
A.M. Ali
Faculty of Environmental Sciences and Marine Biology, Hadhramout University of Sciences and Technology, Mukalla, Yemen
A.A. Basmidi
Faculty of Environmental Sciences and Marine Biology, Hadhramout University of Sciences and Technology, Mukalla, Yemen
M.Sh. Aideed
Faculty of Environmental Sciences and Marine Biology, Hadhramout University of Sciences and Technology, Mukalla, Yemen
Al-Quffail A. Saeed
Faculty of Environmental Sciences and Marine Biology, Hadhramout University of Sciences and Technology, Mukalla, Yemen
Journal of Fisheries and Aquatic Science
Year: 2009 | Volume: 4 | Issue: 5 | Page No.: 210-227







ABSTRACT
Abalone has been exploited by humans around the ocean coasts of the world. On contrary, it is still neglected in the Gulf of Aden. This study is a first attempt to explore the biological and ecological status of the absolutely unexploited abalone Haliotis pustulata in the Aden Gulf, Yemen. Between Jan 2004 and June 2006, about 700 abalones were collected and analyzed from Intertidal and sublittoral zones along 6 sites of Hadhramout coast. It is revealed that the studied species is relatively small, measuring up to 62 mm, with a slow growth rate of 4-10 mm year-1 and has moderate fertility compared with many other species in other localities. The highest GSI as well as the highest proportion of mature Sufaylah were prevailed from March to May. Morphological traits, spread, age-size structure and reproduction behavior are discussed in the light of the absence of local similitude studies and well documented world wide haliotids. Results call for necessity of more detailed studies.
PDF Abstract XML References Citation
How to cite this article
A.M. Ali, A.A. Basmidi, M.Sh. Aideed and Al-Quffail A. Saeed, 2009. First Remarks on Abalone Biology (Haliotis pustulata) on the Northern Coast of Aden Gulf, Yemen. Journal of Fisheries and Aquatic Science, 4: 210-227.
DOI: 10.3923/jfas.2009.210.227
URL: https://scialert.net/abstract/?doi=jfas.2009.210.227
DOI: 10.3923/jfas.2009.210.227
URL: https://scialert.net/abstract/?doi=jfas.2009.210.227
INTRODUCTION
Haliotis is solely genus of family Haliotidae, one of the more reputable marine organisms from molluscs. It is highly appreciated for its high quality of meat and delightful shells nacreous luster. Abalone culture comes to be one of the largest growth industries in the worlds seafood sectors in addition to achieved successes in its stock enhancement, sea ranching and habitat improvement (Gordon and Cook, 2004; James et al., 2007). This, beside the recent practices of implantation and cultivation of pearl under its mantle to produce more precious pearls than those of oysters (Fankboner, 2002).
Abalone's life history was well studied for centuries in the Far East, Australia, North America and Europe; even that, seemingly, in many countries, in particular Japan and China, its cultural production exceeding its landing from wild fishery (Karpov et al., 1999).
Although, Arab divers, from immemorial times took an interest in marine shells, especially pearl oysters; meanwhile they ignore the abalone ones, actually that because pearl oysters brought them the traditional valuable pearls. Thus abalone, for long time, somewhere up-to-date, had been forgotten bottom crawling in the coast of Arabian Peninsula.
Nowadays, except in Omani waters where the abalone is well gathering, highly estimating, thoroughly studied and commercially cultured (Shepherd et al., 1995; Gindy, 1999; Benny et al., 2003); very little is known about haliotids in Arabian coasts, particularly in Yemeni coasts. Here, it is still disregarded in fisheries, although it is moderately existing and maybe regionally abundant. The sole place, where abalone, Haliotis pustulata, or Sufaylah as it is known locally, very occasionally and singly harvested, is the Intertidal Zone of Socatra Island; here seemingly two species were noticed: H. pustulata and H. mariae (Shepherd and Brown, 1993; Geiger, 2000). The latter notices confined to just repetition their occurrence, without any details in ecology or biology.
Due to the absence of authenticated information and voucher material for H. pustulata from Aden Gulf, it becomes rightfully to consider the present description as first report of occurrence and ecology of this species in the specified region. What can support that conclusion is an absence of documented natural range of this species, disregarding abbreviated notes of respected authors (Shepherd and Steinberg, 1992; Geiger, 1996, 2000).
Although, Sufaylah has no commercial importance in North coast of Aden Gulf yet, there are evidences that it moderately existing. It makes this study a first attempt to throw the light on, gain some knowledge on biological characteristics and pay attention of scientists to more detailed studies on this species.
MATERIALS AND METHODS
Study Area
The Hadhramout coast of Aden Gulf-Yemen, as for our present survey, lies between Beer Ali district with its Islets: Sikha, Halania and Bureiqa-N13°50' E48°14'. This site is located approximately 150 km South-Westwards of Mukalla City. Survey is prolonged up to Ras Sharma (Sharma cape)- N14°50' E51°10´-about 150 km Eastwards of Mukalla City (Fig. 1). The coast line is characterized by wide variety of its beach and bottom composition or construction. Most sections of this coast have relatively wide stretch of Intertidal zone; some has been completely represented as exposed Intertidal rocky shelf, which is substituted, as being more deeper 5-20 m, by consecutive hard igneous rock platforms-bedrocks and some were coralline complexities. The shallow sub-littoral formations often pervaded by many fissures, clefts, crevices, ledges and boulders.
In spite of vulnerability of the diverse ecosystem of Aden Gulf to industrial pollution and other human activities last time, this coast is influenced by the summer South-Western monsoon, that has been prevailing during the period from June to September, producing renewed a wealthy and intense Upwelling phenomenon. Latter conditions make the coastal water of the meant region subject to relatively wide temperature variation. As a result, while the coastal water is strongly related to the prevalent tropical climate and remained warm (25-30°C) nearly most of the year-October up to mid June next year, it rapidly decreases to 18-16°C in last days of July-August and sometimes, even to 12°C as it happened in the end of July of 2005.
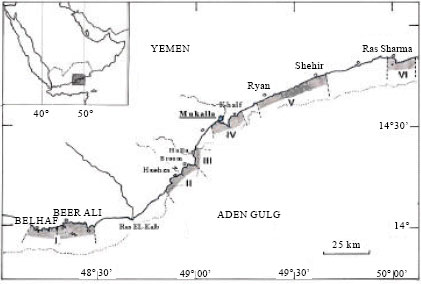 | |
| Fig. 1: | The study area and sampling locations: the shaded areas show the investigated sites with Roman numerals that detailed in the text |
At this time the upwelled water overspread the coast. Water salinity of the study coast ranged between 35.5 and 36.7 g L-1 (our water quality data at 4-5 m depth for 2004-2005).
It must be understood that we are dealing with a much disguised, slowly moving, relatively sedentary and ecologically sensitive Intertidal gastropod dweller. These basic facts make it easy to lessen the area of selected sites along Hadhramout coast and then snorkelling were carried out in restricted and randomly defined sectors. These sectors we included in six main sites with some adjoining, arranged from South-West to East (Fig. 1): I-Balhaf-BeerAli; II-Hsehsa-Broom; III-Halla; IV-Khalf Mukalla shore, V-Ryan and Shehir and VI-Ras Sharma.
Survey prolonged regularly on monthly base from the mid of January 2004 up to mid of June 2006. Seven hundred and eight individuals of abalone were collected from different places of each site from exposed rocks at intertidal and sublittoral zones up to 15 m depth.
Field investigations were carried out monthly by snorkeling around subjected fixed station by a Global Positioning System (GPS) and across the advance detected bottom area by fixed visual marks. After each spawning season and along the following month, deeper bottoms (20-40 m) of some sites were investigated by scuba diving for justifying abalone occurrences (expected settling) in these depths, it gave absolutely negative results yet.
Observations of the behaviors of Haliotis pustulata Reeve had been performed thrice: at night during November 7 in 2004; repeated at night of July 28, 2005 and December 5, 2005. The water temperature at sites had been 21, 17 and 26°C, respectively. The accidentally chosen specimens for observations were within restricted shell length, which had been 38-41 mm.
All gathered samples freshly measured and weighed to the nearest 0.1 cm and 0.01 g, then dissected, defined and separated by sex at the laboratory.
The age was determined by very well apparent interrupted growth checks on the shell surface, which in some shells were very well visible nacreous ridges (Fig. 2b). It is also partially affirmed upon the well filing shell's spires by polishing with fine grit wet-and-dry paper, following McShane and Smith (1992) and Shepherd et al. (1995). The shells and their sections were examined under a microscope. Alternate dark and light growth bands are easily seen and used to estimate age; then they have been successfully used to estimate the annual growth of the organisms. Age in years is considered the number of growth checks which has been verified from a special experiment. In this experiment a 10 abalone were measured, tagged and placed back in the V-Khalf Mukalla site at a depth of about 5 m in 15th of April 2005, to grow naturally. An attempt was made to estimate the growth monthly. Only 4 of 10 tagged specimens are found regularly each month and are measured up to March, 2005 (others were found irregularly up to date). The initial shell lengths of recovered abalones were between 30.8, 30.9, 32 and 38.1 mm.
Due to complete absence of commercial and recreational catch of abalone and due to lack of sexual dimorphism, the size composition and growth estimation were based on total specimens collected during the entire study period. Because of the samples from only 3 adjacent sites-II, III and IV, we had recourse to consider the mixed populations from all areas and of mixed sexes concerning actual growth rate and morphometric relationships between shell length, height, width, total weight and weight of animal without shell.
The relationship between these features in addition to length-weight relationship was studied using regression analysis. length and Weight data for Broom-Hsehsa (II), Halla (III) and Mukalla (IV) sites were fitted to the power equation separately (Whole weight = a length b) then as whole sample, where a and b are parameters of the power relationship (Fig. 6).
 | |
| Fig. 2: | The (a) body and (b) shell appearance and size of H. pustulata |
For reproduction, gonads of 668 individuals were studied. From each fresh animal gonad were carefully and scrupulously removed, examined and distinguished, sometimes microscopically. Each gonad weighed to nearest 0.01 g then preserved in 6% formalin. Accordingly, the gonad semblance, shape, features and colours were used to determine the sex and maturity stages which expressed here following Martin et al. (1983), Wells and Keesing (1989), Litaay and De Silva (2001) and Najmudeen and Victor (2004).
| • | Stage I: Juveniles and immature individuals: the gonads of very small size and invisible to naked eyes are still in phase of somatic growth. It is characterized by small gonadal bulk |
| • | Stage II: Premature, where rapid increase in number and size of gamete cells occurs. The gametes become larger and take shape of pears. In this stage it is still partially difficult to differentiate sexes; although the ovaries become fleshier than the testis and little pinkish. Animal with latter stage could not be considered as mature in aspect of maturation size and age |
| • | Stage III: Mature stage, the gonads notably enlarge due to maturation of oocytes and spermatozoa. The ovaries become more granulated and displaying well differentiated greenish colour, whereas testis filled up with spermatozoa and creamy, white or beige semen fluid, which gives them a distinct pattern. In this stage, the ovaries and testes with accumulated mature and liberated oocytes and spermatozoa. Here the gonads very clearly are observed under the foot or mantle without dissecting |
| • | Stage IV: Ripe, ovaries are dark greenish, covering the most visceral area and is fully surrounding the hepatopancreas capsule. The outwardly manifest ripe oocytes here are so large and tight that they becoming polygonal. In this stage, the male gonad lobes apparently filled with ivory spermatic fluid. Oocytes in this stage assumed to be fully vitellogenic (Tutschulte and Comnell, 1981; Campbell et al., 2003). This stage can be described as partially spawned in some individuals |
| • | Stage V: Spawning, the gonads engorged with fully matured gametes in this relatively short temporary stage. Factually, it is a short period of liberation of ripens gametes or gametes just before releasing. In this stage the ripe gametes released under any light pressure |
| • | Stage VI: Spent, this stage might be considered as already spawned. It is the period in which the gonads state was just after spawning and the beginning of the gonad recovery. The wall of gonad seems so wrinkly due to discharging most ripen eggs and sperms. Very little unovulated mature eggs and sperms remain in gonad compartments. The gematogeneses (for short time) cease and gonads obviously and greatly reduced in size and become yellowish or creamy in color, but not far as that the animal sex cannot be distinguished; contrarily, in/or after this stage the sexes become persistently apparent |
To study the reproductive cycle, the estimation of widely used in abalone, gonadosomatic index (GSI) was done through wet weight of accurately removed gonad and calculated as a percentage to wet body weight (Litaay and De Silva, 2001, 2003; Awaji and Hamano, 2004).
The sex ratio, initial size at sexual maturity, spawning period and fecundity of the H. pustulata were studied on base of sexual condition by visual inspection of the staged gonad.
The size at sexual maturity was determined as average for a population as the size at which 50% of a representative sample shows mature gonad development (Nash et al., 1994; Tarbath, 1999; Tarbath and Officer, 2003). Two millimeter size class determines size at 50% sexual maturity.
The fecundity was calculated directly from randomly chosen and carefully dissected matured female gonads that were at the ripe stage. These 54 ovaries precisely weighed to nearest 0.01 g, then; from three sections of horn-shape ovary a sub sample was taken from the base of horn, mid part and the apical part of horn. The sub sample was weighed to nearest 0.001 g and fixed in formalin as well as ovary it self. After few days all eggs in sub sample were disjointed, carefully counted under a magnifier or binocular microscope and recalculated as total number of matured oocytes in the entire ovary what was defined as actual fecundity (Campbell et al., 2003).
The total gathered, on monthly based, specimens were 708 individuals: (1) 228 from Broom-Hsehsa; (2) 342 from Halla and (3) 138 specimens from Khalf-Mukalla. The monthly percentage of the gonad development stages were determined the yearly pattern of spawning. All obtained data were statistically treated.
RESULTS
Description and Morphology
The distinctive characteristics of emergent Abalone, Haliotis pustulata in Northern Aden Gulf coast is relatively of moderate size, reaches up to 62 mm.
The collected shells were mainly epiphytic free, thin, flattened and had moderate weight (31.7 % of total b.wt), in addition to the absence of scare muscle attachment in the interior of the shell. The shell is well ellipse or oval shaped with clearly defined smooth ridges, slightly convex on folded right-Columella side and moderately convex on the loose left sides. The spiral cord is moderately strong, loosely narrow coiled and general sculpture with fine spiral ribs and prominent growth check. Spiral series had mainly 6-8 (92%), rarely five and nine open holes, which ends with closed holes along the spire of the left side of the shell. The open holes are somewhat rimmed, but not elevated above surface (Fig. 2b).
The shell colour of predominant alive specimens is dark brown with greenish tint, tending to be light brownish with mottled orange and whitish tan in adult specimens. Very few specimens are brick red with pale patches when it crops out. The shell surface lost this colour pattern to be reddish or light brown, slowly fad away after discarded from the water. Shell interior is very smooth and iridescent, having deep violet, blue and green luster combined with main pearly and silvery ground.
The foot sole has bright yellow colour with yellowish to beige epipodium surface. The epipodium have scalloped tuberculated edges that give it a slightly frilled shape. Epipodial tentacles are yellowish beige, short, thick and very slightly projected.
Spreading and Behavior
Onset, the result of our investigations emphasized the fact of abalone rarity in Southern coast of Yemen, which, seemingly led to the prevailing historical absence of its commercial and coastal fishing. One of the prominent evidences was that, three of six chosen study sites: I-Balhaf-Beer Ali, V-Ryan (Shiheir) and Shehir and VI-Sharma cape, are completely free from abalone occurrence according to our investigations, in spite of well developed macroalgae assemblages and prevalence of the moderate and big boulders covered by crustose coralline algae within rocky shore.
Haliotis pustulata juveniles preferred hidden habitats (cryptic), mainly under the rocks, boulders, narrow clefts and cracks. The juveniles were mainly found at 3-4 m depth. The 35-55 mm sizes prefer the depths between 0.5-3 m. In general, the distribution of individuals gradually occurred from 0.5 m depth, which occupied by adults and stretched to 10-15 m where the juveniles spread; this might be due to the active surf where the organisms have been subjected to wave interference in the intertidal zone.
Although, home scars in majority of the sampling areas were present, suggesting that some Sufaylah had stayed in the same spot for a long time; as we directly observed, Sufaylah lived in settled life within preferable habitats, with relatively short distances of movement that happened, mainly at night, which is considered as feeding time. Moreover, it was moving within a very restricted light and area not more than 4-6 m upon 6 months (special observations, 2005).
Observations showed that there was an assemblage of organisms that were providing a camouflage making Sufaylah disguised from predators during its nocturnal feeding. At daytime, it was often hiding between the sea urchins, which provided a shelter for its juvenile.
Latter behavior was noted occasionally in situ, where the special observations were demonstrated the nocturnal reaction of Sufaylah against the moon and searchlight. At moon shining time through new lunar month, when it was shining for less than 3 h after sun set, Sufaylah does not appear from its refuges under the crevices, caves, cavities and boulders. It emerges, for searching and feeding, after moonset and darkness prevailed. At this time, the organism’s excursions have been within a relatively large area, but it returned to its refuges before dawn. When the moon is shining for a long time, Sufaylah obliged to moves out for nocturnal feeding excursions, but it inclined to shaded sides of the feeding substrates to which the moon light did not reach. At this time Sufaylah does not leave away from their places; may be to less prone to predators.
Likewise, when we directed the searchlight to feeding organism for few minutes, when it was above the boulders at the dark nights, it rapidly turns away from the luminous side to shaded one.
These observations give an emphasis that H. pustulata has a strong sensitivity against light and darkness.
Size, Age Structure and Growth
Due to scarcity of samples from some sites, the morphometric indices were recorded as to all study samples. The main results of this aspect is shown in Fig. 3a and b. The relationship between body length and shell length and body length and shell width was adequately firm; they were: Y = 1.05 X+1.3945 (R2 = 0.94) and Y = 0.78 X - 0.74 (R2 = 0.93), respectively.
Length and age frequency distributions were studied for Northern Aden Gulf population (Fig. 4, 5). Figure 4 shows the predominance sizes for the length groups were between 36-48 mm, with apparent prevailing of 36-38 and 42-46 mm groups, which reflect the fact of prevalent of the Sufaylah of Halla site that would have almost firstly matured individuals. Furthermore, it is found that the males were almost preponderate after juveniles' sizes- up to 22 mm, then, only after 42 mm shell length the females somewhat predominate.
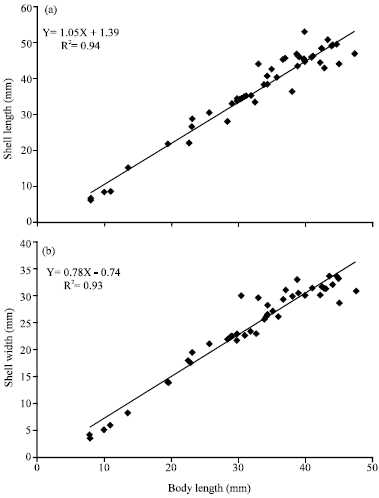 | |
| Fig. 3: | The relationship between (a) body length and shell length and (b) body length and shell width of H. pustulata |
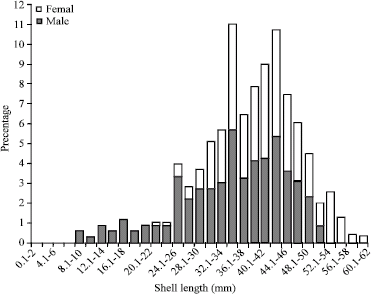 | |
| Fig. 4: | Length distribution of H. pustulata in Aden Gulf Coast, considering the percentages of males and females |
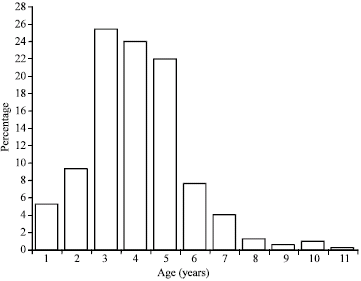 | |
| Fig. 5: | Haliotis pustulata age distribution in Aden Gulf Coast |
The mean shell total length, of Sufaylah was 39.91 mm (SE = 0.28, N = 668); it varied significantly among the more abundant three sites-III (Halla), IV (Mukalla) and II (Broom): The Halla Bay population has a larger individuals (40.76 mm, SE = 0.29, N = 342) than Broom-Hsehsa shore (38.85 mm, SE = 0.14, N = 228) and Mukalla sites (38.86 mm, SE = 0.36, N = 138), whereas SE is standard error and N is the number of animals. The higher proportion of large individuals at Halla Bay (the highest density location) suggests the suitability of this site as spawning ground that is well demonstrated by predominance of initially matured individuals. In general population the mean male and female shell length were 40.1 and 42.1 mm, respectively.
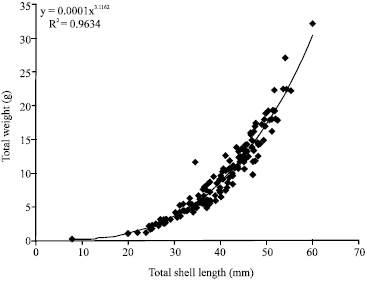 | |
| Fig. 6: | The shell length-weight relationship of H. pustulata, from Aden Gulf Cost (general sample) |
This peculiarity had taken place in the age case too, where the predominance was mainly to age group 3 years, which followed by 4 and 5-year groups (Fig. 5). Whereas the latter 3 groups consist more than 70% of the sample, we find only nine specimens having 9-11 years old-less than 2% of samples.
Concerning shell length-weight relationship for Sufaylah, it appears that (b) values was very close to 3 in most abundance sites: Broom-Hsehsa Y = 0.00006 X 3.20, Mukalla Y = 0.00009 X3.08 and Halla-Y = 0.0001 X 3.09, also (a) is very small, <0, 0001. Although, a relatively large sample is used (n = 708), of all sites together, to determine the relationship between length and weight, we find that W = 0.0001 x 3.1 (Fig. 6). Thus, growth is isometric (b around 3).
The result of the special tagging experiment accidentally reveals that, in addition to apparent one yearly growth ring, the yearly growth rate were been 7.8, 8.3, 10.2 and 10.8 mm for 4 recovered tagged specimens. These give some confidence to the results concerning age and growth estimations.
It is found that growth rates of Sufaylah were high initially and size-dependent for about the first 4 years during which shell length was a good predictor of age. Thereafter, growth rates were declined with increasing size resulting in a sigmoid growth curve (Fig. 7). Whereas Sufaylah attained a length of 14.4 mm within first year, its preadults (<40 mm SL) grew to around 10 mm year-1, individuals 40-50 mm SL grows only to about 4 mm year-1. Maximum sizes (>51 mm SL) were relatively few.
Reproduction
The results showed that the length at 50% maturity of H. pustulata from Northern coast of Aden Gulf varied within confined range, from 34 mm in males to 36 mm in females. General full sexual maturity achieved at 40-42 mm, smallest female with mature oocytes was 27 mm, with a body weight of 4.2 g. Males mature at a pit smaller size, the smallest mature male, with matured gonad had a shell length of 25.8 mm and a body weight of 3.8 g. Upon the females ranging between 27-62 mm in size (mean-42.05), about 62% had mature gonads. All the March-April samples of males and females who are larger than 36 mm had mature oocytes and were ready to spawn-IV-V stages of maturity.
According to present investigations Sufaylah are dioecious and lack any external evidences of sexual dimorphism. Gonads, especially their semblance, shape, features and color, can directly determine the sex of maturing specimens in early premature individuals. The ovarian tissue is dark green, some time with brownish shade; testis is usually beige, or yellow-cream.
 | |
| Fig. 7: | Growth curve and growth rate of H. pustulata |
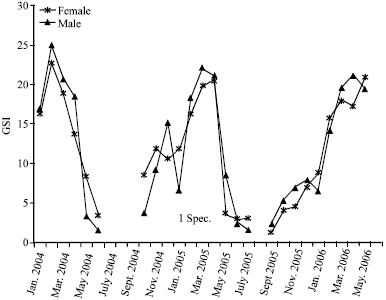 | |
| Fig. 8: | Seasonal variations of GSI of Aden Gulf H. pustulata |
Concerning the age of maturity, although it varied, but results revealed that 50% of males become mature in 2 years age, whereas females at third year.
Although, Sufaylah showed a typical progressive reproductive cycle, which seemingly terminates in almost synchronized spawning in mid spring (March and April) (Fig. 8); but frequently one can find gravid individuals in another months, especially in autumn. Gonadosomatic index (GSI) was apparently high, ranging between 10 and 25% in males and females, respectively. The highest GSI as well as the highest proportion of mature organisms prevailed from March to May (Fig. 8).The temperature of coastal water at these months varied between 25-28°C.
A result of monthly sex ratio data suggests that, the Sufaylah male ratios were almost less than 50%, except in April and May where males ratio is somewhat high (Fig. 9).
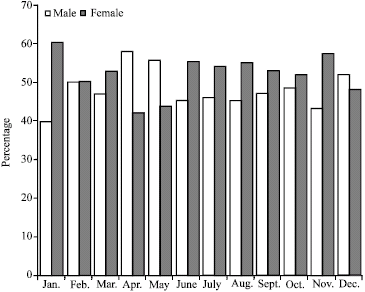 | |
| Fig. 9: | Seasonal sex ratio of H. pustulata from Aden Gulf Coast |
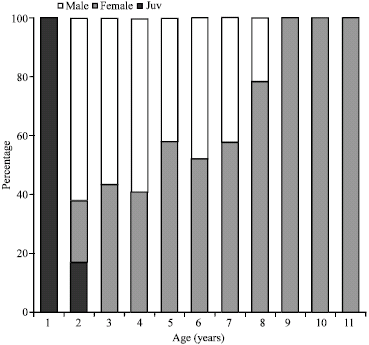 | |
| Fig. 10: | Age sex ratio of H. pustulata from Aden Gulf Coast |
As it is for age, while sex ratio of, firstly, matured Sufaylah, tends to support factual male preponderance; however, it seems as if more females were recorded at larger sizes and ages. Undoubtedly this can be explained by rarity of old ages specimens (Fig. 10).
Fecundity of Sufaylah in a Northern coast of Aden Gulf ranged from 77989 to 2,929,998 eggs for females of 36-62 mm shell length and 9-33 g total body weight (mean was 899771).
The variation in fecundity attains more than 1.250 million as soon as the individuals shifted from a shell length group of 40-45 to 50-55 mm group. Nevertheless, the number of matured eggs was increasing constantly with size. In spite of this trend, the relationship between fecundity and shell length was corresponding to a power relationship and expressed with relatively low coefficient-R2<0, 48, whereas with the body weight, the coefficient was higher-R2 = 0, 62 (Fig. 11).
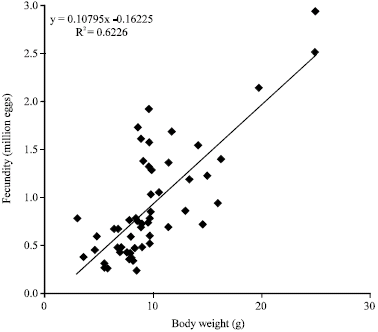 | |
| Fig. 11: | Bogy weight-fecundity relationship of H. pustulata from Aden Gulf Coast |
The relationship between age and fecundity was quite weak - R2 = 0, 28. Even so, the evident and relatively more reliable feature for fecundity evaluation in our Sufaylah is body weight, rather than shell length and of course age.
Additional data are still being collected to augment those already obtained. This will prove ecological characteristics and facilitate affirmation analyses. Actually, there is an ongoing assessment of the Sufaylah spawning cycle and behavior, growth and population evaluation on Aden Gulf coasts including Socotra Island.
DISCUSSION
Although, Haliotis sp. were thoroughly studied, commercially managed and farmed around the world, including neighboring Omani coast (Benny et al., 2003; Gordon and Cook, 2004; James et al., 2007; Hamasaki and Kitada, 2008) very little is known about it in Yemeni coast. It is still unexploited, in spite of the deduced here fact about its existence in large numbers.
From the study area investigation, it is found that most H. pustulata aggregations prevailing on Intertidal and adjacent zones of Hadhramout coast, which characterized by rocky shore intersected by sandy and somewhat gravel stripes and fringed by some hard coral complexities. This bottom, in addition to some seagrasses, especially well developed macroalgae assemblages occurred mainly in the study sites: Broom, Halla and Khalf of Mukalla and Shehir shorelines. Latter habitat was considered as a good provider of food and shelter for our subject to study-abalone (McShane, 1995; Day and Branch, 2000; Scheibling, 2006). As well as an intimate relationship between the organism presence and the moderate and big pink crusty boulders within rocky shore were prevalent. Such case is observed in all sites, where Sufaylah was found.
Spreading and Behavior
In general Sufaylah adopts the spreading pattern upon the different depths, that are the small sizes(juveniles), less than 25 mm are very rare or even absent in adult habitats, but frequently present in shallow depths <5 m, this phenomena was observed too in H. tuberculata (Campbell, 1996).
As survey shows, the pattern of restricted movement of Sufaylah intimately relates with the type and size of rocks and pink crusty boulders, under and around which it moves, mainly for feeding. Within such bottom assemblages, abalones are feeling safety and protected from predators; therefore it is moving within a very restricted area upon same site for prolonged periods, what corroborated by present of home scars in many of the samples sites. In the same sequence H. pustulata demonstrated an evident and promptly reaction against, not only searchlight, but even moderate moon light. These patterns of movement and behavior were observed in many other species, like H. rubra, H. tuberculata, H. cracherodii, H. rufescens and H. midae (Shepherd, 1973; Forster et al., 1982; Guzman-del-Proo, 1992; Tarr, 1995).
Observation also shows a strong relationship between Sufaylah and sea urchin, especially at day time and during nocturnal feeding, what may be due to similarities in the ecological niche. it can explained too as a type of disguise for abalone; in addition to the fact that sea urchin considers the main sharer of abalone by rasping the algae from rocky substrates (Tarr et al., 1996; Day and Branch, 2002).
Size, Age Structure and Growth
The population size structure using size-frequency data has often used to monitor the abalone stocks (Day and Fleming, 1992; Shephard and Brown, 1993; Siddeek and Johnson, 1997; Tarbath, 1999; Rogers-Bennett et al., 2004). This method could be of high value for providing baseline information, especially when it done for the first time, as present case.
From coupling the results of the size and age composition, it confirms that H. pustulata in Northern coast of Aden Gulf, naturally being substantially with relatively small size, as it is commonly known within distribution area-shell length were recorded up to 56 mm with average 25-35 mm (Geiger and Thacker, 2005). In present survey it is found little more -up to 62 mm, but its life span is relatively short-manly to 7-8 years. While this fact is corresponding with the general conclusion about predominance of small-bodied species in the Tropics, H. mariae from adjacent Omani coast which attains 122 mm and H. asinina from Thai waters and Eastern Australia 111 mm shell length, to some extent; transcend some large cold-water species (Keesing and Wells, 1989; Shepherd and Steinberg, 1992; McNamara and Johnson, 1995; Estes et al., 2005). However, H. pustulata from Aden Gulf is more large than Western Atlantic abalone H. pourtalesii 30 mm (Titgen and Bright, 1985) and slightly defer from some another small abalone species in other geographical areas like, H. diversicolor supertexta from Taiwan 85 mm and H. tuberculata 80 mm from the Macaronesian Archipelago (Peng et al., 1984; Clavier and Chardy, 1989). Although, present results are not comparable with big temperate zones Abalones, still, the latter species are much larger and their shell lengths can attain 200 mm - H. rufescens and 250 mm as it recorded for H. sorenseni and even more than 300 mm in H. rufescens California (Cox, 1962; Haaker et al., 1986; Karpov et al., 1999; Leighton, 1963).
Concerning the morphometric analyses conducted here, therefore can provide a useful baseline for reference in future studies with H. pustulata from Aden Gulf.
It seems from size and age structure of Sufaylah, that instead of the full absence of the artisanal and recreational fishery of abalone in a Yemeni coast, the older ages and big sizes tend to disappear quickly after a short matured life span. This case gives an impression of high natural mortality, which, can be reasoning of precocious aged (senility) in the species. This phenomenon often described for many species of abalone in other studied regions (Berthou et al., 1985; Shepherd and Breen, 1992); it was well observed in H. tuberculata in North-West of Mediterranean Sea (Mgaya, 1995). That is in addition to apparent effect of predatory by numerous discarded empty shells (Shepherd and Breen, 1992).
From many literatures, it becomes clear, that there is not full agreement about the validity of used methods of ageing and growth assessment in abalone (Prince et al., 1988; Day and Fleming, 1992; McShane and Smith, 1992; Wood and Buxton, 1997; Lee-Anne, 2006; Lee-Anne et al., 2008). Therefore, considering the research of many researcher on validity of growth bands or checks, undoubtedly we can confirm the present obtained results of Sufaylah age and growth rate (Poore, 1972; Nash et al., 1994; McNamara and Johnson, 1995; Haddon et al., 2008). Moreover, the deposition of the alternate annuli layers in the shells of Sufaylah is thought to be related to obvious low water temperatures of summer upwelling (June-August) and the just recently elapsed spawning the season-April (Fig. 8) (Peng et al., 1984; Shepherd et al., 1995; Leighton, 1963).
The growth rates of Sufaylah takes, with a little extent, the general nature of the rate decline: from more than 10 mm in first years, being about 2.4 mm year-1 in 7th year old; thereby it is not far from H. midae, H. discus, H. rubra and H. mariae (Hahn, 1989; Benny et al., 2003). This pattern is not varying from H. asinina that may grow 55.0 mm in one year (McNamara and Johnson, 1995); or from H. diversicolor from Taiwan which found to be at 1st and 2nd years old 39 and 56 mm, respectively (Peng et al., 1984). H. roei grow relatively quickly, attaining about 40 mm shell length during their first year, 60 mm in their second year, whereas average length of H. laevigata from Tasmania is only 55.6 at 4th year age (Keesing and Wells, 1989). Although, these indicates far higher growth rates than our results, in which pattern of growth markedly agreed with H. rubra from Tasmania (Hadden et al., 2008); H. cracherodii, from California; latter reach maximum shell diameters only about 20 mm in their first year and then grow at rates of 10-20 mm year-1 for the next several years (Leighton, 1963). Furthermore, the faster growth in first years of life is a biologically intuitive result, consistent, not only with other species of abalone, but with most of marine organisms (Tarrbath, 1999; Hancook, 2004).
Comparing present results with another Haliotis sp., disregarding the genetic size, it is found that the pattern and rate of the growth of Sufaylah from Aden Gulf, somewhat resembling and consistent with many of them.
The obtained in our results isometric growth of Sufaylah, which expressed by the equation of Shell length-weight relationship-W = 0.0001 SL3.1 was not far as for many other abalone species-for H. diversicolor supertexta from Taiwan-W = 0.0001 L2.9931; W = 0.0000521 L3.190 for H. rubra from Tasmania (Kent); for H. laevigata, from North West Tasmania was: W = 0.00006 L3.146; just as for H. laevigata from Western Australia-W = 0.0001 L3.07 (Peng et al., 1984; Officer et al., 2001; Tarrbath, 1999; Rogers-Bennett et al., 2007). It is obvious here that these relationships were close to a cubic relationship, as it in many regions.
Reproduction
Although visual identification of sex of immature Sufaylah less than 26 mm is not so difficult, however gonads of all individuals greater than 25 mm were identifiable, even they do not start spawning. This fact, with small size sexual maturity can observe in some tropical H. varia 17.3 mm from Thailand (Bussarawit et al., 1990), as well as temperate abalones (Campbell et al., 1992).
Whereas full sexual maturity in Sufaylah from Aden Gulf achieved at 40-42 mm, the shell length at 50% maturity ranged from 34 mm in males to 36 mm in females. This rang appeared to be very limited if compared with another species from other regions. H. kamtschatkana can mature at 31 mm shell length, but Campbell et al. (1992) found that the 50% matured at 55 mm and 100% individuals matured only at size 70 mm. These findings consistent with the results of maturity of South Australian abalones: H. cyclobates, H. laevigata, H. roei, H. rubber and H. scalaris (Shepherd and Laws, 1974); but far from present results. At the same manner H. rufescens from California reach sexual maturity at 1 25 mm, but many individuals become mature at about 41 mm, just like tropical abalone Haliotis asinine from Philippines, that matured at 40 mm shell length (Karpov et al., 1999).
Reproduction of many species of abalone is reported to occur at different times of the year in different coasts, whereas at others, spawning is to be epidemic, occurring within few days in spring or summer (Shepherd, 1986; Nash et al., 1994; Rodda et al., 1997; Litaay and De Silva, 2003). The finding that mature Sufaylah actually with a seasonal peak of spawning in April-May and may have ripe gonads year round agreed with majority of temperate and tropical species; for example H. kamtschatkana and H. varia (Campbell et al., 1992; Bussarawit et al., 1990). This fact implies that this species can spawn during other times of the year if environmental conditions, availability of food, are favorable (Shepherd and Laws, 1974; Campbell et al., 2003).
The results show some prevailing of females in monthly sex ratio, except in April and May where males is somewhat high (Fig. 9), hence the general sex ratio did not far from 1:1, what agree with most abalone species on other regions (Clavier, 1992; Capinpin et al., 1998; Litaay and De Silva, 2003); this may be coinciding with spawning emergence necessity when males were considered as the main motivator of spawning trigger in abalone at all (Cox, 1962; Clavier, 1992).
From the results, it was found that the fecundity is distinctively varied, what for abalone is quite usual (Wells and Keesing, 1989; Bussarawit et al., 1990; Clavier, 1992; Nash et al., 1994; Benny et al., 2003; Campbell et al., 2003).
The fecundity of H. pustulata as many H. sp. bear a positive but relatively weak linear relationship with body size parameters (Wells and Keesing, 1989; Litaay and De Silva, 2001, 2003).
From the present preliminary observation and result on H. pustulata from Northern Aden Gulf coast one can concludes that the untreated organism have an specific morphological, biological and ecological status slightly differs from other temperate and tropical species, it's life pattern and behavour in agreement with most of them. In spite of this, it requires more detailed studies, especially in growth, reproduction and feeding habits to provide a firm base for its forthcoming enhancements and farming.
ACKNOWLEDGMENT
We wish to thank anonymous scientists whose peer review of this article added to the quality and clarity of its content.
REFERENCES
- Awaji, M. and K. Hamano, 2004. Gonad formation, sex differentiation and gonad maturation processes in artificially produced juveniles of abalone, Haliotis discus hannai. Aquaculture, 239: 397-411.
Direct Link - Benny, A., A.L. Khalfan, A.R. Rashdi, A.L. Mashiki and M.A.L. Balker, 2003. Laboratory observation on the spawning, larval settlement and early growth of the Omani abalone Haliotis mariae wood. Proceedings of the National Seminar on Marine Biodiversity as a Source of Food and Medicine, September 26-28, 2003, SDMRI Res. Publ., pp:133-136.
- Campbell, A., J. Lessard and G.S. Jamieson, 2003. Fecundity and seasonal reproduction of northern abalone, Haliotis kamtschatkana, in Barkley Sound, Canada. J. Shellfish Res., 22: 811-818.
Direct Link - Day, E.G. and G.M. Branch, 2002. Influences of the sea urchin Parechinus angulosus (Leske) on the feeding behaviour and activity rhythms of juveniles of the South African abalone Haliotis midae Linn. J. Exp. Mar.Biol. Ecol., 276: 1-17.
CrossRefDirect Link - Estes, J., D.R. Lindberg and C. Wray, 2005. Evolution of large body size in abalones (Haliotis): patterns and implications. Paleobiology, 31: 591-606.
CrossRefDirect Link - Geiger, D.L. and C.E. Thacker, 2005. Molecular phylogeny of Vetigastropoda reveals non-monophyletic Scissurellidae, Trochoidea and Fissurelloidea. Molluscan Res., 25: 47-55.
Direct Link - Hamasaki, K. and S. Kitada, 2008. The enhancement of abalone stocks: Lessons from Japanese case studies. Fish Fisheries, 9: 243-260.
Direct Link - James, D.S., R.W. Day and S.A. Shepherd, 2007. Experimental Abalone ranching on artificial reef in port Phillip Bay, Victoria. J. Shellfish Res., 26: 687-695.
Direct Link - Leighton, D. and R.A. Boolootian., 1963. Diet and growth in the black abalone, Haliotis cracherodii. Ecology, 44: 228-238.
CrossRefDirect Link - Litaay, M. and S. De-Silva, 2001. Reproductive performance indices based on physical characteristics of female blacklip abalone Haliotis rubra L. J. Shellfish Res., 20: 673-677.
Direct Link - Litaay, M. and S. De-Silva, 2003. Spawning season, fecundity and proximate composition of the gonad of wild-caught blacklip abalone (Haliotis rubra) from Port Fairy waters, south eastern Australia. Aquat. Living Resour., 16: 353-361.
CrossRefDirect Link - McShane, P.E., 1995. Estimating the abundance of abalone: The importance of patch size. Mar. Freshwater Res., 46: 657-662.
CrossRefDirect Link - McShane, P.E. and M.G. Smith, 1992. Shell growth checks are unreliable indicators of age of the abalone Haliotis rubra (Mollusca: Gastropoda). Aust. J. Mar. Freshwater Res., 43: 1215-1219.
CrossRefDirect Link - Nash, W.J., T.L. Sellers, S.R. Talbot, A.J. Cawthorn and W.B. Ford, 1994. The population biology of abalone (Haliotis species) in Tasmania. I. Blacklip abalone (H. rubra) from the North Coast and the Islands of Bass Strait. Dept. Prim. Indust. Fish. Tasmania. Tech. Rep., 48. Sea Fisheries Division, Hobart, Tasmania 7001, Australia.
- Martin, G.G., K. Romer and C. Miller-Walker, 1983. Fine Structure of the Ovary in the Red Abalone Haliotis rufescens (Mollusca: Gastropoda). Zoomorphology, 103: 89-102.
CrossRef - Prince, J.D., T.L. Sellers, W.B. Ford and S.R. Talbot, 1988. A method for ageing the abalone Haliotis rubra (Mollusca: Gastropoda). Aust. J. Mar. Freshwater Res., 39: 167-175.
CrossRef - Rogers-Bennett, L., D.W. Rogers and S.A. Schultz, 2007. Modeling growth and mortality of red abalone (Haliotis rufescens) in Northern California. J. Shellfish Res., 26: 719-727.
Direct Link - Scheibling, R.E., 2006. Molluscan grazing and macroalgal zonation on a rocky Intertidal platform at Perth, Western Australia. Aust. Ecol., 19: 141-149.
CrossRefDirect Link - Shepherd, S.A., 1973. Studies on Southern Australian Abalone. (Genus Haliotis).1. Ecology of live Sympatric species. Aust. J. Mar, Freshwater. Res, 24: 217-257.
CrossRef - Shepherd, S.A., 1986. Studies on Southern Australian abalone (Genus Haliotis) VII. Aggregative behaviour of H. laevigata in relation to spawning. Mar. Biol., 90: 231-236.
CrossRef - Shephard, S.A. and L.D. Brown, 1993. What is an abalone stock: Implications for the role of refugia in conservation. Can. J. Fish. Aquat. Sci., 50: 2001-2009.
Direct Link - Shepherd, S.A. and H.M. Laws, 1974. Studies on southern Australian abalone (Genus Haliotis). II. Reproduction of five species. Aust. J. Mar. Freshwater Res., 25: 49-62.
CrossRef - Shepherd, S.A. and P.D. Steinberg, 1992. Food Preferences of Three Australian Abalone Species with a Review of the Food of Abalone. In: Abalone of the World: Biology, Fisheries and Culture, Shepherd, S.A., M.J. Tegner and S.A. Guzm�n-del-Pr�o (Eds.). Blackwell Scientific Publications Ltd., Oxford, ISBN: 978-0-85238-181-6.
- Shepherd, S.A., D. AL-Wahaibi and A.R. AL-Azri, 1995. Shell growth checks and growth of the Omani abalone Haliotis mariae. Mar. Freshwater Res., 46: 575-582.
CrossRefDirect Link - Siddeek, M.S. and D.W. Johnson, 1997. Growth parameter estimates for Omani abalone (Haliotis mariae, Wood 1828) using length-frequency data. Fisheries Res., 31: 169-188.
CrossRefDirect Link - Tarr, R.J.Q., 1995. Growth and movement of the South African abalone Haliotis midae: A reassessment. Mar. Freshw. Res., 46: 583-590.
CrossRefDirect Link - Tarr, R.J., P.V. Williams and A.J. Mackenzie, 1996. Abalone, sea urchins and rock lobster: a possible ecological shift that may affect traditional fisheries. S. Afr. J. Mar. Sci., 17: 319-323.
CrossRefDirect Link - Wells, F.E. and J.K. Keesing, 1989. Reproduction and feeding in the Abalone Haliotis roei Gray. Aust. J. Mar. Freshwater Res., 40: 187-197.
CrossRef - Wood, A.D and C.D. Buxton, 1997. Aspects of the biology of the abalone Haliotis midae (Linne 1758) on the East Coast of South Africa. 2. Reproduction. S. Afr. J. Mar. Sci., 17: 69-78.
Direct Link - Capinpin, E.C., V.C. Encena and N.C. Bayona, 1998. Studies on the reproductive biology of the donkeys Aquaculture, 166: 141-150.
Direct Link - Clavier, J., and P. Chardy, 1989. Investigation into the ecology of the ormer Haliotis tuberculata L. factors influencing spatial distribution. Aqut. Living Resour., 2: 191-197.
CrossRef - Mcnamara, D.C. and C.R. Johnson, 1995. Growth of the ass,s ear abalone Haliotisasinina Linne on heron reef tropical eastern Australia. Marine Freshwater Res., 46: 571-574.
Direct Link - Peng, S.C., K.T. Shao and K.H. Chang, 1984. Age and growth of small abalone H. diversicolor supertexta (Lischke) in hualien taiwan. Bull. Inst. Zool. Academia Sinica, 23: 29-38.
Direct Link - Lee-Anne, P., S. Kaehler and C.D. McQuaid, 2008. Using growth band autofluorescence to investigate large-scale variation in growth of the abalone Haliotis midae. Mar. Biol., 153: 789-796.
CrossRefDirect Link