
L.S. Kavan
Science and Research Branch, Islamic Azad University, Iran
S.R. Gilkolaei
Iranian Fisheries Research Organization, Iran
G. Vossoughi
Science and Research Branch, Islamic Azad University, Iran
S.M.R. Fatemi
Department of Marine Biology,Agricultural Science and Natural Resources, Iran
R. Safari
Department of Fisheries, Agricultural Science and Natural Resources, Iran
S. Jamili
Iranian Fisheries Research Organization, Iran
Journal of Fisheries and Aquatic Science
Year: 2009 | Volume: 4 | Issue: 6 | Page No.: 316-322







ABSTRACT
The genetic diversity and population structure of Rutilus frisii kutum from three regions in the Iranian coastline and one region from Azerbaijan were investigated using microsatellite DNA markers. Genomic DNA from 140 specimens was extracted and using PCR approach nine loci with reasonable polymorphism were amplified. The results showed that the lowest mean number of alleles per locus (5.22) was observed in Tonekabon River and the highest (5.77) in Azerbaijan population. The observed heterozygosity in the Tonekabon River (0.625) population was higher than those of the other two populations in Iran and Azerbaijan population (0.473). Significant deviations from HWE were found at more loci in the Iranian populations than Azeri population. In spite of geographical distance, both the highest and lowest population differentiation (Fst) value was between Iranian populations not among the Iranian and Azerbaijan populations. The highest and significant was between Khoshkrud and Tonekabon (0.098) and the lowest and significant was between Khoshkrud and Gorganrud (0.062). The genetic distance was the lowest (0.27) between the Khoshkrud and Gorganrud populations, whereas the highest distance (0.493) was between Khoshkrud and Tonekabon River.The AMOVA analysis with consideration of 2 sampling regions (Iran and Azerbaijan) and 4 sampling locations (Iran: Khoshkrud, Tonekabon, Gorganrud and Azerbaijan: the Kura mouth) revealed that almost all of the variance in data namely 86% (p = 0.01) was within locations, genetic variances among locations was 12% (p = 0.01) and among regions was 2% (p = 0.01). The reported results could be of interest for management and conservation programmes of this species in the Caspian Sea.
PDF Abstract XML References Citation
How to cite this article
L.S. Kavan, S.R. Gilkolaei, G. Vossoughi, S.M.R. Fatemi, R. Safari and S. Jamili, 2009. Population Genetic Study of Rutilus frisii kutum (Kamansky 1901) from the Caspian Sea; Iran and Azerbaijan Regions, using Microsatellite Markers. Journal of Fisheries and Aquatic Science, 4: 316-322.
DOI: 10.3923/jfas.2009.316.322
URL: https://scialert.net/abstract/?doi=jfas.2009.316.322
DOI: 10.3923/jfas.2009.316.322
URL: https://scialert.net/abstract/?doi=jfas.2009.316.322
INTRODUCTION
Kutum, Rutilus frisii kutum (Kamenskii, 1901), live in the Caspian Sea near the coast, including Azeri, Russian, Turkmen and Iranian waters. This species is known as a migratory anadromous fish spawning in rivers (Iranian Coastline: Haviq, Lemir, Khoshkrud, Tajan, Shafarud, Gorganrud; Azerbaijan: Kura) in March-April. This is a very valuable commercial fish in the southern part of the Caspian Sea and has a great demand, due to its good taste and culinary customs of the local people. It’s stocks declined dramatically mainly due to over fishing, illegal catch, pollution and deterioration of habitats and natural spawning grounds in most of its spawning. In the 2000 IUCN Red List this species has been listed as Data Deficient (Taylor and Mittermeier, 2000). Coad (2000), using 18 criteria, found this species to be one of the top 4 threatened species of freshwater fishes in Iran. Therefore, artificial propagation became the chief way for the maintenance and increase of Kutum abundance. About 140 million specimens of kutum fingerlings (1-2 g weight) are released into the sea annually from two Iranian hatcheries; this resulted in significant increase of catches of Iran in recent years. Scince well established that genetic variation promotes adaptation to changing environmental conditions and heterozygous individuals are usually superior to less heterozygous individuals in many economically important characteristics like growth, fertility and disease resistance (Beardmore et al., 1997), conservation of resources is an essential component of species management programmes (Connell and Wright, 1997; Gilkolaei, 1997). An effective strategy for the conservation of a particular species should, part in, be determined by information on its genetic structure.
Microsatellites represent co-dominant molecular genetic markers, which are ubiquitously distributed within genomes. Due to their high level of polymorphism, relatively small size and rapid detection protocols, these markers are widely used in a variety of fundamental and applied fields of life and medical sciences. In the field of aquaculture, microsatellites represent workhorse markers, which are useful for the characterization of genetic stocks, broodstock selection, constructing dense linkage maps, mapping economically important quantitative traits, identifying genes responsible for these traits and application to marker assisted breeding programmes (Chistiakov et al., 2006). The objective of this study was to compare the levels of genetic variation of Kutum samples between Iranian and Azeri populations based on microsatellites.
MATERIALS AND METHODS
Fish Samples and DNA Extraction
The fish were caught from 3 different regions of the Iranian coastline (Khoshkrud, Tonekabon, Gorganrud) and 1 region in Azerbaijan (Waters of the Caspian Sea close to Kura River mouth) (Fig. 1). Musscles tissue samples of 35 fish from each location were stored in 1.5 mL Eppendorf tubes with 96% ethanol for subsequent DNA extraction and amplification.
 | |
| Fig. 1: | Map showing sampling locations of four populations of Rutilus frisii kutum |
Genomic DNA was extracted from a 1 cm2 (50-60 mg) piece of muscle using the phenol-chloroform procedure described by Hillis and Moritz (1990). The quality and concentration of DNA from samples were assessed by 1% agarose gel electro-phoresis and then the samples were stored at -20°C for further studies.
PCR Amplification and Electrophoresis
Nine microsatellite loci were analysed: Ca1, Ca2, Ca3, Ca4 (Dimsoski et al., 2000), Lco1, Lco2, Lco3, Lco 4 (Turner et al., 2004) and MFW2 (Croojimans et al., 1997). GenBank Accessionnumbers are AF277573, AF277574, AF277575, AF277576, AY318777, AY318778, AY318779, AY318780 and EF144125, respectively. The Polymerase Chain Reaction (PCR) conditions, especially the annealing temperatures, were optimized for the 9 microsatellite loci as necessary to produce scorablestorable amplification products. Annealing temperatures were 55°C for Ca1, 58°C for Ca2 and Ca3 and 61°C for Ca4, 60°C for Lco1, 62°C for Lco2, 53°C for Lco3, 57°C for Lco4 and 66°C for MFW2. Amplification was performed in PCR system (Gradient Eppendorf) using a 25 μL reaction mixture. Each PCR reaction (final volume 25 μL) was composed of 5 μL of 10X reaction buffer, dNTPs 10 mM, MgCl2 50 mM, primer 20 pmol, genomic DNA 100 ng and 1.5-2 unit of Taq polymerase. The temperature profile consisted of a 3-mininitial denaturation at 94°C, followed by 30 cycles of: 30 sec at 94°C, 30 sec at the respective annealing temperature and 1 min at 72°C, ending with 5 min at 72°C. PCR products were separated on 8% polyacrylamide gels stained with silver nitrate. The recorded microsatellite genotypes were used as input data for the GENALEX software version 6 package (Peakall and Smouse, 2006) in order to calculate allele and genotype frequencies, observed (Ho) and (He) expected heterozygosities and to test for deviations from Hardy-Weinberg equilibrium. Genetic distance between two populations was estimated from Nei standard genetic distance and genetic similarity index (Nei, 1972). Genetic differentiation between populations was also evaluated by the calculation of pairwise estimates of Fst values. All calculations were conducted using the GENALEX version 6.
RESULTS
Within populations, the lowest mean number of alleles per locus (5.22) was observed in Tonekabon River and the highest (5.77) in Azerbaijan population. The observed heterozygosity in the Tonekabon river (0.625) population was higher than those of the other two populations in Iran and Azerbaijan population (0.473) (Table 1). Significant to highly significant deviations from Hardy-Weinberg expectations were observed in 30 out of 36 (nine locixfour populations) cases (Table 1), showing deficit of heterozygote. Population differentiation was modest among all populations. Both the highest and lowest population differentiation (Fst) value was between Iranian populations not among the Iranian and Azerbaijan populations. The highest and significant (p = 0.01) was between Khoshkrud and Tonekabon (0.098) and the lowest and significant (p = 0.01) was between Khoshkrud and Gorganrud (0.062) (Table 2). The estimated gene flow (Nm) value between the Gorganrud and Khoshkrud populations across all the studied loci was the highest, while the Nm value between the Khoshkrud and Tonekabon populations was the lowest (Table 2). Genetic distance (D) and genetic similarity index (I) between any four populations were shown in Table 3. The genetic distance was the smallest (0.27) between the Khoshkrud and Gorganrud populations, whereas the largest distance (0.493) was between Khoshkrud and Tonekabon River.The AMOVA analysis with consideration of 2 sampling regions (Iran and Azerbaijan) and 4 sampling locations (Iran: Khoshkrud, Tonekabon, Gorganrud and Azerbaijan: the Kura mouth) revealed that almost all of the variance in data namely 86% (p = 0.01) was within locations, Genetic variances among locations was 12% (p = 0.01) and among regions was 2% (p = 0.01) (Table 4).
| Table 1: | Variability of nine microsatellite loci in four Rutilus frisii kutum populations from Iran and Azerbaijan |
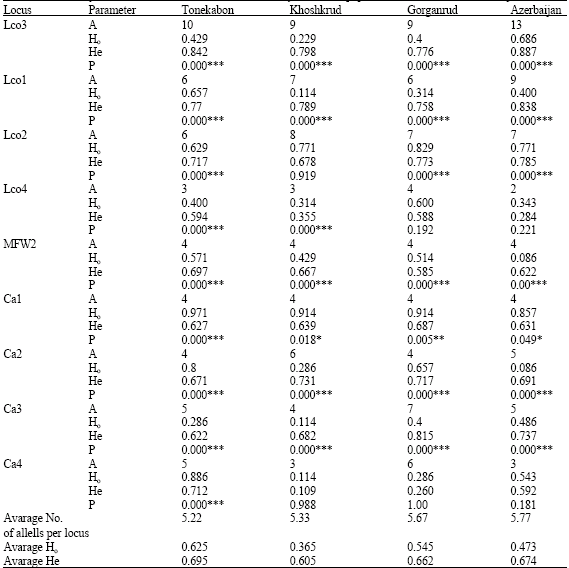 | |
| A: No. of alleles; Ho: Observed heterozygosity; He: Expected heterozygosity; P: p-values of χ2tests for Hardy-Weinberg equilibrium are significant | |
| Table 2: | Multi locus Nm(above diagonal) and Fst values (below diagonal) between pairs of Rutilus frisii Kutum population across all loci |
 | |
| Table 3: | Genetic distance (D) (above diagonal) and genetic similarity (below diagonal) between pairs of Rutilus frisii Kutum populations |
 | |
| Table 4: | Analysis of Molecular Variance (AMOVA) based on microsatellite data |
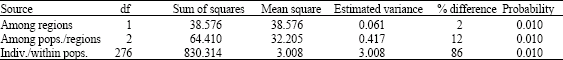 | |
DISCUSSION
Information about genetic variation and population structure of species has pivotal role in their conservation and sustainable use. Despite preparing the majority of kutum stocks from artificial propagation, unfortunately, the knowledge on the molecular genetics and genetic structure of this species is not extensive.
Genetic variability estimates for Rutilus frisii kutum in southern shores of the Caspian Sea (heterozygosity 0.48; alleles per locus 5.4) for these microsatellite loci are comparable with estimated variation in Azarbaijan (heterozygosity 0.48; alleles per locus 5.7) and lower than those reported for most anadromous fish (heterozygosity 0.68; alleles per locus 11.3) by Woody and Avise (2000). Natural spawning grounds of this species has been destroyed due to pollution and its stock has decreased in both Azerbaijan and Iran. It appears that most of restocking of this species by Iranian Fisheries Organization and Azerian hatcheries did not have important role in restocking of this species. To restock this valuable species in the Caspian Sea, annually, more than 150 millions fries up (average weight 1 g) were produced and released into the Caspian Sea. As the hatchery populations used for restocking has been founded with a small effective number of parents (Ne), it is likely that all populations have lost some alleles during the course of restocking. Loss of allelic variation has also been reported for Danish and Polish populations of brown trout (Hansen et al., 2000; Was and Wenne 2002). Reduction in genetic heterozygosity may be also explained by inbreeding and genetic drift. Genetic variability loss recovers very slowly (by mutation or migration). Therefore, suitable genetic management of hatchery fish is an important step to increase the success of stocked fish into the aquatic habitat. Significant deviations from HWE were found at more loci in the Iranian populations than Azeri population. It may explain by presence of null alleles. Also, heterozygote deficiency due to inbreeding between related individuals in the Iranian populations can describe it. It means where homozygote excesses were detected, generally such deviations indicate that factors such as non-random mating, reduction in effective breeding population or specific locus could be under selection pressure were the cause for observed violations (Gilkolaei, 2002). Pairwise genetic differentiation (Fst) was used to assess genetic differentiation, which is the acquisition of allele frequencies that differ among populations . The value of Fst is a useful measure of genetic differentiation among populations and different values mean different variation degrees. Ward et al. (1994) reviewed 7 anadromous fish species and observed Fst estimates with a mean of 10%. In this survey, the Fst value between the Iranian populations was significantly different (p = 0.05), suggesting that all of the Iranian populations are significantly differentiated from one another that can be explained by geographical distance between them (Fig. 1), this should be considered in restocking of this species. Significant population differentiation was observed between the Azeri and the Iranian populations that can also be explained by geographical distance between them. Genetic differentiation can be influenced by a number of evolutionary forces and their interaction that act on natural populations including; migration, random genetic drift, mutation etc. The AMOVA analysis of data also indicates significant genetic differentiation among sampled populations as well as sampled regions (p = 0.01). As releasing a large number of this to Caspian Sea was done by Iranian Fisheries Organization annually and put financial pressure on our country species, differentiation of Iranian samples with Azeri should consider in restocking of this species. The average of Nm was 3.22 between Iranian populations indicates low levels of gene flow that prevent from representing a single panmictic population. The genetic distance between Iranian samples was 0.368 averagely, which indicates that the genetic difference among them is pronounced. Higher genetic distance between Iranian populations, Khoshkrud and Tonekabon, (0.098) than Azeri and Iranian populations were observed (0.078). Shaklee et al. (1982) and Thorpe and Cave (1994) showed that genetic distance values (Nei, 1972) for conspecific populations averaged 0.05 (range: 0.002-0.07), averaged 0.30 for congeneric species (range: 0.03-0.61) and ranged from 0.58 to 1.21 for confamilial genera. The genetic distance between populations in this survey falls within the range of congenerics, suggesting their genetic divergence. In conclusions, this study provides useful information on the levels of genetic variability and differentiation in the Azeri and Iranian populations of Kutum. The comparable number of alleles in both Azeri and Iranian populations was lower than those reported for anadramous fish that should consider in genetic management and conservation programme of this species in Caspian Sea.
ACKNOWLEDGMENTS
This study was supported by Iranian Fisheries Research Institute. We express our sincere gratitude to personnel at the Ecology Research Institute of the Caspian Sea for callabording collaborating with us.
REFERENCES
- Beardmore, J.A., G.C. Mair and R.I. Lewis, 1997. Biodiversity in aquatic systems in relation to aquaculture. Aquacult. Res., 28: 829-839.
CrossRef - Chistiakov, D.A., B. Hellemans and F.A.M. Volckaert, 2006. Microsatellites and their genomic distribution, evolution, function and applications: A review with special reference to fish genetics Aquaculture, 255: 1-29.
CrossRefDirect Link - Croojimans, R.P.M.A., V.A.F. Bierbooms, J. Komen, J.J. Van der Poel and M.A.M. Groene, 1997. Microsatellie marker in common carp (Cyprius carpio L.). Anim. Gen., 28: 129-134.
Direct Link - Woody, J.A.D. and J.C. Avise, 2000. Microsatellite variation in marine freshwater and anadromous fishes compared with other animals. J. Fish Biol., 56: 461-473.
CrossRef - Dimsoski, P., G.P. Toth and M.J. Bagley, 2000. Microsatellite characterization in central stoneroller Campostoma anomalum (Pisces Cyprinidae). Mol. Ecol., 9: 2187-2189.
CrossRef - O'Connell, M. and J.M. Wright, 1997. Microsatellite DNA in fishes. Rev. Fish. Biol., 7: 331-363.
CrossRef - Peakall, R. and P.E. Smouse, 2006. GENALEX 6 genetic analysis in excel population genetic software for teaching and research. Molecular Ecol. Notes, 6: 288-295.
Direct Link - Gilkolaei, S.R., 2002. DNA PCR amplification for species diagnosis of caviar from Caspian Sea Sturgeon. JAST, 4: 40-48.
Direct Link - Thorpe, J.P. and A.M.S. Cava, 1994. The use of allozyme electrophoresis in vertebrate systematics. Zool. Scripta, 23: 3-18.
CrossRef - Turner, T.F., T.E. Dowling, R.E. Broughton and J.R. Gold, 2004. Variable microsatellite markers amplify across divergent lineages of cyprinid fishes (subfamily Leusicinae). Conserv. Gen., 5: 279-281.
CrossRef - Ward, R.D., M. Woodwark and D.O.F. Skinbinski, 1994. A comparision of genetic diversity levels in marine fresh water and anadromous fishes. J. Fish Biol., 44: 213-232.
CrossRefDirect Link - Was, A. and R. Wenne, 2002. Genetic differentiation in hatchery and wild sea trout (Salmo trutta) in the southern Baltic at microsatellite loci. Aquaculture, 204: 493-506.
CrossRef - Hansen, M.M., E.E. Nielsen, D.E. Ruzzante, C. Bouza and K.L.D. Mensberg, 2000. Genetic monitoring of supportive breeding in brown trout (Salmo trutta L.) using microsatellite DNA markers. Can. J. Fish. Aquat. Sci., 57: 2130-2139.
CrossRef