
S. Salcedo-Bojorquez
Centro Interdisciplinario de Ciencias Marinas del IPN, Apartado Postal 592, La Paz, 23096, Baja California Sur, Mexico
F. Arreguin-Sanchez
Centro Interdisciplinario de Ciencias Marinas del IPN, Apartado Postal 592, La Paz, 23096, Baja California Sur, Mexico
Journal of Fisheries and Aquatic Science
Year: 2011 | Volume: 6 | Issue: 6 | Page No.: 578-591







ABSTRACT
This contribution is aimed to the identification of reproductive patterns of billfishes, which is in turn a key issue supporting management and conservation policies. Information reported in the literature on features of reproductive processes of billfishes was examined with a Principal Components Analysis (PCA) to identify groups of species with similar reproductive strategies. PCA was applied based in a correlation matrix and the significances (p<0.05) of the factors and loadings of the variables were used as criteria for the identification of reproductive patterns. In general, four groups were found. The first group consists of Tetrapturus albidus and T. audax, which are characterised by rapid growth, an early age at first maturity with respect to maximum age (T/Tmax) and high values of reproductive variables (e.g., Spawning Duration (Sd), Spawning Frequency (Sf), Annual Fecundity (AF) and Relative Fecundity (RF). A second group consists of T. belone, T. georgii and T. pfluegeri, which present rapid growth, high Tm/Tmax ratios and Batch Fecundity (BF) values and low Sd values. Another group consists of the genera Makaira and Xiphias, which present low Lm/Lmax ratios and high values for AF, RF, Sf and BF. The last group consists of species of the genus Istiophorus, which present the lowest AF values and the highest RF and BF values. In billfishes, the reproductive pattern is strongly influenced by the growth process and the age at which a species reaches sexual maturity relative to its maximum age. The observed differences in reproductive variables result from adaptive mechanisms that allow species greater reproductive success in the epipelagic environment.
PDF Abstract XML References Citation
Received: March 23, 2011;
Accepted: May 18, 2011;
Published: August 13, 2011
How to cite this article
S. Salcedo-Bojorquez and F. Arreguin-Sanchez, 2011. An Exploratory Analysis to Identify Reproductive Strategies of Billfishes. Journal of Fisheries and Aquatic Science, 6: 578-591.
DOI: 10.3923/jfas.2011.578.591
URL: https://scialert.net/abstract/?doi=jfas.2011.578.591
DOI: 10.3923/jfas.2011.578.591
URL: https://scialert.net/abstract/?doi=jfas.2011.578.591
INTRODUCTION
Among the important parameters in reproductive biology are the breeding season and area, the age at maturity, the age at first reproduction and fecundity (Jakobsen et al., 2009). One of the most important aspects for the sustainable use and management of living marine resources is the maintenance of vital rates that will produce the highest levels of resilience. The vital rates are of great importance to reproductive success, which depends on both fishing yields and population maintenance. In this context, understanding the parameters that characterise reproductive processes can help managers determine, for example, the effects of fishing on the renewal potential of stocks (Mace, 1993; Murawski et al., 2001). Investigating these parameters has allowed the characterisation of reproductive strategies, which in turn correspond to species’ adaptations to environmental conditions (Maury and Lehodey, 2005).
Comparative studies of these parameters have been widely recognised; however, for some fish, such as billfishes, studying reproductive parameters is a complex process because the fish migrate and there is insufficient knowledge of breeding areas (Richardson et al., 2009a, b). The management of these species is challenging partly because of their migratory nature, which hinders the study of their life histories, habitat requirements and stock sizes (ICCAT, 2006) and causes the fishes to cross jurisdictional boundaries.
Studies on reproductive processes of billfishes have mainly focused on two or three species (e.g., Xhiphiias gladius andTetrapturus audax), although most species are targeted for commercial and recreational fisheries around the world (Fromentin and Fonteneau, 2001; Brinson et al., 2009). Moreover, most knowledge of reproductive aspects is limited to certain regions and time periods (ICCAT, 2004) and little is known on a global scale.
In general, it is recognised that distributional differences (tropical or temperate) correspond to differences in characteristics of these species and their responses to fishing pressure (Pauly, 1998; Fromentin and Fonteneau, 2001; Boyce et al., 2008). We hypothesis that different reproductive strategies among billfishes can be identified by examining combinations of reproductive characteristics with their variations. Thus, the aim of this study was to synthesise available information on reproduction of these species to identify patterns in the reproductive process.
MATERIALS AND METHODS
We collected information on reproductive life histories of species in the fish families Xiphiidae and Istiophoridae using literature from databases, such as ASFA (Aquatic Sciences and Fisheries Abstracts), EBSCOhost (http://www.ebscohost.com/), FishBase (www.FishBase.org.) and grey literature (thesis dissertations, technical reports). Key parameters that characterise the reproductive process are listed below. In this list, age is measured in years and length is defined as the Lower Jaw-Fork Length (LJFL) and is measured in centimetres unless otherwise specified.
| • | Length (Lmax) and maximum age (Tmax) |
| • | Length (Lm) and age (Tm) at first maturity (at which 50% of the individuals are mature and able to reproduce) |
| • | Duration of the spawning period (Sd) (the number of months per year during which spawning occurs, measured in days) |
| • | Spawning frequency (Sf) (number of spawns per month, measured in days) |
| • | Relative Fecundity (RF) (number ova/grams of body weight) |
| • | Batch Fecundity (BF) (number of ova) |
| • | Annual Fecundity (AF) (i.e., the product of batch fecundity, spawning frequency and the duration of the spawning season) |
Because some species are sexually dimorphic (e.g., X. gladius and Makaira spp.), certain features were reported separately for each sex in the literature; in these cases we used the data on females. Because most researchers did not report all of the parameters listed above, we also considered additional information to estimate missing data. For example, we used the parameters of the Von Bertalanffy growth equation (Von-Bertalanffy, 1938), K (y-1), L8 (cm) and t0 (y-1), or estimate some of them when needed (Pauly, 1980; Froese and Binohlan, 2000; Froese and Pauly, 2000). Also, in some cases, length and maximum age were used to estimate the length and age at first maturity (Froese and Binohlan, 2000). Other empirical equations were also used to calculate total length, total weight, eye-fork length and gutted weight (Wares and Sakawa, 1974; Prince, 1990; Prager et al., 1995; IOTC, 2005).
Statistical analysis: We used a Principal Component Analysis (PCA) based on a correlation matrix to analyse the reproductive data for different species. The statistical significance of the components and the contribution of each variable in each component (i.e., loading) were computed following a procedure that is based on the rotation of the normalised variance method (Kaiser, 1960). In our case, loading values ≥0.7 were considered significant.
RESULTS
The life history parameters considered for our analysis are shown in Table 1. From the eleven species analysed, only three (X. gladius, T. audax and I. platypterus) were described by all of the reproductive variables, while values for some variables were not available for Tetraturus belone, T. pfluegeri, T. georgii and T. angustirostris. Other species account for some variables.
We did a preliminary analysis to explore how variables were related and to identify whether a significant correlation could bias the results of the PCA. Table 2 shows the significant correlations (p<0.05); correlations between Wm and Lmax, Wmax, Tmax and BF, as well as between BF and Lmax, Wmax, Tmax and Wm showed evidence some autocorrelation through Wm. Thus, this variable was eliminated from the analysis.
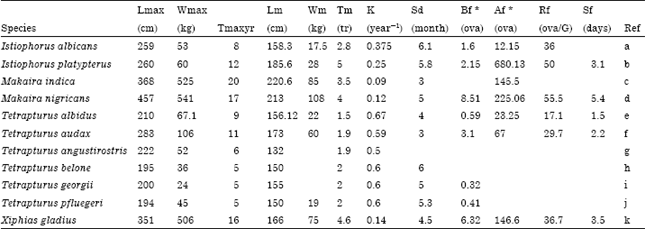
| Table 2: | Correlation coefficients between variables considered for PCA |
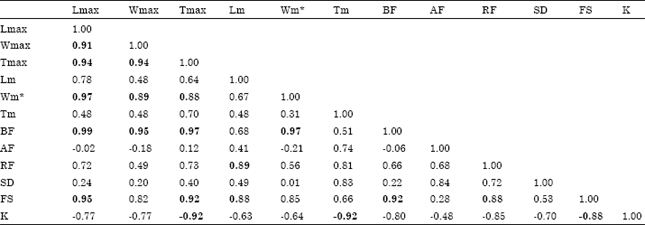 | |
| Significant correlations (p<0.05) are in bold. Lmax: Maximum length (cm); Wmax: Maximum weight (g); Tmax: Maximum age (year); Lm: Length (cm) at first maturity; Wm: Weight at first maturity; Tm: Age (year) at first maturity; K (y-1): Von Bertalanffy growth coefficient; Sd: Spawning duration (month); Bf: Batch fecundity (ova/spawning); Af: Annual fecundity (ova); Rf: Relative fecundity (ova/g); Sf: Spawning frequency (days). *Variable not used for the final analysis (see text for explanation) | |
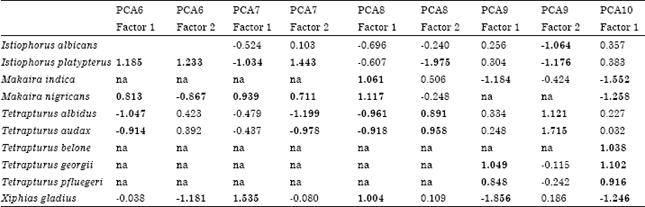
| Table 3: | Results of the primary principal components analysis (PCA) of variables associated with reproductive processes of billfishes. PCA# indicates different scenarios of PCA |
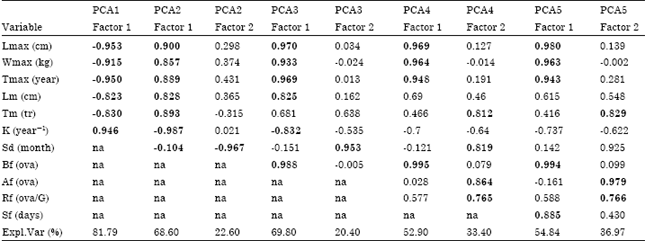 | |
| Significant (p<0.05) components (factors) and variables within each factor (loadings>0.7) are indicated in bold. Values for some variable are not included in the analysis because there were no data (na = not applicable). Expl.Var. refers to the variance explained by each factor | |
Other significant correlations did not show clear evidence of functional relationships or patterns that could be maintained across species. Because there was not enough evidence to eliminate other variables, we made a first approximation of the PCA with all variables except Wm.
Given that a PCA requires complete data sets and that our data set was incomplete for some species, it was not possible to incorporate all of the parameters into a single analysis. Therefore, different species combinations were used to explore the different variables and species. Table 3 shows the combinations and the main results.
Present initial analysis produced two main findings. The variables in the first component, Lmax Wmax, Tmax and, except in two cases, Lm, were consistently significant. Also, species tended to fall into three groups, corresponding to the genus: Istiophorus, the complex Makaira-Xiphias and a third group called Tetrapturus1 (which includes species T. albidus and T. audax). Significant variables resulting from the PCA, in conjunction with the significant correlations in Table 2, suggest covariance or functional relationships between some of the variables. For this reason, the PCA was redefined with the following variables: Lm/Lmax, Wmax, K, Sd, BF, AF, RF and Sf. Lm/Lmax is an index that relates the size at which a fish reaches maturity to the maximum size of the fish; it is an expression that, from a physiological point of view, is equivalent to Tm/Tmax. The species Tetrapturus angustirostris was not included in the PCA because of the lack of reproductive data for these parameters.
Results of the five PCA scenarios are shown in Table 4a and b. In general, groups similar to those described above were identified and, when other species of Tetrapturus were incorporated into the analysis (scenarios PCA9 and PCA10), they formed a second group we named Tetrapturus2 (Fig. 1).
| Table 4a: | Results of the principal components analysis (PCA) applied to variables associated with reproductive processes of billfishes. PCA# indicates different applications of PCA |
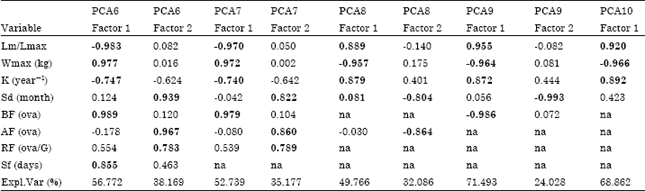 | |
| Significant (p<0.05) components (factors) and variables within each factor (loadings>0.7) are indicated in bold.Values for some variable are not included in the analysis because there were no data (na = not applicable). Expl.Var. refers to the variance explained by each factor. K: Von Bertalanffy growth coefficient; Sd: Spawning duration; BF: Batch fecundity; AF: Annual fecundity; RF: Relative fecundity; Sf: Spawning frequency | |
| Table 4b: | Results of the principal components analysis (PCA) applied to variables associated with reproductive processes of billfishes. PCA# indicates different applications of PCA |
 | |
| Significant (p<0.05) components (factors) and species within each factor (loadings>0.7) are indicated in bold. Values for some variable are not included in the analysis because there were no data (na = not applicable). Expl. Var. refers to the variance explained by each factor | |
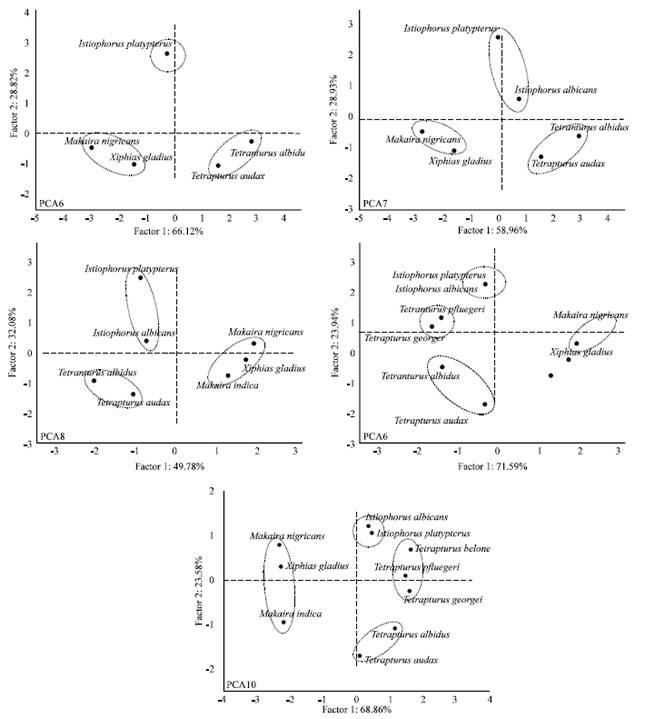 | |
| Fig. 1: | Graphs of five PCA scenarias |
With respect to the significant variables, there are two types: those associated with individual development (i.e., K, Wmax and Lm/Lmax), which were significant in the first component in almost all PCA scenarios and those directly expressing an attribute of the reproductive process. For the latter, two variables represent the number of eggs produced (RF and BF) and three variables represent reproductive rates (Sd, AF, Sf). Except for Sf in the PCA6 scenario, these rates were consistently significant in the second component.
Results from the PCA suggest the presence of four groups with clearly identifiable reproductive characteristics (Fig. 1):
Tetrapturus 1: This group is represented by the species T. albidus and T. audax. These species are generally associated with the lowest values of the reproductive variables (i.e., Sd, Sf, AF and RF). The low Tm/Tmax ratios (<20%) and the high values of the growth coefficient, K, suggest rapid growth and early maturity.
Tetrapturus 2: This group is represented by the species T. belone, T. georgii and T. pfluegeri. While these species are fast growing, they reach maturity at old ages (Tm/Tmax = 40%). They also are small in size and weight and have a longer spawning duration (Sd) and lower batch fecundity (BF).
Makaira-Xiphias: This group consists of species of the genera Makaira and Xiphias. Among billfish, the species in this group have the lowest Lm/Lmax ratios (50-60%) and they have relatively low Tm/Tmax ratios; these data suggest that the fish in this group mature at relatively small sizes even when they have a slow growth rate. These species also tend to have a high Spawning Frequency (SF) and the highest values of annual, relative and batch fecundities (AF, RF and BF, respectively).
Istiophorus: This group consists of species of the genus Istiophorus, specifically I. albicans and I. platypterus. They present intermediate values of the growth coefficients and of the Lm/Lmax ratio. However, they have the lowest values of AF and relatively high values of Sd, RF and BF.
DISCUSSION
Females of most billfishes species reach sexual maturity at larger sizes than do males (Nakamura, 1985); this pattern is the only type of "sexual dimorphism" that is non-apparent, although it has been reported that some species, such as T. pfluegeri, T. belone and T. angustirostris, do not show such differences. The first two classified within the group Tetrapturus 2 (the third one was not into the analysis because absence of data).
Most studies reported that spawning duration time is approximately 3 to 6 months and, for some species, such as I. platypterus and I. albicans, there are two reproductive periods. De Sylva and Breder (1997) indicated that males can reproduce year round. Spawning frequency (Sf) data indicate that billfishes lay eggs between 1.5 to 5.4 days.
M. nigricans has a Sf that ranges from 2.4 to 8.3 days and is therefore the species with the greatest range of Sf values during the breeding season. Also, this species has the highest BF and is the largest in length and weight. Relatively larger females are physiologically capable of maintaining higher reproductive frequencies, so this seems to be related to their energy reserve (Schaefer, 2003). The variability in BF can change from year to year for similarly sized individuals that are of the same species and in the same area (Schaefer, 1998). Some factors, such as food, growth rate and population density, influence this variability (Tamate and Maekawa, 2000; Blanck and Lamouroux, 2006).
By observing the distinctive characteristics of each group (Table 5), it is clear that growth is strongly associated with the reproductive processes of these species. In particular, growth is related to how fast a female reaches sexual maturity and, in this context, growth is associated with the significant variables Lm and Lmax.
| Table 5: | Relationships between variables associated with the reproductive processes of billfishes |
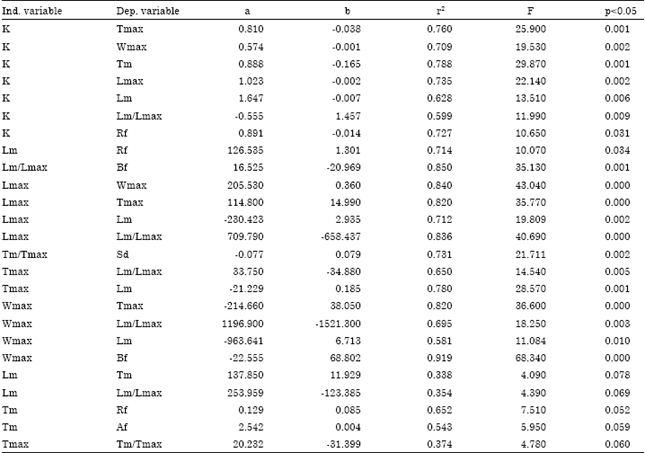 | |
| Significance was set at p<0.05, with the exception of the last five relationships, which were significant at p<0.10 (ind.=independent; dep.=dependent). K (y-1): Von Bertalanffy growth coefficient; Lm: Length (cm) at first maturity; Lmax: Maximum length (cm); Tm: Age (year) at first maturity; Tmax: Maximum age (year); Wmax: Maximum weight (g); Af: Annual fecundity (ova); Rf: Relative fecundity (ova/g); Sd: Spawning duration (dais); Bf: Batch fecundity (ova/spawning). Note that spawning frequency, Sd, was not significantly correlated with any variable. a and b are regression parameters, r2 is determination coefficient expressing how much variance of dependent variable is explained by independent variable, F corresponded to F test for significance | |
Thus, the Relative Fecundity (RF) is significantly correlated with K, Lm and Tm; the Batch Fecundity (BF) is correlated with Wmax and the Lm/Lmax ratio; the Annual Fecundity (AF) is correlated Tm; and the spawning duration (Sd) is correlated with the Tm/Tmax ratio.
Billfish are considered epipelagic species (Block et al., 1992; Boggs, 1992) that occupy high trophic levels >5 when they are adults (Griffiths et al., 2010). Usually, billfishes occupy the same habitat and their distribution often overlap spatially (Campbell and Young, 2008; Shimose et al., 2010; Young et al., 2010). However, each species tolerates different ranges of environmental variation (Nakamura, 1985; Bigelow et al., 1999). Their potential prey may also be similar (Shimose et al., 2010) but may vary between seasons, years, areas and target preys. This variation may reduce competition for food (Young et al., 2010) and thus may be an adaptive strategy that allows billfishes to exploit the same environment (Dagorn et al., 2000).
All billfish species are partial or multiple spawners with asynchronous development of oocytes (De Sylva and Breder, 1997; Hernandez-Herrera et al., 2000; Murua and Saborido-Rey, 2003; Arocha et al., 2005; Kopf et al., 2009). Present a pattern with an indeterminate annual fecundity and therefore the breeding season is prolonged, characteristic of species with this reproductive pattern (Brown-Peterson et al., 1988). According to Lambert and Ware (1984), partial spawning may be a strategy to release eggs over a long period of time, thus increasing the probability of progeny survival.
According to Murua et al. (2003), variation in reproductive characteristics of a species is a function of or a response to, variation in environmental conditions, such as temperature, food availability, habitat and predation intensity; however, the migratory nature of these species allows them to exploit the resources of their habitats to improve their reproductive success (Wootton, 1990).
Reproductive strategies reflect marked differences between species (Wootton, 1984; Murua and Saborido-Rey, 2003), even for billfish species, which present similar processes that govern their reproductive strategies. It is clear, however, that these species have different reproductive strategies due to adaptive mechanisms that allow the fish greater reproductive success. This conclusion is evidenced by our analysis, which identified four groups with distinct reproductive strategies.
CONCLUSION
The major conclusion of our research finding is that billfish present four different reproductive strategies that can be identified based on their growth and how quickly they reach the size or age at first maturity, as well as a differential combination of reproductive characteristics related to fecundity, the frequency and duration of spawning.
ACKNOWLEDGMENTS
The first author thanks CONACyT for Scholarship # 176948. The authors also thank CONACyT for their support through projects SEP-CONACyT (104974) and ANR-CONACyT (111465), as well as the National Polytechnic Institute through the grants SIP-20100404, SIP-20113417, EDI and COFAA.
REFERENCES
- Arocha, F., A. Barrios, J. Silva and D.W. Lee, 2005. Preliminary observations on gonad development, sexual maturity and fecundity estimates of White marlin (Tetrapturus albidus) from the western central Atlantic. Col. Vol. Sci. Pap. ICCAT, 58: 1567-1573.
Direct Link - Von Bertalanffy, L., 1938. A quantitative theory of organic growth (Inquiries on growth laws. II). Hum. Biol., 10: 181-213.
Direct Link - Bigelow, K., J. Hampton and N. Miyabe, 1999. Effective longline effort within the yellowfin habitat and standardized CPUE. Proceedings of the 12th Meeting of the Standing Committee on Tuna and Billfish, June 16-23, Japan, pp: 10-10.
Direct Link - Blanck, A. and N. Lamouroux, 2006. Large-scale intraespecific variation in life-history traits of European freshwater fish. J. Biogeogr., 34: 862-875.
CrossRef - Block, B.A., D.T. Booth and F.G. Carey, 1992. Depth and temperature of the blue marlin, Makaira nigricans, observed by acoustic telemetry. Mar. Biol., 114: 175-183.
CrossRef - Boggs, C.H., 1992. Depth, capture time and hooked longevity of longline-caught pelagic fish-timing bites of fish with chips. Fish Bull., 90: 642-658.
Direct Link - Boyce, D.G., D.P. Tittenson and B. Worm, 2008. Effects of temperature on global patterns of tuna and billfish richness. Mar. Ecol. Prog. Ser., 355: 267-276.
CrossRefDirect Link - Brinson, A.A., D.J. Die, P. Bannerman and Y. Diatta, 2009. Socioeconomic performance of West AFrican fleets that target Atlantic billfish. Fish. Res., 99: 55-62.
CrossRef - Dagorn, L., F. Menczer, P. Bach and R.J. Olson, 2000. Co-evolution of movement behaviours by tropical pelagic predatory fishes in responseto prey environment: A simulation model. Ecol. Modell., 134: 325-341.
CrossRef - De Sylva, D.P. and P.R. Breder, 1997. Reproduction, gonad histology and spawning cycles of North Atlantic billfishes (Istiophoridae). Bull. Mar. Sci., 60: 668-697.
Direct Link - Froese, R. and C. Binohlan, 2000. Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. J. Fish Biol., 56: 758-773.
CrossRef - Fromentin, J.M. and A. Fonteneau, 2001. Fishing effects and life history traits: A case study comparing tropical versus temperate tunas. Fish. Res., 53: 133-150.
CrossRef - Griffiths, S.P., J.W. Young, M.J. Lansdell, R.A. Campbell and J. Hampton et al., 2010. Ecological effects of longline fishing and climate change on the pelagic ecosystem off eastern Australia. Rev. Fish Biol. Fish., 20: 239-272.
CrossRef - Hernandez-Herrera, A., M. Ramirez-Rodriguez and A. Muhlia-Melo, 2000. Batch fecundity and spawning frequency of sailfish (Istiophorus platypterus) off the Pacific coast of Mexico. Pac. Sci., 54: 189-194.
Direct Link - Kaiser, H.F., 1960. The application of electronic computers to factor analysis. Educ. Psychol. Measur., 20: 141-151.
CrossRef - Murawski, S.A., P.J. Rago and E.A. Trippel, 2001. Impacts of demographic variation in spawning characteristics on reference points for fishery management. ICES J. Mar. Sci., 58: 1002-1014.
CrossRef - Murua, H. and F. Saborido-Rey, 2003. Female reproductive strategies of marine fish species of the North Atlantic. J. Northwest Atlantic Fish. Sci., 33: 23-31.
Direct Link - Murua, H., G. Kraus, F. Saborido-Rey, P.R. Witthames, A. Thorsen and S. Junquera, 2003. Procedures to estimate fecundity of marine fish species in relation to their reproductive strategy. J. Northwest Atlantic Fish. Sci., 33: 33-54.
Direct Link - Pauly, D., 1980. On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 Fish stocks. J. Cons. Int. Explor. Mer., 39: 175-192.
CrossRefDirect Link - Prager, M.H., E.D. Prince and D.W. Lee, 1995. Empirical length and weight conversion equations for blue marlin, white marlin and sailfish from the North Atlantic Ocean. Bull. Mar. Sci., 56: 201-210.
Direct Link - Richardson, D.R., R.K. Cowen, E.D. Prince and S. Sponaugle, 2009. Importance of the Straits of Florida spawning ground to Atlantic sailfish (Istiophorus platypterus) and blue marlin (Makaira nigricans). Fish. Oceanogr., 18: 402-418.
CrossRef - Richardson, D.E., J.K. Llopiz, K.D. Leaman, P.S. Vertes, F.E. Muller-Karger and R.K. Cowen, 2009. Sailfish (Istiophorus platypterus) spawning and larval environment in a Florida Current frontal eddy. Prog. Oceanogr., 82: 252-264.
CrossRef - Schaefer, K.M., 1998. Reproductive biology of yellowfin tuna (Thunnus albacares) in the eastern Pacific Ocean. Int. Am. Trop. Tuna Commun. Bull., 21: 201-272.
Direct Link - Shimose, T., K. Yokawa and H. Saito, 2010. Habitat and food partitioning of billfishes (Xiphioidei). J. Fish Biol., 76: 2418-2433.
CrossRef - Tamate, T. and K. Maekawa, 2000. Interpopulation variation in reproductive traits of female masu salmon, Oncorhynchus masou. Oikos, 90: 209-218.
CrossRef - Young, J.W., M.J. Lansdell, R.A. Campbell, S.P. Cooper, F. Juanes and M.A. Guest, 2010. Feeding ecology and niche segregation in oceanic top predators off eastern Australia. Mar. Biol., 157: 2347-2368.
CrossRef - Arocha, F. and L. Marcano, 2006. Life History Characteristics of Makaira nigricans, Tetrapturus albidus and Istiophorus albicans from the Eastern Caribbean Sea and Adjacent Waters. In: Reconciling Fisheries with Conservation: Proceedings of the Fourth World Fisheries Congress, Nielsen, J., J. Dodson, K. Friedland, T. Hamon, N. Hughes, J. Musick and E. Verspoor (Eds.). Symposium 49, American Fisheries Society, Bethesda, Maryland, pp: 587-597.
- Bumguardner, B.W. and J.D. Anderson, 2008. Age and growth, reproduccion and genetics of billfish in Gulf of Mexico waters off Texas. Proceedings from the Atlantic Billfish Research Program Simposium. Gulf States Marine Fisheries Commission Spring Meeting, March 11, 2008, Galveston, Texas, pp: 52-66.
- Hernandez-Herrera, A. and M. Ramirez-Rodriguez, 1998. Spawning seasonality and length at maturity of sailfish (Istiophorus platypterus) off the Pacific coast of Mexico. Bull. Mar. Sci., 63: 459-467.
Direct Link - Hoolihan, J.P., 2006. Age and growth of Indo-Pacific sailfish, Istiophorus platypterus, from the Arabian Gulf. Fish. Res., 78: 218-226.
CrossRef - Chiang, W.C., C.L. Sun, S.Z. Yeh, W.C. Su and D.C. Liu, 2006. Spawning frequency and batch fecundity of the sailfish (Istiophorus platypterus) (Istiophoridae) in waters off Eastern Taiwan. Zool. Stud., 45: 483-490.
Direct Link - Chiang, W.C., C.L. Sun, S.Z. Yeh, W.C. Su, D.C. Liu and W.Y. Chen, 2006. Sex ratios. Size at sexual maturity and spawning seasonality of sailfish Istiophorus platypterus from Esatern Taiwan. Bull. Mar. Sci., 79: 727-737.
Direct Link - Speare, P., 2003. Age and growth of black marlin Makaira indica, in the East coast Australian waters. Mar. Freshwater Res., 54: 307-314.
CrossRefDirect Link - Brown-Peterson, N.J., J.S. Franks, B.H. Comyns and J.R. McDowell, 2008. Reproductive biology, potential spawning and nursery areas and larval identification of blue marlin, Makaira nigricans, in the North central Gulf of Mexico. Proceedings from the Atlantic Billfish Research Program Symposium, (ABRPS'08), Gulf States Marine Fisheries Commission, pp: 92-133.
- Luckhurst, B.E., E.D. Prince, J.K. Llopiz, D. Snodgrass and E.B. Brhoters, 2006. Evidence of blue marln Makaira nigricans spawning in bermuda waters ans elevtes mercury levels in large specimens. Bull. Mar. Sci., 79: 691-704.
Direct Link - Molony, B., 2008. Fisheries biology and ecology of highly migratory species that commonly interact with industrialised longline and purse-seine fisheries in the western and central Pacific Ocean. WCPFC-SC4-2008/EB-IP-6, Department of Fisheries, Western Australia. http://www.wcpfc.int/doc/eb-ip-6/fisheries-biology-and-ecology-highly-migratory-species-commonly-interact-with-industrial.
- Shimose, T., M. Fujita, K. Yokawa, H. Saito and K. Tachihara, 2009. Reproductive biology of blue marlin Makaira nigricans around Yonaguni Island, Southwestern Japan. Fish. Sci., 75: 109-119.
CrossRef - Sun, CL., Y.J. Chang, C.C. Tszeng, S.Z. Yeh and N.Y. Su, 2009. Reproductive biology of blue marlin (Makaira nigricans) in the western Pacific Ocean. Fish. Bull., 107: 420-433.
Direct Link - Arocha, F. and A. Barrios, 2009. Sex ratios, spawning seasonality, sexual maturity and fecundity of white marlin (Tetrapturus albidus) from the Western Central Atlantic. Fish. Res., 95: 98-111.
CrossRefDirect Link - Oliveira, I.M., F.H.V. Hazin, P. Travassos, P.B. Pinheiro and H.G. Hazin, 2006. Preliminary results on the reproductive biology of the white marlin, Rapturus albidus Poey 1960, in the Western equatorial Atlantic Ocean. ICCAT Sci. Pap., 60: 1738-1745.
Direct Link - Arocha, F. and D.W. Lee, 1995. The spawning of swordfish from the Northwest Atlantic. Col. Vol. Sci. Pap. ICCAT, 44: 179-186.
Direct Link - De Martini, E., J.H. Uchiyama and H.A. Williams, 2000. Sexual maturity, sex ratio and size composition of swordfish, Xiphias gladius, caught by the Hawaii-based pelagic longline fishery. Fish. Bull., 98: 489-506.
Direct Link - Poisson, F. and C. Fauvel, 2009. Reproductive dynamics of swordfish (Xiphias gladius) in the Southwestern Indian Ocean (Reunion Island). Part 2: Fecundity and spawning pattern. Aquat. Living Resour., 22: 59-68.
CrossRef - Taylor, R.G. and M.D. Murphy, 1992. Reproductive biology of the swordfish Xiphias gladius in the Straits of Florida and adjacent waters. Fish. Bull., 90: 809-816.
Direct Link - Young, J., Q. Drake, M. Brickhill, J. Farley and T. Carter, 2003. Reproductive dynamics of broadbill swordfish Xiphias gladius, in the domestic longline fishery off Eastern Australia. Mar. Freshwater Res., 54: 315-332.
CrossRef