFLP method which elucidate the variations indirectly (Davis and Sterrett, 2011). The provided data from analysis of mtDNA sequencing and RFLP can be used in estimation of each haplotype divergence time and to elucidate the ancient division of different species (Heist et al., 1995, 1996).
Main species of marine turtles existing in Iran is Eretmochelys imbricata and their main habitat is the Persian Gulf (Mobaraki, 2004b). Although precise census for identifying the number of sea turtles in Qeshm, Kish and Nakhiloo has not been performed yet but according to the IUCN reports in respect to the Persian Gulf pollution, this species is prone to extinction due to mortal environmental effects. Therefore, to study and identify this species, we need more information about gene pool of sea turtle species. The purpose of this research is first, to study the different population of Eretmochelys imbricata and compare different habitat based on dominant population in each environment and second, nomination and comparison between various population in Iran and study of their genetics to detect the genetic structure of species based on PCR-RFLP using mtDNA samples (Fayazi et al., 2006).
In the present study, we have tested the similarity hypothesis in three distinct islands (Kish, Qeshm and Nakhiloo) in the Persian Gulf (Zolgharnein et al., 2010). Kish (26 32 N 53 58 E) is located in the Persian Gulf 19 km from mainland Iran and has an area of around 91 km2 with an outer boundary of 40 km and a nearly elliptical shape. The Island is positioned along the 1359 km long Iranian coastline north of the Persian Gulf. Qeshm Island (26 45 N 55 49 E) is located a few kilometers off the southern coast of Iran (Persian Gulf), opposite the port cities of Bandar-Abbas and Bandar Khamir. The island which hosts a 300 km2 free zone jurisdiction, is 135 km long and lies strategically in the Strait of Hormuz. Nakhiloo Island (27 49 N 51 28 E) located in the southern coast of the province is devoid of inhabitants. The Nakhiloo Island is the area where giant tortoises inhabit, the presences which have made this area environmentally important (Zolgharnein et al., 2011). The object of the study was to findout the heterogeniety between different population in haplotype level.
MATERIALS AND METHODS
In this study, sampling was performed in the end of spring season (2010) by digging the marked nests where turtles put their eggs and some random samples were taken from each hole. Tissue specimens were obtained from their beach nests at three locations (Qeshm Island, #40; Kish Island, #25 and Nakhiloo Island, #4) in the Persian Gulf. Embryos of each nest were collected separately and fixed immediately in absolute ethanol onsite and later frozen (-18°C), until further use. Total genomic DNA from 69 specimens was extracted using phenol-chloroform method (Alam et al., 1996). Finally, DNA was dissolved in 100 μL of double distilled water and stored at -20°C. PCR primers (TAG Copenhagen, Denmark) were designed based on a sequence of control region in D-loop of mitochondrial genome, each primer had 20 bp nucleotide length and their sequences were 5'-GTCTCGGATTTAGGGGTTTG-3' and 5'-GCTTAACCCTAAAGCATTGG-3' for LCM15382 (Diaz-Fernandez et al., 1999) and H950 (Laurent et al., 1998) , respectively. Amplified fragments were restricted by seven (EcoRI, HindIII, HinfI, HindII, AfaI, BamHI and NdeI) different restriction enzymes (Fermentas GmbH, Germany) then the samples along with 50 bp ladders loaded on Polyacrylamide gel electrophoresis (Bio-Rad, Germany).
Statistical analysis: To test the significance of hypothesis and to estimate the diversity between three studied populations, chi-square test was used and likelihood ratio for these populations calculated (Eyduran et al., 2006). According to the test result (p<0.05), enough evidence was not found to eliminate the hypothesis. The estimated G2 elucidate the similarity between these populations. In addition to the number of genotypes (na) and gene diversity (h) for each population, we also tried to calculate total na and h in all samples simultaneously. Another factor which we calculated was genetic distance to identify the level of similarity between three different populations (Bjorndal et al., 2005).
RESULTS
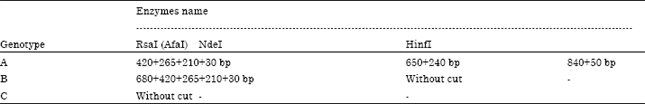
We analyzed sixty nine samples of unknown haplotypes in order to determine if RFLP analysis could be used to discriminate haplotypes in a blind test or not. Four different haplotypes were detected by seven restriction enzymes cut sites (Table 1). Digestion of the 890 bp segment PCR product with restriction endonucleases showed diverse genotypes. Three different genotypes were observed based on number and size of fragments created by restriction enzymes (Table 2). First enzyme was NdeI which produced two genotypes, one of them showed two electrophoretic bands and was called genotype A which was the dominant genotype in this population. Another genotype showed only one band since there was no cut in DNA and was named genotype B. The restriction of DNA samples by AfaI enzyme produced three different genotypes which were called A, B and C. The restricted DNA samples by HinfI produced only genotype A.
| Table 1: | Different types of genotypes which are produced due to restriction of PCR products by seven various Restriction enzymes |
 | |
| Table 2: | The size of DNA fragments in each genotype after cutting by enzymes |
 | |
| Table 3: | The calculation of χ2 test values based on Restriction enzyme |
 | |
| Table 4: | The estimation of Similarity ratio (G2) between three populations |
 | |
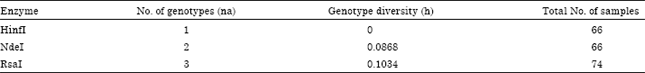
| Table 5: | The estimation of na and h for all samples based on Restriction enzymes |
 | |
| Table 6: | Matrix of Similarity and Genetic distance between three populations based on restriction of PCR products by HinfI, NdeI, AfaI enzymes |
 | |
The results of cutting by HindII, EcoRI, BamHI and HindIII were similar and only genotype A was observed in all cases. RFLP analysis from two polymorphic enzymes, three and four restriction and non restriction enzymes , respectively, shows four different genotypes in studied populations. The frequency of one haplotype from 4 haplotypes was only 1, so it is called "rare haplotype". Although the less amount of variation was observed in these populations but most of the variation belonged to Qeshm Island. Comparison between the obtained results due to cutting by three enzymes of NdeI, AfaI (RsaI), HinfI shows that HinfI enzyme is the best proof of geographical similarity between these three populations. The highest level of differences in genotype frequency was observed in Kish and Qeshm populations. According to the χ2 test, we did not have significant evidence to eliminate the similarity hypothesis (Table 3). Estimated similarity ratio (G2) elucidated the similarity between these populations (Table 4). Gene diversity was calculated and indicated the highest level of Gene diversity (h) present in Qeshm populations while DNA was restricted using AfaI enzyme. The calculated total "na" (number of genotypes) and "h" reveals that the highest and lowest amount of similarity ratio belonged to AfaI and HindI enzymes which are equal to 0.1034 and 0.0, respectively (Table 5). Moreover, we calculated standard genetic distance (Ds) based on the level of genetic divergence between different populations, the highest and lowest level of similarity between three populations observed when DNA samples were restricted using HinfI and AfaI (RsaI) enzymes, respectively (Table 6). Actually, the highest genetic distance was very less (equal to 0.0035) so it can prove that the similarity between these populations is significantly high.
DISCUSSION
The sensitivity of the RFLP technique was demonstrated by coordination between banding patterns and nucleotide sequence haplotypes assigned for sixty nine samples of three populations tested (Rozihan and Ismail, 2011). We were able to identify a dominant haplotype pattern (haplotye A) which can be registered as a new haplotype of the hawksbill turtles in the Persian Gulf. According to the findings, dominant haplotype dispersions in these three different populations were approximately equal and the less number of haplotypes show that the variation in mitochondrial genome of Eretmochelys imbricata is low in studied populations. In studied haplotypes, one rare haplotype was observed with frequency equal to 1, it could be possible that this rare haplotype was a dominant haplotype in the past and its frequency has decreased or it might be a new haplotype which is produced in the population and may increase due to life condition over times or perhaps they are produced by new mutation or crossing over phenomenon. Finally, this study proved that the level of genetic variation in these populations was very less and it seems that there is a significant genetic relationship between these islands population. These results show that the PCR-RFLP method applied to the analysis of sea turtle mtDNA control region is efficient in distinguishing the species in the Cheloniidae family and its haplotypes. This simple and fast technique could also be employed to reduce the sequencing efforts (Kaska, 2000; Kavan et al., 2009).
Among these seven enzymes only three of them restricted PCR products which included AfaI, NdeI and HinfI. Among these three restriction enzymes, AfaI had the highest amount of restriction sites. The highest level of genetic distances were detected between Qeshm and Kish populations (GD = 0.0035) and also between Qeshm and Nakhiloo (GD = 0.0035) using AfaI enzymes although the highest level of genetic distance was observed between Kish and Nakhiloo population (GD = 0.0007) using NdeI enzyme. Four different haplotypes with increasing the frequencies include CAA, ABA, BAA, AAA , respectively, were produced by these three enzymes. Among these haplotypes, a rare haplotype with frequency 1.00 has been seen. In addition, the results showed no considerable heterogeneity between different populations and there are no enough evidences at least in haplotypic level for separation and distinction of studied samples.
ACKNOWLEDGMENTS
This study was supported by grants from the research council of Shahid Chamran University of Ahvaz and Khorramshahr University of Marine Science and Technology. We also appreciate the Hormozgan provincial office of Iranian department of environment for their support. The authors declare that they have not conflicts of interest.
REFERENCES
- Alam, S., A. Popplewell and N. Maclean, 1996. Germ line transmission and expression of a lacZ containing transgene in tilapia (Oreochromis niloticus). Transgenic Res., 55: 87-95.
Direct Link - Atashin, S., M. Pakshir and A. Yazdani, 2010. Comparative studying of marine parameters' effect, via qualitative method. Trends Applied Sci. Res., 5: 120-128.
CrossRefDirect Link - Bjorndal, KA., AB. Bolten and S. Troeng, 2005. Population structure and genetic diversity in green turtles nesting at Tortuguero, Costa Rica, based on mitochondrial DNA control region sequences. Mar. Biol., 147: 1449-1457.
Direct Link - Cheraghi, M., N. Khorasani, M. Karami, M. Shariat and B. Riazi 2008. The conformity of Iran`s protected areas with IUCN categorization system. J. Boil. Sci., 8: 441-445.
CrossRefDirect Link - Davis, A.K. and S.C. Sterrett, 2011. Prevalence of haemogregarine parasites in three freshwater turtle species in a population in Northeast Georgia, USA. Int. J. Zool. Res., 7: 156-163.
CrossRef - Diaz-Fernandez, R., T. Okayama, C.E. Uchiyama, G. Espinosa, R. Marquez, C. Diez, and H. Koike, 1999. Genetic sourcing for the hawksbill turtle, Eretmochelys imbricata, in the Northern Caribbean region. Chelonian. Conserv. Biol., 3: 296-300.
Direct Link - Eyduran, E., T. Ozdemir, M. Kazim Kara, S. Keskin and B. Cak, 2006. A study on power of chi-square and g statistics in biology sciences. Pak. J. Biol. Sci., 9: 1324-1327.
CrossRefDirect Link - Fayazi, J., M. Moradi, G. Rahimi, R. Ashtyani and H. Galledari, 2006. Genetic differentiation and phylogenetic relationships among Barbus xanthopterus (Cyprinidae) populations in Southwest of Iran using mitochondrial DNA markers. Pak. J. Biol. Sci., 9: 2249-2254.
CrossRefDirect Link - Gharibkhani, M., M. Pourkazemi, M. Soltani, S. Rezvani and L. Azizzadeh, 2009. Population genetic structure of pikeperch (Sander lucioperca Linnaeus, 1758) in the Southwest Caspian Sea using microsatellite markers. J. Fish. Aquatic Sci., 4: 161-168.
CrossRef - Heist, E.J., J.E. Graves and J.A. Musick, 1995. Population genetics of the sandbar shark (Carcharhinus plumbeus) in the Gulf of Mexico and mid-Atlantic Bight. Copeia, 3: 555-562.
Direct Link - Heist, E.J., J.A. Musick and J.E. Graves, 1996. Mitochondrial DNA diversity and divergence among sharpnose sharks, Rhizoprionodon terraenovae, from the Gulf of Mexico and Mid-Atlantic bight. Fishery Bull., 94: 664-668.
Direct Link - Kavan, L.S. S.R. Gilkolaei, G. Vossoughi, S.M.R. Fatemi, R. Safari and S. Jamili, 2009. Population genetic study of Rutilus frisii kutum (Kamansky 1901) from the Caspian Sea; Iran and Azerbaijan regions, using microsatellite markers. J. Fish. Aquat. Sci., 4: 316-322.
CrossRefDirect Link - Laurent, L., P. Casale, M.N. Bradai, B.J. Godley and G. Gerosas et al., 1998. Molecular resolution of marine turtle stock composition in fishery bycatch: A case study in the Mediterranean. Mol. Ecol., 7: 1529-1542.
CrossRefPubMedDirect Link - Norazila, K.S. and I. Patimah, 2002. Mitochondrial 16S and 12S rRNA/tRNA-Val gene analysis in tiger barbs (Puntius tetrazona). J. Biol. Sci., 2: 754-756.
CrossRefDirect Link - Rahman, K.S.M. and E. Gakpe, 2008. Production, characterisation and applications of biosurfactants-review. Biotechnology, 7: 360-370.
Direct Link - Rozihan, M. and E. Ismail, 2011. Genetic structure and haplotype diversity of Tachypleus gigas population along the West Coast of peninsular Malaysia-inferred through mtDNA AT rich region sequence analysis. Biotechnology.
CrossRefDirect Link - Sanna, D., A. Addis, F. Biagi, C. Motzo, M. Carcupino and P. Francalacci, 2008. mtDNA control region and D-HPLC analysis: A method to evaluate the mating system in Syngnathidae (Teleostei). Mar. Biol., 153: 269-275.
Direct Link - Sultana, G.N.N. and A.H. Khan, 2007. Optimization of the sample preparation method for DNA sequencing. J. Boil. Sci., 7: 194-199.
CrossRefDirect Link - Zolgharnein, H., M. Kamyab, S. Keyvanshokooh, A. Ghasemi and S.M.B. Nabavi, 2010. Genetic diversity of Avicennia marina (Forsk.) Vierh. populations in the persian gulf by microsatellite markers. J. Fish. Aquat. Sci., 5: 223-229.
CrossRefDirect Link - Zolgharnein, H., M.A. Salari-Aliabadi, A.M. Forougmand and S. Roshani, 2011. Genetic population structure of Hawksbill turtle (Eretmochelys imbricta) using microsatellite analysis. Iranian J. Biotechnol., 9: 56-62.
Direct Link - Mobaraki, A., 2004. Nesting of the hawksbill turtle at Shidvar Island, Hormozgan province, Iran. Marine Turtle Newslett., 103: 13-13.
Direct Link