
Md. Mer Mosharraf Hossain
Fish Disease Laboratory, Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Kenji Kawai
Fish Disease Laboratory, Department of Aquaculture, Faculty of Agriculture, Kochi University, Nankoku Shi, Kochi 783-8502, Japan
Journal of Fisheries and Aquatic Science
Year: 2009 | Volume: 4 | Issue: 6 | Page No.: 296-305







ABSTRACT
This study aimed to design to evaluate the immunogenicity as well as the stability of inactivated bacterins in storage conditions to prevent edwardsiellosis in fish species. Three vaccine formulations, formalin (0.4%, FKC), pressure (600 kgf cm-2 for 5 min, PKC) and electric current (100 mA at 12v DC for 5 sec, ECKC) killed bacterin and a routes of administration with intraperitoneal injection (i.p.) was tested. The effectiveness of the immunization strategies was evaluated in terms of Relative Percent Survival (RPS) and antibody levels. On the basis of the results pressure inactivated vaccine via i.p. which confers RPS values over 85% at least 6 months post-vaccination. In the search for a more stable bacterin, inactivated E. tarda antigen was subjected to different storage temperatures. Storage at 4°C did not significantly affect the titer of PKC and antigenic potency remained stable for 6 month. However, with the bacterins FKC and ECKC there is a considerable loss of potency during stored at 4°C. Bacterins were discarded if exposed to temperature of 0°C or below, because the precise freezing point is not established. Bacterins loosed significant potency after 1 month when stored at 25°C.
PDF Abstract XML References Citation
How to cite this article
Md. Mer Mosharraf Hossain and Kenji Kawai, 2009. Stability of Effective Edwardsiella tarda Vaccine Developed for Japanese Eel (Anguilla japonica). Journal of Fisheries and Aquatic Science, 4: 296-305.
DOI: 10.3923/jfas.2009.296.305
URL: https://scialert.net/abstract/?doi=jfas.2009.296.305
DOI: 10.3923/jfas.2009.296.305
URL: https://scialert.net/abstract/?doi=jfas.2009.296.305
INTRODUCTION
Edwardsiella tarda is a Gram negative bacterium of the family Enterobacteriaceae described by Ewing et al. (1965). Edwardsiella tarda is the causative agent of edwardsiellosis and leads to extensive losses in many commercially important freshwater and marine fish worldwide (Plump, 1993).
Over the last decade vaccination has become increasingly important for the prevention of infectious diseases in farmed marine and freshwater fishes (Gudding et al., 1999; Ashida et al., 1999; Kwon et al., 2007; Carrias et al., 2008; Castro et al., 2008). Several attempts have been made to induce protection by immunizing fish against edwardsiellosis with formalin-killed whole cells, cellular lipid and lipopolysaccharides (Mekuchi et al., 1995; Kwon et al., 2006; Lan et al., 2007; Huang et al., 2008; Dumrongphol et al., 2008) producing variable results. With regard to eel, to our knowledge no study about vaccination with pressure inactivated bacterin against E. tarda has been published.
Much study has been done on the inactivation methods of bacteria for medical and food purposes including heat, variety of lipids, high-pressure carbon dioxide, hydrostatic pressure, pulsed electric fields, electric current and so on (Erkmen, 2001; Manolache et al., 2001; Jong-Chul et al., 2003). As fish vaccines are mainly used in aquaculture to treat fish, which are parts of the human diet, safe vaccine inactivation methods should be developed (Chinabut and Puttinaowarat, 2005). Relatively very few research has been performed on the inactivation of fish pathogenic microbes concerning to vaccine preparation. Most of the available fish vaccines, viral and bacterial, are inactivated with formalin (Huang et al., 2008; Dumrongphol et al., 2008). Recently, formalin (formaldehyde) is recognized as a toxic substance which should not be included in human foods or environmental materials. Moreover, formalin is recognized as a non-safe substance which affects the immune responses of host animal (fish) cells to induce early immunoglobulin M response and lower the antigenicity of formalin-inactivated bacterium (Bachmann et al., 1993; DePaola et al., 1995).
In the present study we evaluated and compared the immunogenicity of three vaccine preparations and a route of administration with i.p. were assayed with the aim to establish the effective vaccine with stability that had been maintained storage at 4°C for 6 month just after preparations.
MATERIALS AND METHODS
Bacterial Strains and Media
This study was conducted at Fish Disease Laboratory, Department of Aquaculture, Faculty of Agriculture, Kochi University, Japan. From October, 2006 to April, 2009, E. tarda strain V-1 which is originally isolated from kidney of diseased eel (Anguilla japonica) in Japan. E. tarda strain V-1 was temporally differentiated into serotypes by a cross absorption test of the O-antigen (Tamura et al., 1988). Different serotype strains of E. tarda V-1 strain (Liu et al., 2005) used as the strain for antigen preparation and infection to test the vaccine efficacy against Edwardsiellosis.
Following the recommendations of the European Pharmacopeia (2002), to test the effectiveness of these vaccines, the homologous and the heterologous E. tarda strain HH-1 (isolated from Japanese flounder, Paralichthys olivaceus, 1988 Hiroshima, Japan) were included. LD50 values for V-1 and HH-1 strains in eel were calculated (1.6x101 and 1.2x101 cells mL-1, respectively). Cultures were made on brain heart infusion (BHI, Difco) broth supplemented with 1% NaCl and incubated at 37°C. Stock cultures were maintained frozen at -80°C until used.
Vaccine Preparation
Three types of vaccines were assayed; formalin, pressure and electric current killed bacterin. For the preparation of those vaccines the strain V-1 was grown in BHI for 48 h, obtaining a final concentration of 1x109 cells mL-1. Bacterial cells were inactivated by adding 0.4% (v/v) formalin, pressurized (600 kgf cm-2 for 5 min) using a French press (No. 5501, Ohtake Mfg. Tokyo, Japan) and treated by the current in the range of 100 mA at 12v DC was applied for 5 sec and incubated the treated suspensions at 15°C for 24 h.
Vaccine absorbance (A = 580 nm) was adjusted to 1 (which represents a concentration of 1x109 cells mL-1) with sterile saline solution (NaCl at 0.85%) (SS). Sterility was confirmed by spreading the vaccines on BHI plates and absence of toxicity was tested by the injection of 25 eel with each vaccine.
Fish Vaccination, Experimental Challenges and Measure of the Vaccine Efficacy
Vaccines were evaluated in eel of 102.8±6.6 g (Mean±SD) body weight obtained from an eel farm in Yoshikawa at Kochi Prefecture, Japan. Before vaccination, fish were monitored by bacteriological and serological standard methods (Thoesen, 1994) in order to confirm the absence of pathogens that could interfere in the experiments. Tank water was continuously aerated with an airstone and maintained at approximately 25°C during the experiments.
Fish vaccination was carried out by intraperitoneal injection (i.p.). In the i.p. immunization experiments, vaccines were administered by injection of 0.1 mL of three vaccines, respectively.
Four groups of 100 fish each were employed: group 1 was vaccinated by i.p. injection with FKC; group 2 was vaccinated by i.p. injection with PKC; group 3 was vaccinated by i.p. injection with ECKC and in group 4 was vaccinated by i.p. injection with sterile saline water (control group, CON).
The efficacy of vaccination was confirmed under laboratory conditions by experimental challenges. One month after vaccination three replicas of 10 eel from each group (vaccinated and not vaccinated) were inoculated by i.p. injection with 0.1 mL of a bacterial suspension of the homologous strain V-1 adjusted to a concentration of 104 cfu mL-1. In parallel, challenge trials employing the heterologous strain HH-1 were conducted.
Mortalities were monitored for at least 27 days after the first mortality and all dead fish were examined to confirm the reisolation of the inoculated strain from the internal organs (liver and kidney) by streaking directly onto BHI plates. In addition, in order to determine the possible side effects, including organ adhesions caused by the bacterins, eel were observed internally.
The potency of the vaccines by the i.p. administration was expressed in terms of the Relative Percent Survival (RPS) (Amend, 1981) using the formula:
RPS = {1 – (vaccinated fish mortality % / non-vaccinated mortality %)} x 100% |
The challenge experiments were repeated at 2, 3, 4 and 6 month intervals employing only the homologous strain.
Prior to each challenge, serum-pools (from eight to 10 fish) of each group (vaccinated and unvaccinated fish) were collected in order to measure the antibody levels employing an antibody capture enzyme-linked immunosorbent assay (ELISA) performed as described (Estévez et al., 1994), employing peroxidase-conjugated goat anti-rabbit IgG (Gaithersburg MD 20878-USA) and a rabbit anti-eel serum. The optical density at 492 nm was measured with an ELISA reader (Thermo Electron Corporation). Moreover, eel were weighed (10 fish per group) at 0, 1, 3 and 6 months after vaccination to determine the possible influence of vaccination on the growth rate of fish.
Evaluation of Bacterial Protein Stability
Pre-storage and after 6 month post storage, the serum agglutinating antibody titer against E. tarda FKC, PKC and ECKC was determined by the microtiter method (Nowotny, 1969) with incubation for 2 h at 25°C and overnight at 4°C. Protein electrophoresis was performed by using the method of Laemmli (1970).
Statistical Analysis
To evaluate the significance of the differences between protection results obtained with each group, the student’s t-test was used. The data of ELISA were represented as Means±SD and analyzed by one-way Analysis of Variance (ANOVA) and Tukey’s test was used to compare the mean values between individual treatments (Zar, 1984) using Stat plus 2007 professional. Values were considered significantly different at p<0.05.
RESULTS
Efficacy of Vaccines 1 Month Post-Vaccination
In the present study we studied the protection against edwardsiellosis conferred by three different vaccine formulations administered by i.p. to eel. The results obtained in this study showed that 1 month post-vaccination the protection conferred in eel against E. tarda varied depending on the type of vaccine. While the vaccination of ECKC bacterin via i.p. showed ineffective for the protection of eel against edwardsiellosis. In fact, the evolution of mortalities of this group and its respective control (group 4) followed a similar kinetics (Fig. 1) resulting in a RPS value of 10.13% at the end of the experimental period (Fig. 3). Moreover, when the ELISA assay was conducted, antibody levels in the eel sera of group 3 were 0.045, near to the group 4 (control group) (Fig. 2). Based on these data, no more experimental challenges were performed with group 3 fish in subsequent months.
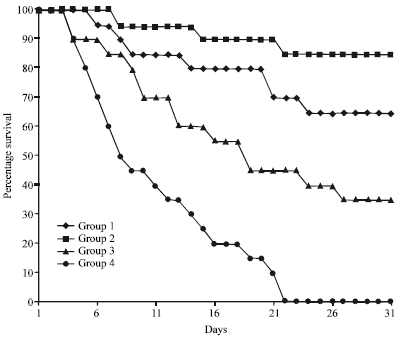 | |
| Fig. 1: | Percent survival obtained in experimental challenge 1 month post-vaccination (i.p.). Group 1, vaccinated with FKC bacterin; group 2, vaccinated with PKC bacterin; group 3, vaccinated with ECKC bacterin; group 4, control fish vaccinated with sterile saline water |
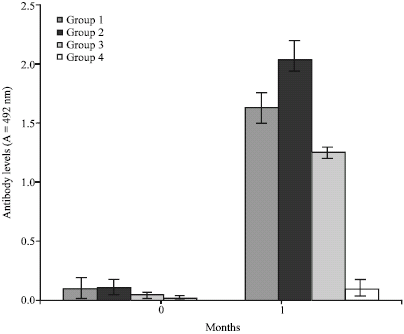 | |
| Fig. 2: | Specific antibody titers in eel 1 month after i.p. vaccination with different vaccines. Group 1, vaccinated with FKC bacterin; group 2, vaccinated with PKC bacterin; group 3, vaccinated with ECKC bacterin; group 4, control fish vaccinated with sterile saline water. Bars indicate standard deviations |
In the case of groups 1 and 2, immunized by i.p. with the FKC bacterin and the PKC vaccine, respectively, the highest percent of fish survival was found in the group 2, injected with antigen with 15% mortalities registered in this group (Fig. 1) which represents a 83.13% RPS (Fig. 3).
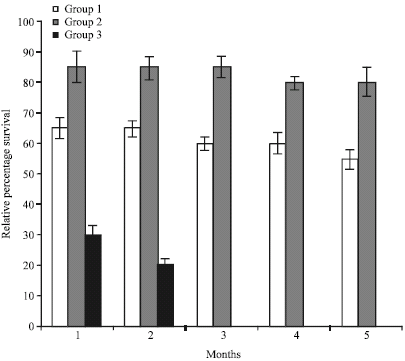 | |
| Fig. 3: | Relative Percent Survival (RPS) obtained in the different experimental challenges. Group 1, vaccinated with FKC bacterin; group 2, vaccinated with PKC bacterin; group 3, vaccinated with ECKC bacterin; group 4, control fish vaccinated with sterile saline water. Bars indicate standard deviations |
The RPS in the fish receiving FKC bacterin (group 1) was of 59% (Fig. 3).
In the ELISA assay, the antibody levels of group 1 and 2 presented values of 1.107 and 1.546, respectively, representing a significant increase with regard to the values displayed by the unvaccinated groups 4 (Fig. 2). Although both the FKC and PKC vaccines administered by i.p. conferred high levels of protection in terms of RPS and antibody levels, when these values were statistically analyzed, significant differences (p<0.05) were obtained with PKC vaccines.
With regard to the challenge experiments employing the heterologous E. tarda strain HH-1, similar RPS and antibody level values as those showed to the homologous strain were obtained.
Duration of the Protection
Four additional experimental challenges at 2, 3, 4 and 6 months post-vaccination were performed to determine the duration of the protection conferred to eel i.p. immunized with the three different vaccine formulations. Data obtained in these challenges are shown in Fig. 3 and 4.
Stability of Inactivated Vaccine at Different Storage Temperature
Representative samples of sera from eel immunized with three bacterins were tested after storage at 4 and 25°C for 6 month. The resulting serum agglutination titers are reported in Table 1 and are compared with the titers of different vaccine prepared from E. tarda. Evidence of the stability of the vaccine is demonstrated by the residual antigenic capacity shown. Aside from the magnitude of the serum-agglutination titers for the PKC bacterin being higher than FKC or ECKC. The serum agglutination titers were not demonstrated after storage at 25°C for 6 month with tested bacterins, that indicates antigenic potency or stability with these bacterins were loosed.
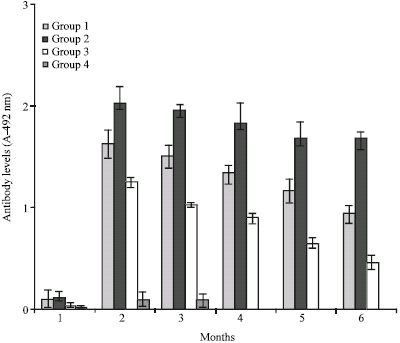 | |
| Fig. 4: | Evolution of specific antibody titers in eel after i.p. vaccination with the different vaccines. Group 1, vaccinated with FKC bacterin; group 2, vaccinated with PKC bacterin; group 3, vaccinated with ECKC bacterin; group 4, control fish vaccinated with sterile saline water. Bars indicate standard deviations |
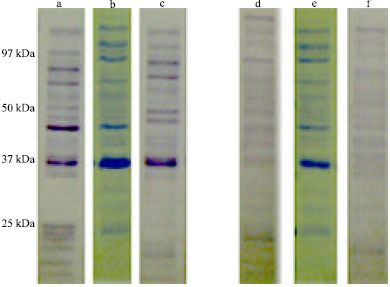 | |
| Fig. 5: | Sodium Dodecyl Sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) of three bacterin samples (a) FKC, just after preparation (b) PKC, just after preparation (c) ECKC, just after preparation (d) FKC, just after preparation, stored for 6 month at 4°C (e) PKC, just after preparation, stored for 6 month at 4°C and (f) ECKC, just after preparation, stored for 6 month at 4°C. Molecular weight markers in kDa are shown at the left margin |
In the SDS-PAGE of the total protein extracted from all inactivated conditions of E. tarda V-1 strain, many proteins were detected in each inactivation conditions in Fig. 5 (lane a, b and c).
| Table 1: | Effect of storage at 4°C on antigenic potency of inactivated E. tarda vaccine after storage for 6 month |
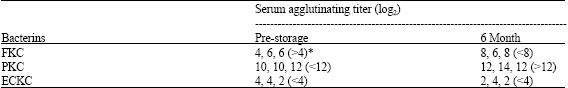 | |
| *Values in parentheses represent the geometric mean | |
Several discrete bands over a wide range of molecular weights can be seen in the PKC (lane e) vaccine that had not been FKC and ECKC (lane d and f). Protein bands have disappeared or could not be distinguished in FKC and ECKC sample that had been stored for 6 month at 4°C. Moreover, a major protective antigen at 37 kDa was strongly detected by SDS-PAGE with PKC after stored for 6 month at 4°C.
DISCUSSION
Fish vaccinated with the FKC bacterin, showed high levels of RPS (64.33%) at 1 months post-immunization. However, this value decreased in the following challenge experiments. Thus, in month 3 post-vaccination, the RPS value decreased to 57.53% and was only 54.63% 6 months post-vaccination (Fig. 3).
In the case of the PKC vaccine, the RPS obtained with this group of eel revealed that this vaccine conferred a high degree of protection for at least 6 months after its administration. The RPS values were maintained among 80-85% during all the experimental period (Fig. 3). These values are in agreement with those recommended by the previous observation (Hamaguchi and Kusuda, 1989; Vinitnantharat and Plumb, 1993; Ashida et al., 1999; Kwon et al., 2005, 2007; Swain et al., 2007; Castro et al., 2008).
With regard to the antibody levels in eel sera, the PKC vaccine always displayed statistically significantly (p<0.05) higher values than those obtained with the FKC and ECKC bacterin (Fig. 4). Although, with all vaccines a progressive decrease in the antibody levels was detected along the experimental period, with the PKC vaccine these values remained constant from month 4 (antibody levels around 1.67).
On the other hand, the immediate alteration in the biochemical profile determined by electrophoresis seems to be much more dramatic. The ECKC sample (Fig. 1b, c) had an almost complete absence of the bands, suggesting extensive protein degradation, nearly similar result seen in FKC sample. This observation could be associated with the reduced immunogenicity of the FKC and ECKC vaccine. Taken together, these results do not encourage the storage at 4°C as a means for preserving the immunogenic and biochemical properties of this vaccine. The all bacterins sample had a complete absence of the bands or complete protein degradation seen after storage at 25°C for 6 month.
It was encouraging that PKC gave a better protection than other vaccines in the in vivo experiments-which may be due to the preservation of antigenicity by pressure. However, another possibility is that quantity of the protective antigens 37 kDa (Kawai et al., 2004; Tu and Kawai, 1999) in this preparation is higher, destroyed less of the protective epitopes, particle structure between the vaccines, mechanisms of inactivation (Taisuke et al., 2003) and antigen form (Schirmbeck et al., 1995).
In summary, an effective PKC vaccine against eel edwardsiellosis was developed which confers levels of protection higher than 85% for at least 6 months post-vaccination. Although, this high protection could later decrease gradually with time, it can be assumed that the RPS levels will remain sufficiently (p<0.05) high to protect eel against edwardsiellosis during all the production cycle. In fact, this inactivated vaccine is currently being employed by i.p. in an eel rearing facility as an autovaccine and, to our knowledge; no outbreaks of edwardsiellosis were detected in this facility during the eel rearing period.
Present findings have demonstrated that an inactivated PKC vaccine can be stable and is able to stimulate an antigenic response in eel against E. tarda at least 6 month after the vaccine has been prepared. A PKC bacterin, prepared from pressure treated E. tarda V-1 strain was stable for 6 month at 4°C. Vaccine stored under these conditions stimulated an antigenic response in eel which was significantly different from that of formalin and electric current prepared vaccine. The magnitude of serum-antibody titers was higher when vaccine compared with FKC and ECKC.
The ability of the PKC bacterin to induce specific cell-mediate immunity should be investigated to analyze total immune responses of eel against E. tarda infection and to develop a commercial product in the fish immunization calendar (Toranzo et al., 1997).
CONCLUSION
In conclusion, the efficacy of PKC vaccine against eel edwardsiellosis sustained its protective antigenicity after storage at 4°C for 6 months. This result demonstrates that the inactivated PKC vaccine is stable and being able to stimulate protective immune responses in eel against E. tarda at least 6 month after the vaccine has been prepared.
ACKNOWLEDGMENTS
We would like to thank the staff and students of Fish Disease Laboratory, Faculty of Agriculture, Kochi University for their help in the field.
REFERENCES
- Ashida, T., E. Okimasu, M. Ui, M. Heguri, Y. Oyama and A. Amemura, 1999. Protection of Japanese flounder Paralichthys olivaceus against experimental edwardsiellosis by formalin-killed Edwardsiella tarda in combination with oral administration of immunostimulants. Fish. Sci., 65: 527-530.
CrossRefDirect Link - Castro, N., A.E. Toranzo, S. Nunez and B. Magarinos, 2008. Development of an effective Edwardsiella tarda vaccine for cultured turbot (Scophthalmus maximus). Fish Shellfish Immunol., 25: 208-212.
Direct Link - Chinabut, S. and S. Puttinaowarat, 2005. The choice of disease control strategies to secure international market access for aquaculture products. Dev. Biol., 121: 255-261.
Direct Link - DePaola, A., J.T. Peeler and G.E. Rodrick, 1995. Effect of oxytetracycline-medicated feed on antibiotic resistance of gram-negative bacteria in catfish ponds. Applied Environ. Microbiol., 61: 2335-2340.
PubMedDirect Link - Estevez, J., J. Leiro and A.E. Toranzo, 1994. Role of serum antibodies in protection of vaccinated turbot (Scophthalmus maximus) against vibriosis. Aquaculture, 123: 197-204.
Direct Link - Gudding, R., A. Lillehaug and O. Evensen, 1999. Recent developments in fish vaccinology. Vet. Immunol. Immunop., 72: 203-212.
CrossRefDirect Link - Kawai, K., Y. Liu, K. Ohnishi and S.I. Oshima, 2004. A conserved 37 kDa outer membrane protein of Edwardsiella tarda is an effective vaccine candidate. Vaccine, 22: 3411-3418.
CrossRefDirect Link - Kwon, S.R., E.H. Lee, Y.K. Nam, S.K. Kim and K.H. Kim, 2007. Efficacy of oral immunization with Edwardsiella tarda ghosts against edwardsiellosis in olive flounder (Paralichthys olivaceus). Aquaculture, 269: 84-88.
CrossRefDirect Link - Kwon, S.R., Y.K. Nam, S.K. Kim, D.S. Kim and K.H. Kim, 2005. Generation of Edwardsiella tarda ghosts by bacteriophage PhiX174 lysis gene E. Aquaculture, 250: 16-21.
Direct Link - Kwon, S.R., Y.K. Nam, S.K. Kim and K.H. Kim, 2006. Protection of tilapia (Oreochromis mosambicus) from edwardsiellosis by vaccination with Edwardsiella tarda ghosts. Fish. Shellfish Immunol., 20: 621-626.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lan, M.Z., X. Peng, M.Y. Xiang, Z.Y. Xia and W. Bo et al., 2007. Construction and characterization of a live, attenuated esrB mutant of Edwardsiella tarda and its potential as a vaccine against the haemorrhagic septicaemia in turbot, Scophthamus maximus (L.). Fish. Shellfish Immunol., 23: 521-530.
CrossRefPubMedDirect Link - Liu, Y., S. Oshima, K. Kurohara, K. Ohnishi and K. Kawai, 2005. Vaccine efficacy of recombinant GAPDH of Edwardsiella tarda against Edwardsiellosis. Microbiol. Immunol., 49: 605-612.
Direct Link - Schirmbeck, R., W. Bohm, K. Melber and J. Reimann, 1995. Processing of exogenous heat-aggregated (denatured) and particulate (native) hepatitis B surface antigen for class I-restricted epitope presentation. J. Immunol., 155: 4676-4684.
Direct Link - Swain, P., A. Behura, S. Dash and S.K. Nayak, 2007. Serum antibody response of Indian major carp, Labeo rohita to three species of pathogenic bacteria; Aeromonas hydrophila, Edwardsiella tarda and Pseudomonas fluorescens. Vet. Immunol. Immunopathol., 117: 137-141.
Direct Link - Taisuke, W., F. Soichi, H. Junichi, K. Tetsuya, O. Hirokazu and Y. Makari, 2003. Inactivation of geobacillus stearothermophilus spores by high-pressure carbon dioxide treatment. Applied Environ. Microbiol., 69: 7124-7129.
CrossRefDirect Link - Tamura, K., R. Sakazaki, A.C. McWhorter and Y. Kosako, 1988. Edwardsiella tarda serotyping scheme for international use. J. Clin. Microbiol., 26: 2343-2346.
PubMedDirect Link - Vinitnantharat, S. and J.A. Plumb, 1993. Protection of channel catfish Ictalurus punctatus following natural exposure to Edwardsiella ictaluri and effects of feeding antigen on antibody titer. Dis. Aquat. Org., 15: 31-34.
Direct Link - Huang, Y., H. Lou, X. Wu and Y. Chen, 2008. Characterization of the BPI-like gene from a subtracted cDNA library of large yellow croaker (Pseudosciaena crocea) and induced expression by formalin-inactivated Vibrio alginolyticus and Nocardia seriolae vaccine challenges. Fish Shellfish Immunol., 25: 740-750.
PubMed - Dumrongphol, Y., T. Hirota, H. Kondo, T. Aoki and I. Hirono, 2008. Identification of novel genes in Japanese flounder (Paralichthys olivaceus) head kidney up-regulated after vaccination with Streptococcus iniae formalin-killed cells. Fish Shellfish Immunol., 26: 197-200.
CrossRef - Carrias, A.A., J.S. Terhune, C.A. Sayles and J.A. Chappell, 2008. Effects of an extended hatchery phase and vaccination against enteric septicemia of catfish on the production of channel catfish, Ictalurus punctatus, fingerlings. J. World Aqua. Soc., 39: 259-266.
CrossRefDirect Link