
N. Bouriga
Unite de Biologie marine. Faculte des Sciences de Tunis. 2092 Campus universitaire, Tunisie
M. Cherif
Institut National des Sciences et Technologies de la Mer (INSTM), Port de Peche 2060, La Goulette-Salammbo, Tunisie
G. Hajjej
Institut National des Sciences et Technologies de la Mer INSTM, 3018 Sfax, Tunisie
S. Selmi
Institut National des Sciences et Technologies de la Mer (INSTM), Port de Peche 2060, La Goulette-Salammbo, Tunisie
J-P. Quignard
Laboratoire d`Ichtyologie, Universite Montpellier II, Pl. E. Bataillon, case 102, 34095 Montpellier cedex, France
E. Faure
LATP, CNRS-UMR 6632, Evolution biologique et modelisation, case 5, Universite de Provence, Place Victor Hugo, 13331 Marseille cedex 3, France
M. Trabelsi
Unite de Biologie marine. Faculte des Sciences de Tunis. 2092 Campus universitaire, Tunisie
Journal of Fisheries and Aquatic Science
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 322-333







ABSTRACT
The growth, reproductive properties and seasonal variation in fatty acid profile of sand smelt Atherina sp. caught from Kerkennah islands in Tunisia were studied by sampling carried out between March 2008 and February 2009. A total of 793 specimens were examined. The sex of these species was determined macroscopically and the proportion of males (35.94%) was significantly lower than that of females (64.05%) with a ratio of 1:1.782 (males/females). The mean total length (TL) and weight (W) of the individuals ranged from 14-70 mm and 0.07-2.76 g, respectively. Regression coefficients (b) were estimated by using the logarithms of the total length and the total weight. Length-weight relationship was estimated as W = 0.0085TL2.9313 for females and W = 0.009TL2.8789 for males, being allometrically negative for both sexes. The reproductive season, evaluated from the gonado-somatic index (GSI), extended from March to June, with a peak in May. The levels of lipid extracted, and displayed pronounced seasonal fluctuations with the highest value in July (5.9 g/100 g fresh weight). Docosahexaenoic acid (DHA), linoleic acid and eicosapentaenoic acid (EPA) obtained by the method described by Metcalfe were the most abundant polyunsaturated fatty acids (PUFAs). The highest percentages for EPA and DHA were found in spring season and the highest amount were 6.48-9.32%, respectively for Sand Smelt in Kerkennah islands. The highest n-3/n-6 ratio was 1.65 in May and the lowest value was 0.69 in November.
PDF Abstract XML References Citation
Received: October 14, 2010;
Accepted: December 08, 2010;
Published: February 07, 2011
How to cite this article
N. Bouriga, M. Cherif, G. Hajjej, S. Selmi, J-P. Quignard, E. Faure and M. Trabelsi, 2011. Growth, Reproduction and Seasonal Variation in the Fatty Acid Composition of the Sand Smelt Atherina sp. from Kerkennah Islands, Tunisia. Journal of Fisheries and Aquatic Science, 6: 322-333.
DOI: 10.3923/jfas.2011.322.333
URL: https://scialert.net/abstract/?doi=jfas.2011.322.333
DOI: 10.3923/jfas.2011.322.333
URL: https://scialert.net/abstract/?doi=jfas.2011.322.333
INTRODUCTION
In Europe, the family Atherinidae is represented by two genera and six species, i.e., Atherinomorus lacunosus, a Lessepsian fish migrant, the sand smelts Atherina hepsetus, Atherina presbyter (Bucciarelli et al., 2002) and three possible species belonging to the Atherina boyeri complex: A. boyeri, A. punctata and A. lagunae as respectively non-punctuated marine, punctuated marine and lagoon atherines (Trabelsi et al., 2002a, b). Recently, Trabelsi et al. (2009) reported that Atherina populations which inhabit marine shallow waters of the coasts of the Kerkennah Islands in Tunisia are A. lagunae including firstly sympatric speciations followed by a post-Pleistocene colonisation of the lagoons.
The sand smelt, Atherina boyeri is common in the Mediterranean and adjacent seas and in the northeast Atlantic from the Azores to the northwestern coast of Scotland (Quignard and Pras, 1986). It is a small, short-lived, euryhaline atherinid fish that mainly inhabits coastal and estuarine waters including coastal lagoons, salt marshes and more rarely, inland waters, over a wide range of salinities from freshwater to hypersaline conditions (Henderson and Bamber, 1987).
The age, growth, mortality, reproductive and feeding biology of this species has been investigated by several authors (Leonardos and Sinis, 2000; Pallaoro et al., 2002; Patimar et al., 2009). Marine food and especially marine fish are an important part of the Mediterranean diet. The beneficial effect of fish consumption on human health has been related, among other factors to the high content of n-3 or ω 3 fatty acids, especially eicosapentaenoic acid (C20:5n-3) and docosahexaenoic acid (C22:6n-3). Today it is known that is diet rich in n-3 fatty acids is essential for normal growth and development and may play an important role in the prevention and treatment of coronary artery disease, diabetes, hypertension and cancer. They also affect neurodevelopment in infants, fat glycemic control, learning ability and visual function (Goodstine et al., 2003). It is thus important, for human health to increase the consumption of fish or fish products, which are rich in polyunsaturated fatty acids of the n-3 family and poor in polyunsaturated fatty acids of the n-6 family (Sidhu, 2003).
In Tunisia, the sand smelt namely Scharkaw, is the most commercialized fresh and dried species especially in the centre and south coast of Tunisia. The aim of this study was to describe the growth, reproduction and the fatty acid profile of the sand smelt population in Kerkennah islands, which has not been studied in the area and which provides an insight into the life history of this species and to test whether this region has developed locally different life-history strategies of this species.
MATERIALS AND METHODS
The present study was carried out in Kerkennah islands in Tunisia. A total 793 individuals of Atherina sp. were collected monthly from March 2008 to February 2009 at three sites of the Kerkennah Islands coasts: El Attaya (34°44’N, 11°18’E), Sidi Fraj (34°41’N, 11°07’E) and Sidi Youssef (34°38’N, 10°58’E) (Fig. 1). The fish were collected with a small net (5 mm mesh) used locally for the sand smelt fishery. It is one square meter of net on a metal frame connected by ropes to a main rope; when the fish appear above the net it is lifted out of the water.
Fresh specimens were transported to the laboratory where total (TL) and Standard Length (SL) were measured to the nearest 0.1 mm. Total Weight (Wt), and Weight of Gonads (Wg) recorded with an electronic analytical balance to the nearest 0.01 g. Gonads were examined macroscopically to determine sex (257 males and 458 females). Samples for biochemical analysis were kept in ice, transported immediately to the laboratory where they were rapidly weighted, measured frozen in liquid nitrogen and stored at -40°C until analysis.
Sex ratio (SR): The sex-ratio was estimated monthly during the study period, being expressed as the percentage of females in the various size classes (Kartas and Quignard, 1984).
 | |
| Fig. 1: | Sampling area |
The overall sex ratio was assessed using Chi square test (χ2) (Sokal and Rohlf, 1987);
Where:
| fiobs | = | Observed proportion. |
| fth | = | Theoretical proportion. |
Statistical analyses were performed with SPSS 11.5 software package and a significant level of 0.05 was accepted.
Sexual cycle and period of spawning: The spawning period of sand smelt was determined by analyzing the evolution of the gonado-somatic index (GSI). The GSI was calculated as: GSI = Gonads Weight (Wg).100/Total Weight (Wt) (Wootton, 1990). The GSI was computed for each month and in total for females males and both sexes together. Comparison of GSI between months was carried out by analysis of variance (ANOVA).
Length-weight relationship: The relationship between the total length and total weight were determined by fitting the data to a potential relationship in the form of: W = aLb (Ricker, 1973) where W is the weight in grams, L the total length in centimeters, a and b are the parameters to be estimated, with b being the coefficient of allometry based on the test given by Schneider et al. (2000). The degree of association between the variables was computed by the determination coefficient, R2. Student’s t-test was used to determine if the coefficient b was significantly different from 3 (Dagnelie, 2007).
Fatty acid profile: For this analysis samples were collected at bimonthly intervals for a one-year period from the local fishers in Kerkennah islands.
Total lipids were extracted according to the method of Folch et al. (1957) using chloroform/methanol (2/1). Aliquots of the chloroform layer extracts were evaporated to dryness under nitrogen and the lipids were quantified gravimetrically.
Fatty acids methyl esters (FAMEs) were obtained by the method described by Metcalfe et al. (1966). A fraction of the lipid extract was saponified with 0.5 N NaOH in methanol followed by methylation in 14% boron trifluoride in methanol (BF3/MeOH). The methylated sample was then extracted with 8 mL n-hexane. All of these reactions were performed in quadruplet for each sample. The resulting methyl esters were analysed using an Agilent Gaz chromatograph system 6890N equipped with a Flame Ionization Detector (FID), a splitless injector and a polar INNOWAX fused silica capillary column (30 m *0.25 mm i.d. *0.25 μm film thickness). The temperature of the injector and the detector were 250 and 275°C respectively. Helium was used as a carrier gas with a flow rate of 1.5 mL min-1. Peaks were identified by comparison of their retention times with PUFA 3 FAMEs standards (SUPELCO).
Statistical analysis: Statistical analysis was performed using SPSS software, version 10.0. The comparison of different biochemical parameters were tested using Duncan’s test (95% confidence interval) with one-way ANOVA.
RESULTS AND DISCUSSION
This study has established key population parameters of Atherina sp. in Kerkennah islands. Variation in life-history parameters of species could be explained on the basis of the different exploitation patterns and/or ecological conditions.
Sexual cycle and spawning period: The Gonado-Somatic Index (GSI) was used to determine the reproductive period, which was calculated from samples taken monthly from males and females. The average GSI during the 12-month period was 1.87% for females and 1.82% for males. The GSI began to rise in March (5.22% for females and 4.02% for males, peaked in May (8.72% for females and 7.37% for males) and dropped in June (females 4.27% and males 3.71%) and July (females 3.98% and males 3.22%) (Fig. 2). In July, the GSI value decreased because all samples had presumably dispensed of their eggs. No specimens of either sex with an above average GSI were found in July-January.
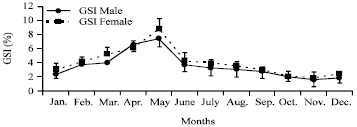 | |
| Fig. 2: | Monthly changes in the gonadosomatic index (GSI) of Atherina sp. (female and male) |
After July the gonads began to develop and the values of GSI again started to gradually increase until January.
This study showed that the highest average values of GSI were observed in March for males and females respectively. The GSI of both sexes followed almost the same pattern. The monthly evolution of Gonado-Somatic Index (GSI) of the sand smelt in Kerkennah islands indicated an extended spawning period, from March to June, with a peak in May when GSI is considerably higher. Then, it decreases in July showing the start of the resting period. In Iznik lake, the maximum GSI was reached in May for boy sexes, whilst it was in April from brackish lagoons of Southern France (Bartulovic et al., 2006). Leonardos and Sinis (2000), and Patimar et al. (2009) observed a maximum spawning from March to the end of July.
Fernandez-Delgado et al. (1988) compared the spawning season of Atherina boyeri in the outlet of the Guadalquivir river (south of Spain) to that reported by Palmer and Culley (1983) for the British waters and found that, in southern latitudes the spawning period was earlier in January. The spawning period can be changed because of the latitude and climatic conditions. In general, inshore species in lower latitudes exhibit a spawning period longer (Longhurst and Pauly, 1987). According to Nikolsky (1980), the spawning properties of a fish alter in relation to the species and ecological characteristics of water system which they inhabit.
Sex-ratio: All individuals were sexed, the proportion of males was significantly lower than that of females, with reference to the distribution of males and females in the Atherina sp. samples, the females predominate in all months (Fig. 3).
The overall sex-ratio value estimated as 64 % in favour to females. Chi-square test showed a significant difference (χ2 obs = 32.11>>χ2 th = 3.84; p<0.05). Further, an unequal sex ratio was observed, females were dominant in all size classes (Fig. 4). The sex ratio for the whole sample was 1:1.782 (males/females). The overall sex ratio in Atherina sp. was significantly different from the ratio 1:1 (p<0.05) indicated a dominance of females in Kerkennah islands, as was observed for Atherina boyeri by Gon and Ben-Tuvia (1983). However, Turnpenny et al. (1981) and Creech (1992) did not suggest significant sex ratio differences in samples of Atherina presbyter. Fernandez-Delgado et al. (1988) pointed out Atherina boyeri was composed of shoals with a non-balanced sex ratio in the pre-spawning and spawning periods.
Recent study showed that the overall sex ratio for Atherina boyeri was 1:1.24 in the Mesolongi and Etolikon lagoons (Leonardos and Sinis, 2000), 1:1.30 in the Malta Neretva River system (Bartulovic et al., 2004a), 1:1.25 in an estuarine system of Northen Greece (Koutrakis et al., 2004), 1:1.30 in the Gomishan Wetland (Patimar et al., 2009), 1:1.7 in Lake Iznik in favor of females.
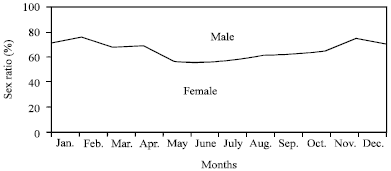 | |
| Fig. 3: | Sex ratio monthly evolution for Atherina sp. (female and male) |
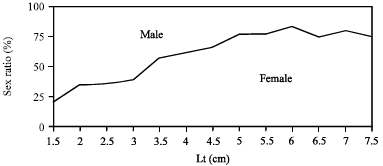 | |
| Fig. 4: | Sex ratio evolution by size class in Atherina sp. (female and male) |
Length-weight relationship: Of the 793 specimens ranged in size and weight from 14-70 mm TL and 0.07-2.76 g WT, respectively, 458 were females and 257 were males. Females measured 2.1 to 7.0 cm TL (x = 44.01 mm TL), males 1.4 to 6.7 cm TL (x =45.87 mm TL). Moreover, weight distribution was found to range from 0.07-2.64 g for females and from 0.09-2.76 g for males.
Leonardos and Sinis (2000) and Bartulovic et al. (2004a) reported that the maximum total length and weight of Atherina boyeri were 110 mm and 9.0 g for males and 115.0 mm and 9.0 g for females respectively. Henderson and Bamber (1987) indicated that sand smelt Atherina boyeri population can adapt its morphology to environmental condition and growth and maximum size can reduce along an oceanic-coastal-estuarine-freshwater habit range.
The length-weight relationship of Atherina sp. in Kerkennah islands indicated a negative allometry (Fig. 5 and 6) and was described by the following equation:
WT = 0.0084TL2.9290. The analysis by sex (males: WT = 0.009TL2.8789; females: WT =0.0085TL2.9313) showed a significant difference in the b coefficient (Table 1). The results of statistical analyses showed significant differences between males and females. The value b for males (2.8789), for females (2.9313) and both sexes (2.9290) displayed a negative allometric growth.
The experimental relationship between total length and weight indicated a negative allometric growth (b<3) Fig. 6 . The b value estimated for the sand smelt population from the study of Gon and Ben-Tuvia (1983) showed a negative allometric growth from the Bardawil lagoon, Sinai population where the salinity of water is 40-110% (hyperhaline) as 2.93, while Andreu-Soler et al. (2003), Bartulovic et al. (2004a), Koutrakis et al. (2004) and Patimar et al. (2009) reported a positive allometric growth. Patimar et al. (2009) mentioned that the variation “b” values between populations can be affected differently, because of the geographic location and environmental conditions.
Total lipids: The fat content of the analysed samples shows an important dependency on season (Table 2). Substantial differences between the seasons were observed in the lipid levels. The lipid levels were between 2.2 and 5.9 g/100 g fresh weight. The highest level of lipid was found in July while the lowest was in November.
A high fat content in the summer and a low fat content in the winter has also been reported for the Japanese sardine Sardinops melanostictus (Shirai et al., 2002). Researching the effect of climate on lipid content variation, Krzynowek (1985) reported that the fat content of some fish species might vary by approximately 10% according to the season. The percentage of body fat is known to depend on the life cycle stage and energy intake of the animal (Jobling, 1994) and higher temperature periods are characterised by faster growth rates and large intakes (Garcia et al., 2001). Our studies about sexual cycle and the spawning period indicated that the reproductive season of the specie is between March and June.
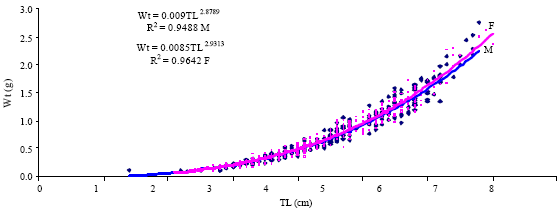 | |
| Fig. 5: | Relationship between Total length (TL) and Total Weight (W) in Atherina sp. (Sexes separated) |
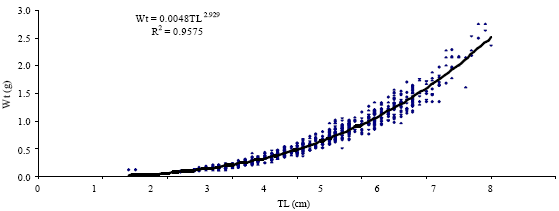 | |
| Fig. 6: | Relationship between Total Length (TL) and Total Weight (WT) in Atherina sp. (sexes combined) |
| Table 1: | Values of the regression parameters (a, b, R2) of the length-weight relationship |
 | |
| Table 2: | Lipids composition of Atherina sp. in different seasons (g/100 g fresh weight) |
| Av: Analysis of variance. ***Significant at 0.001 | |
In this period, sexual maturation has been found to reduce lipid body stores in sand smelt because lipid stores are directed to gonad lipids or used for energy. As many fish species tend to reduce their food intake during sexual maturation, essential fatty acids and other nutrients needed for ovarian growth are taken from the reserves in their bodies. Low levels of lipid were observed in the reproductive season of the species and at the end of autumn.
For the sand smelts from the Kerkennah islands, the maximum level of lipid determined in July may indicate increasing feeding and growth rate of fish. The increase of lipid level in summer may also be explained by an enhanced feeding activity due to higher sea water temperature and increased day length which have a positive influence on appetite (Olsson et al., 2002). Means (n = 6) with the same letter within rows are not significantly different (p>0.05).
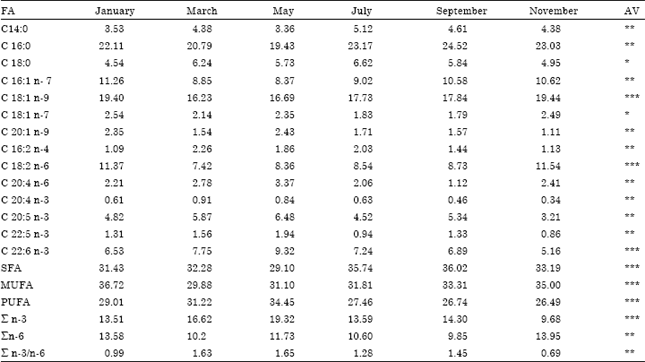
Fatty acid profile: The fatty acid compositions of the Atherina sp. are presented in Table 3. The results show remarkable changes in the individual fatty acids during the one-year period. The fatty acid profile in silverside was typical of marine animals with the dominance of the palmitic acid (C16:0) for Saturated Fatty Acids (SFA), contributing approximately 19.43-24.52% to the total SFA content of the lipids for Atherina sp. (Passi et al., 2002) pointed out that palmitic acid was a key metabolite in fish and its level was not influenced by diet. Also, the Oleic acid (C18:1) was identified as the major monounsaturated fatty acids MUFA in the fish (16.23-19.44% of total MUFAs). According to the season, DHA (C22:6 n-3), LA (C18:2 n-6) and EPA (C20:5 n-3) were the predominant PUFAs. The same data were found by Pirini et al. (2002) and Surh et al. (2003). An increase in the lipid level resulted in an increase also in the fatty acid levels, except for n-3. However, most of the fatty acids determined in July were saturated fatty acids, followed by mono unsaturated fatty acids. The maximum level of n-3 was found in May. From these results, it could be speculated that Sand Smelt in Kerkennah islands uses the n-3 series as an energy source.
In the present study, the percentages (in total lipid) of EPA and DHA which have a vital role in human nutrition were between 3.21-6.48 and 5.16-9.32%, respectively. The highest percentages for EPA and DHA were found in spring season and the highest amounts were 6.48-9.32%, respectively for the Sand Smelts from Kerkennah islands. Thus, among the n-3 series, the Sand smelt are good sources of EPA and DHA. Sargent (1996) reported that n-3 PUFA, principally DHA, has a role in maintaining the structure and functional integrity of fish cells. In addition, DHA has a specific and important role in neural cell membranes, the brain and eyes.
| Table 3: | Seasonal fatty acid composition of Atherina sp. (% of total lipid) |
 | |
| * Significance at 0.05; ** Significance at 0.01; *** Significance at 0.001. Av: Analysis of variance | |
Moreover, it is considered a desirable property in fish for human nutrition and health.
The results obtained in this study show shorter (C: 18) chain n-3 acids in the food to be elongated and desaturated in the sand smelt body, where longer-chain PUFAs, mainly DHA, are formed. The results demonstrate that Atherina sp. is highly capable of transforming native forms of C18n-3 into long-chain acids, which leads to a high DHA content. Similar results were obtained by Xu et al. (2001) who analysed dietary effects on fatty acid composition in muscles of Perca fluviatilis; they found high DHA contents. There are close relationships between the fish lipid composition and the diets of fish (McKenzie et al., 2000). The most common prey of Sand Smelt were Crustaceans, copepods, gammarid amphipods, cladocerans, decapod larvae, mysids and insects. Due to their consumption of preys in which chain elongation and desaturationis completed, the fish were rich in long chain n-3 PUFAs (Bartulovic et al., 2004b).
The n-3/n-6 ratio is a good index for comparing relative nutritional value of fish oils (Pigott and Tucker, 1990). It is important for human health to increase the consumption of fish or fish products, which are rich in PUFAs of the n-3 family (Marchioli et al., 2002). However, there is no recommended intake in terms of n-3/n-6 ratios but evidence in wild animals and estimated nutrient intake during human evolution suggest a diet ratio of 1:1 (Simopoulos, 1989). The present data show that the n-3/n-6 ratio was 1.65 in May, 1.63 in March, 1.45 in September, 0.99 in January and that the lowest value 0.69 was in November. A high level of n-6 lowered the n-3/n-6 ratio in November.
In the human dietary, an increase of the n-3/n-6 fatty acid ratio is essential to help reducing coronary heart disease by reducing plasma lipids and to reduce cancer risk (Kinsella et al., 1990). It is thus important for human health to increase the consumption of n-3 fatty acids (Coetzee and Hoffman, 2002).
This study has shown that the Sand Smelt Atherina sp. in Kerkennah islands is a suitable item in the human diet during the spring season when the levels of EPA, DHA and n-3/n-6 ratio are considered. This condition can be regarded as an explanation for the fact that the Sand Smelt in Kerkennah islands are richer in n-3 fatty acids, taking into consideration with the fatty acid profile of the fish. As a consequence, when human health is taken into account, the Sand Smelt appears to be quite nutritious in terms of fatty acid composition and ratio.
CONCLUSION
The Sand Smelt Atherina sp. caught from Kerkennah islands shows a great variation in growth and reproduction proprieties from many other populations in its distribution range. This study has shown that the Sand Smelt is a desirable item in the human diet when the levels of EPA and DHA are considered. This condition can be regarded as an explanation for the fact that the fish in Kerkennah islands are richer in n-3 fatty acids in spring and summer, taking into consideration with the fatty acid profile of the fish. As a consequence, when human health is taken into account, the Sand Smelt from Kerkennah islands appears to be quite nutritious in terms of fatty acid composition and ratio. Revealed the biology of Sand Smelt population in Kerkennah islands in Tunisia is very important to plan the fishery strategy in relation to its commercial value.
ACKNOWLEDGMENT
I would like to thank Dr. Anissa Chouikha for her great helpful comments on the manuscript. I also thank Faiza Trabelsi for her financial support of this research.
REFERENCES
- Andreu-Soler, A., F.J. Olivia-Paterna, C. Fernandez-Delgado and M. Torralva, 2003. Age and growth of the sans smelt, Atherina boyeri (Risso, 1810) in the mar menor coastal lagoon (SE Iberian Peninsula). J. Applied Ichthyol., 19: 202-208.
CrossRefDirect Link - Bartulovic, V., B. Glamuzina, A. Conides, A. Gavilovie and J. Duleie, 2006. Maturation, reproduction and recruitement of the sand smelt, Atherina boyeri Risso,1810 (Piscies, Atherinidae) in the estuary of mala neretva river (South Eastern Adriatic, Croatia). Acta Adriat., 47: 5-11.
Direct Link - Bucciarelli, G., D. Golani, and G. Bernardi, 2002. Genetic cryptic species as biological invaders: The case of a lessepsian fish migrant, the hardyhead silverside Atherinomorus lacunosus. J. Exp. Mar. Biol. Ecol., 273: 143-149.
Direct Link - Coetzee, G.J.M. and L.C. Hoffman, 2002. Effects of various dietary n-3/n-6 fatty acid ratios on the performance and body composition of broilers. S. Afr. J. Anim. Sci., 32: 175-184.
Direct Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Garcia, B., F.M. Rueda, M.D. Hernandez, F. Aguado, M.A. Egea and F. Faraco, 2001. Growth and Conversion Index of Sharpsnout seabream (Diplodus puntazzo, 1789). In: Feedlot Tanks, Fernandez-Palacios, H. and M. Zquiredo (Eds.). Monographs of the Canarian Institute of Marine Science, Las Palmas de Gran Canaria, Spain.
- Goodstine, S.L., T. Zheng, T.R. Holford, B.A. Ward, D. Carter, P.H. Owens and S.T. Mayne, 2003. Dietary (n-3)/(n-6) fatty acid ratio: Possible relationship to premenopausal but not postmenopausal breast cancer risk in US women. J. Nutr., 133: 1409-1414.
Direct Link - Kinsella, J.E., B. Lokesh and R.A. Stone, 1990. Dietary n-3 polyunsaturated fatty acid and amelioration of cardiovascular disease: Possible mechanisms. Am. J. Clin. Nutr., 52: 1-28.
PubMed - Koutrakis, E.T., N.I. Kamidis and I.D. Leonardos, 2004. Age, growth and mortality of a semi-isolated lagoon population of sand smelt, Atherina boyeri (Risso, 1810) (Pisces: Atherinidae) in an estuarine system of Northern Greece. J. Applied Ichthyol., 20: 382-388.
CrossRefDirect Link - Marchioli, R., F. Barzi, E. Bomba, C. Chieffo and D. Di Gregorio et al., 2002. Early protection against sudden death by n-3 polyunsaturated fatty acids after myocardial infarction: Timecourse analysis of the results of GISSI-prevenzione. Circulation, 105: 1897-1903.
PubMed - McKenzie, D.J., G. Piraccini, M. Piccolella, J.F. Steffensen, C.L. Bolis and E.W. Taylor, 2000. Effects of dietary fatty acid composition on metabolic rate and responses to hypoxia in the European eel (Anguilla anguilla). Fish Physiol. Biochem., 22: 281-296.
CrossRefDirect Link - Metcalfe, L.D., A.A. Schmitz and J.R. Pelka, 1966. Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem., 38: 514-515.
CrossRefDirect Link - Olsson, G.B., R.L. Olsen, M. Carlehog and R. Ofstad, 2003. Seasonal variation in chemical and sensory characteristics of farmed and wild Atlantic halibut (Hippoglossus hippoglossus). Aquaculture, 217: 191-205.
Direct Link - Pallaoro, A., M. Franieevie and S. Matie, 2002. Age, growth and mortality of big scale sand smelt, Atherina (Hepstia) boyeri Risso, 1810 in the pantana lagoon, Croatia. Periodicum Biol., 104: 175-183.
Direct Link - Passi, S., S. Cataudella, P. Di Marco, F. De Simone and L. Rastrelli, 2002. Fatty acid composition and antioxidant levels in muscle tissue of different mediterranean marine species of fish and shellfish. J. Agric. Food. Chem., 50: 7314-7322.
Direct Link - Patimar, R., M.M. Yousefi and S.M. Hosieni, 2009. Age growth and reproduction of the sand smelt Atherina boyeri Risso, 1810 in the Gomishan weltland-southeast Caspian Sea. Estuar. Coast Shelf Sci., 81: 457-462.
Direct Link - Pigott, G.M. and B.W. Tucker, 1990. Seafood: Effects of Technology on Nutrition. Marcel Dekker, Inc., New York, USA., ISBN-13: 9780824779221, Pages: 384.
Direct Link - Pirini, M., P.P. Gatta, S. Testi, G. Trigari and P.G. Monetti, 2002. Effect of refrigerated storage on muscle lipid quality of sea bass (Dicentrarchus labrax) fed on diets containing different levels of vitamin E. Food Chem., 68: 289-293.
CrossRef - Ricker, W.E., 1973. Linear regression in fisheries research. J. Fish. Res. Board Can., 30: 409-434.
Direct Link - Shirai, N., H. Suzuki, S. Tokairin, H. Ehara and S. Wada, 2002. Dietary and seasonal effects on the dorsal meat lipid composition of Japanese (Silurus asotus) and Thai catfish (Clarias macrocephalus and hybrid Clarias macrocephalus and Clarias galipinus). Comp. Biochem. Phys., 132: 609-619.
PubMedDirect Link - Sidhu, K.S., 2003. Health benefits and potential risks related to consumption of fish or fish oil. Reg. Toxicol. Pharm., 38: 336-344.
CrossRefPubMedDirect Link - Simopoulos, A.P., 1989. Summary of NATO advanced research workshop on dietery omega-3 and omega-6 fatty acids: Biological effects nutritional essentiality. J. Nutr., 199: 512-528.
Direct Link - Surh, J., J.S. Ryu and H. Kwon, 2003. Seasonal Variations of Fatty acid compositions in various Korean shellfish. J. Agric. Food Chem., 51: 1617-1622.
CrossRefDirect Link - Trabelsi, M., E. Faure, J.P. Quignard, M. Boussaid, B. Focant and F. Maamouri, 2002. Atherina punctata and Atherina laguna (Pisces, Atherinidae) new species found in the Mediterranean sea. 1. Morphological investigations of three Atherinid species. C. R. Biol., 325: 967-975.
CrossRef - Trabelsi, M., A. Gilles, C. Fleury, F. Maamouri, J.P. Quignard and E. Faure, 2002. Atherina punctata and Atherina lagunae (Pisces, Atherinidae), new species found in the Mediterranean Sea. 2. Molecular investigations of three Atherinid species. C.R. Biol., 325: 1119-1128.
Direct Link - Trabelsi, M., N. Bourigat, D. Aurelle, J.P. Quignard, R. Barthelemy and E. Faure, 2009. Some marine tunisian Atherina boyeri populations (Teleostei) have morphological and molecular characteristics of lagoon fishes. Open Mar. Biol. J., 3: 59-69.
Direct Link - Turnpenny, A.W.H., R.N. Bamber and P.A. Henderson, 1981. Biology of the sand-smelt (Atherina presbyter Valenciennes) around Fawley power station. J. Fish. Biol., 18: 417-427.
CrossRefDirect Link - Xu, X.L., P. Fontaine, C. Melard and P. Kestemont, 2001. Effects of dietary fat levels on growth, feed efficiency and biochemical compositions of Eurasian perch Perca fluviatilis. Aquacult. Int., 9: 437-449.
CrossRefDirect Link - Bartulovic, V., B. Glamuzina, A. Conides, J. Dulcic, D. Lucic, J. Njire and V. Kozul, 2004. Age, growth, mortality and sex ratio of sand smelt, Atherina boyeri Risso, 1810 (Pisces: Atherinidae) in the estuary of the Mala Neretva River (Middle-Eastern Adriatic, Croatia). J. Applied Ichthyol., 20: 427-430.
CrossRefDirect Link