
Hamid Ramezani
Yerevan State University, 1 Alex Manoogian Street, 0025, Yerevan, Armenia
Journal of Fisheries and Aquatic Science
Year: 2009 | Volume: 4 | Issue: 4 | Page No.: 203-209







ABSTRACT
The objective of this study is to assess the effects of protein and energy levels of diet on fish performance and growth efficiency of Caspian brown trout in order to develop optimum protein and energy level during the preparation of diet for this species. Fish were fed with six experimental diets containing three protein levels (45, 50 and 55%) and two energy levels (3.5 and 4 kcal g-1) according to a 3x2 factorial design. The diet was assigned to 18 tanks with 50 fish each, with three replicates for each diet. The experiment was conducted for 8 weeks with Caspian brown trout with an initial body weight of around 7 g. Protein content of diet influenced feed conversion ratio (p<0.05) and specific growth rate statistically (p = 0.052). Caspian brown trout demonstrated a better feed conversion ratio and a larger specific growth rate at lower protein levels (45 and 50%) in comparison to high protein level (55%). Protein efficiency ratio improved by a decrease in protein content of the diet (p<0.05), but energy content of diets does not affect any growth related parameter such as feed conversion ratio, specific growth rate and protein efficiency ratio. There was also no interaction between protein and energy levels in the growth related parameters, suggesting the effect of protein on the growth parameters in Caspian brown trout did not depend on energy levels of diet. In conclusion diet containing 50% protein can support the maximum growth and protein sparing by the use of high-energy diet did not occurred in this study.
PDF Abstract XML References Citation
How to cite this article
Hamid Ramezani, 2009. Effects of Different Protein and Energy Levels on Growth Performance of Caspian Brown Trout, Salmo trutta caspius (Kessler, 1877). Journal of Fisheries and Aquatic Science, 4: 203-209.
DOI: 10.3923/jfas.2009.203.209
URL: https://scialert.net/abstract/?doi=jfas.2009.203.209
DOI: 10.3923/jfas.2009.203.209
URL: https://scialert.net/abstract/?doi=jfas.2009.203.209
INTRODUCTION
The Caspian brown trout is one of the nine subspecies of brown trout, Salmo trutta caspius (Kessler, 1877), around the world (Quillet et al., 1992). This species is distributed mainly at the western and southern coast of Caspian Sea (Kiabi et al., 1999) and showed a better growth and higher weight gain compared to other brown trout which are living in Caspian Sea (Sedgwick, 1995). According to Berg (1964) Caspian brown trout entered to Caspian Sea from the Arctic region in the past. Due to its fillet quality, it has been known a commercially valuable fish in the southern cost of Caspian Sea.
The Caspian brown trout natural population has showed a steady decline over the past two decades and it has been characterized as endangered species in the southern part of Caspian Sea (Kiabi et al., 1999). River pollution, over-fishing, poaching and destruction of natural spawning sites are known as main reasons for such a sharp stock decline. There is no evidence showing that this species is able to do a successful reproduction in the rivers. Therefore, artificial reproduction has been introduced as a solution for enhancement and protection of wild populations.
In the restocking centers, Caspian brown trout larvae were raised in the concrete ponds until releasing weight of 20 g. Due to lack of specific production system for this species, rainbow trout production model has been applied to grow Caspian brown trout in the region. This condition caused some problems in restocking program such as a lowered growth rate and a high mortality of fingerlings.
Beside the stock enhancement program, there has been an attention toward artificial production of Caspian brown trout in Iran (Kalbassi et al., 2006). The aim is to introduce a new species into Iranian aquaculture industry. This species can be cultured in the cage or concrete pond along rainbow trout (Kalbassi et al., 2006). Understanding the requirements is a key step toward adjustment of this species to an artificial production system.
Preparation of a balanced diet and their adequate feeding is known as a key step for both restocking program and aquaculture. Nowadays, feed formulation for Caspian brown trout is based on information available for other salmonids. This solution however cannot be valid as different species show different requirement although they are classified within a family (Azevedo et al., 2004b, 2005).
Up to now, no nutritional studies have been conducted to produce artificial feeds for Caspian brown trout intensive culture. Growth, feed efficiency and body composition largely depends on protein and energy content in feed (Wang et al., 2006). Although dietary protein and energy requirements have been extensively studied in salmonids (Einen and Roem, 1997; Ruohonen et al., 1999; Lanari and Agaro, 2002; Azevedo et al., 2004a, b), those information for Caspian brown trout is still limited.
Therefore, the main goal of the study is to examine the effects of protein and energy levels of diet on fish performance and growth efficiency of Caspian brown trout. This data is needed to develop a proper diet to ensure the economical production of this species.
MATERIALS AND METHODS
Experimental Diets
In this study, three levels of dietary protein (45, 50 and 55%) and two levels of dietary energy (14.7 and 16.7 kJ g-1) were examined in a 3x2 factorial design. This led to six experimental diets which were formulated by replacing casein as a pure protein source with fat as an energy source in the diets. Ingredients selection and feed formulation were to meet nutrients requirements for brown trout. Protein and energy levels were adopted according to salmonids nutrients requirement range (Einen and Roem, 1997; Azevedo et al., 2004a).
The formulation and chemical composition of the six experimental diets are presented in Table 1 and 2, respectively. The chemical composition of the diets was different to examine the interactive effects of varying dietary protein and energy on growth performance of Caspian brown trout. Feed ingredients were provided from a local fish feed factory (Mazndaran Aqua-feed Company, Sari, Iran). The ingredients were separately ground and homogenously mixed. The pellet diets were made by the use of a cold feed pellet machine with a 2 mm diet. The resulting diets were then dried in an air dryer at 80°C and stored in a refrigerator until use.
| Table 1: | Percentage of ingredients used in the control diet and experimental diet |
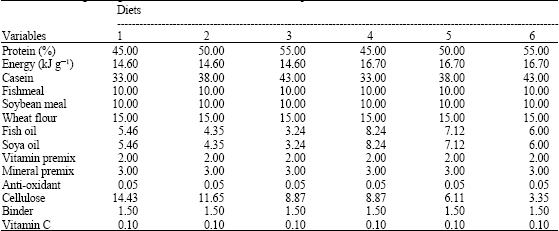 | |
| Table 2: | Nutrient composition of the experimental diets in % or kJ g-1 based on dry matter basis |
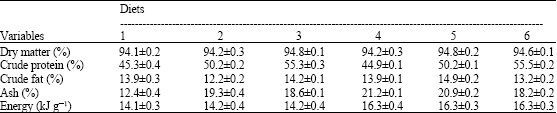 | |
Experimental System and Animals
The experiment was conducted for 8 weeks with Caspian brown trout with an initial body weight of 6-8 g during fall 2007. The fish were bred at the reproduction facility of the Shahid Bahonar, Kelardasht, Iran.
The fish were grown in the 18 fibre-glass tank (2x2x0.3 m), with a volume of 1200 L. The water supply was maintained by a channel connected to a nearby river. The photoperiod regime during the experiment was 12 h light and 12 h dark. Water quality was checked every week after the first feeding in the outflow of the tanks. The measured parameters were: temperature, pH, oxygen content, NH4+. The water flow for all tanks was equal around 14 l min throughout the experiment. Ammonium was measured in the tank which was received diet containing maximum protein level.
The water temperature was kept at 13-16 (14.3±1.2)°C during the experiment. Oxygen concentration was measured in a randomly selected tank by a digital oxygen detector and always remained above 8.5 (9.4±0.9) mg L-1. The water pH ranged between 7.45 to 7.62 (7.53±0.08) during the experiment. The ammonium concentration was always below 1 (0.33±0.11) mg L-1 during the experiment.
Experimental Procedure
On day one of the experiment, fish were randomly divided over 18 tanks (50 fish per tank). The fish were allowed to adapt to the experimental diets and to the feeding level for one week before the experimental period started.
The six diets were randomly assigned to 18 tanks with three tanks for each diet. The diets were given to the fish as soon as they were introduced into the tanks. The daily ration was calculated according to feeding chart adjusted for salmonids. The daily ration was divided into two feedings and fed by hand at 08:30 and 16:30. Before feeding, the pellets were sieved to remove dust and small particles. Fish were weighed at the beginning (day 0) and at the end of the experiment (day 56).
Chemical Analysis
Feed samples were collected at regular intervals (twice a week) during the experimental period and ground with a 1 mm screen before analysis. All chemical analysis were done in triplicate. Feed was analysed for dry matter by drying samples for 4 h at 103°C until constant weight (ISO, 1983). Ash content was determined by ashing the samples by incineration in a muffle furnace for 4 h at 550°C (ISO, 1978). Acid insoluble ash was measured by dissolving ash in hydrochloric acid (ISO, 1981). Crude protein (Nx6.25) was measured by the Kjeldahl method after acid digestion, according to ISO (1979). Lipid was extracted by petroleum ether extraction in a Soxhlet apparatus (ISO/DIS, 1996).
Calculations and Statistical Analysis
Fish Performance
Weight gain was determined by the difference between initial and final body weight. Specific Growth Rate (SGR) was calculated from the natural logarithm of mean final weight minus the natural logarithm of the mean initial weight and divided by the total number of experimental days expressed as a percentage (Amirkolaie et al., 2005).
SGR = 100 x (Ln Wfinal - Ln Winitial )/days |
The Feed Conversion Ratio (FCR) was calculated per tank from feed intake data and weight gain (Amirkolaie et al., 2005).
FCR = Feed intake/Wet weight gain |
Protein Efficiency Ratio (PER) was calculated by dividing the fish weight gain to total protein ingested during the experiment. Total protein ingested was estimated from the daily feed ration multiplied by diets protein content.
PER = Wet weight gain/Total protein ingested |
Statistical Analysis
Data are presented as means of each treatment with standard deviation. The data were verified for normality by the use of Kolmogorov-Smirnov test after transformation (ASIN). All data were analysed by a 2 way ANOVA for the effect of protein supplementation (45, 50 and 55%) and the effect of energy supplementation (3.5 and 4 kcal g-1). Protein levels and energy levels are the factors in this study. For all statistical analysis, tank was considered as the experimental unit. The means were compared by a Tukey’s post hoc test at five percent probability level.
RESULTS
Diet protein level influenced significantly FCR (p<0.05) in Caspian brown trout. Fish fed on diets containing 45 and 50% protein showed a better FCR than that of 55% protein. SGR is also affected by protein content of the diets (p = 0.052). Fish demonstrated a higher SGR by feeding on diet containing 45 and 50% protein.
Similar to FCR and SGR, Caspian brown trout showed a larger PER when fed on diets containing a lower level of protein (p<0.05; Table 3). Protein and energy levels did not influence final weight and weight gain statistically in Caspian brown trout, although feeding by diet containing 50% protein caused numerically a higher growth in fish.
In general, the energy levels did not affect the growth related parameters and fish showed similar performance at two experimental energy levels (Table 3). There was no interaction between protein and energy levels in the growth related parameters, suggesting that the effect of protein on growth parameters did not depend on energy levels of diet in Caspian brown trout.
| Table 3: | Growth performance±SD in brown trout fed with three levels of protein (45, 50 and 55%) and two levels of energy (3.5 and 4 kcal g-1) |
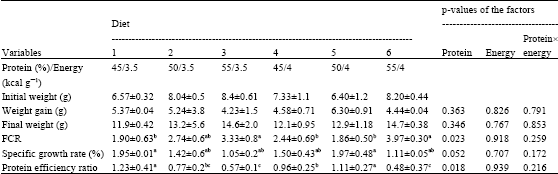 | |
DISCUSSION
In the current study, FCR and SGR of fish were improved as dietary protein level increased up to 50%. A better performance of FCR and SGR suggests that the optimum dietary protein level for Caspian brown trout to be between 45 and 50%. This finding is similar to the experiments in which different protein levels were evaluated in order to determine optimum protein requirement of salmonids. Based on those studies, protein requirement for salmonids vary between 40 and 50% (Webster and Lim, 2002; Lovell, 1989; Lee and Kim, 2001; Chan et al., 2002).
The reduction of growth at higher protein levels (55%) may be caused by catabolism of excessive amino acid by the fish. It has long been known that feeding protein in excess beyond requirement lead to catabolism of amino acid which associated with excretion of ammonia and loss of energy (Lloyd et al., 1978). In other words, since protein is the preferential energy source in fish metabolic routes (Hepher, 1988), brown trout utilize the extra protein for other metabolic functions not related to weight gain. This action cost a high energy loss in fish. Similar observation was made by Meyer and Fracalossi (2004) in Rhamdia quelen at high protein utilization level.
The main cause of larger energy loss is an increase in heat production followed by consumption of diet containing higher levels of protein. An increase in total energy expended on the ingestion, digestion, absorption and biochemical processing of a meal is main responsible for such a higher energy loss (Houlihan et al., 1995; Fu et al., 2007). Smitth et al. (1978) and Fu et al. (2005, 2007) showed that heat production associated with feeding protein to fish was larger than that for fat or carbohydrate. Therefore, high level protein diet may have increased energy cost of production, leading to a lower growth.
The reduction of growth performance may be also caused by poor water quality induced by high protein content diet. Amino acid degradation for energy supply increased ammonium excretion by the fish, leading to higher ammonia concentration in the water. Chakraborty and Chakraborty (1998) observed that ammonia loss by Labeo rohita was directly dependent on the amount of dietary protein. Ammonia build up in water is toxic to fish (El-Shafai et al., 2004) and may reduce the growth rate in the tanks at which fish were fed by diet containing maximum protein level (55%). Relatively higher ammonia concentration in the tank with the largest protein level (55%) may support the idea.
In the present study, surprisingly the growth related parameters were not affected with increase of dietary energy. This is in contrast to a number of earlier studies on different species including Lee and Kim (2001) in masu salmon ; Chan et al. (2002) in coho salmon; Meyer and Fracalossi (2004) in Rhamdia quelen; Wang et al. (2006) in cuneate drum; Salhi et al. (2004) in black catfish. There was also no interaction between dietary protein and energy suggesting that protein requirement dose not depends on energy content of the diets. This situation is also in contrast to the hypothesis that dietary protein requirements of fish are closely related to dietary energy levels and by proper use of non-protein energy sources, such as lipid and carbohydrate, dietary protein in fish feed can be spared (Shiau and Lan, 1996; Santinha et al., 1999; McGoogan and Gatlin, 2000). It appears that Caspian brown trout energy requirement was met by diet containing 3.5 kcal g-1 energy and extra energy does not lead to further growth. However, the idea can be debated by the fact that salmon generally rely more heavily on lipids as an energy source and that divert/spare more amino acids for protein deposition (Azevedo et al., 2004a). The opposite results gained in this study might be an issue that needs further investigation.
Protein efficiency ratio decreased with increasing dietary protein level. This is in agreement with the earlier findings for salmonids Lee and Kim (2001) in masu salmon and Chan et al. (2002) in coho salmon. The range of PER in this study was 0.48 to 1.23. This results are lower than the reported range of PER for coho salmon (1.8 to 2.4; Chan et al., 2002) and for masu salmon (1.1 to 1.8 ; Lee and Kim, 2001). This may support the idea that Caspian brown trout utilizes protein as main energy source. Like growth rate, energy content of the diet did not have an impact on PER. This may suggest that Caspian brown trout heavily depends on protein to supply energy for maintenance and growth. It appears that a high energy diet by the use of non-protein sources cannot spare a bigger protein portion for growth.
In conclusion, the addition of increasing protein level affected growth performance in Caspian brown trout and the best FCR and SGR were attained by diet containing 50% protein. An extra heat production related to the high protein diet may have reduced growth performance in this treatment. An increased dietary energy content of diet to spare the bulk of dietary protein for the growth appears not to work very well in the fish, as dietary energy dose not have any significant impact on growth, PER, FCR and SGR.
REFERENCES
- Azevedo, P.A., S. Leeson, C.Y. Cho and D.P. Bureau, 2004. Growth and feed utilization of large size rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar) reared in freshwater: Diet and species effects and responses over time. Aquacult. Nutr., 10: 410-411.
CrossRef - Azevedo, P.A., S. Leeson, C.Y. Cho and D.P. Bureau, 2004. Growth, nitrogen and energy utilization of juveniles from four Salmonid species: Diet, species and size effects. Aquaculture, 234: 393-414.
CrossRef - Azevedo, P.A., J. van Milgen, S. Leeson and D.P. Bureau, 2005. Comparing efficiency of metabolizable energy utilization by Rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar) using factorial and multivariate approaches. J. Anim. Sci., 83: 842-851.
PubMedDirect Link - Chan, J.C.K., J. Manu, B.J. Skura, M. Rowshandeli, N. Rowshandeli and D.A. Higgs, 2002. Effects of feeding diets containing various dietary protein and lipid ratios on the growth performance and pigmentation of post-juvenile Coho salmon Oncorhynchus kisutch reared in sea water. Aquacult. Res., 33: 1137-1156.
CrossRefDirect Link - Chakraborty, S.C. and S. Chakraborty, 1998. Effect of dietary protein level on excretion of ammonia in Indian major carp, Labeo rohita, fingerlings. Aquacult. Nutr., 4: 47-51.
CrossRefDirect Link - Einen, O. and A.J. Roem, 1997. Dietary protein/energy ratios for Atlantic salmon in relation to fish size: Growth, feed utilization and slaughter quality. Aquacult. Nutr., 3: 115-126.
CrossRef - El-Shafai, S.A., F.A. El-Gohary, F.A. Nasr, N.P. Van der Steen and H.J. Gijzen, 2004. Chronic ammonia toxicity to duckweed-fed tilapia (Oreochromis niloticus). Aquaculture, 232: 117-127.
CrossRef - Fu, S.J., X.J. Xie and Z.D. Cao, 2005. Effect of meal size on postprandial metabolic response in southern catfish (Silurus meridionalis). Comp. Biochem. Physiol., 140: 445-451.
CrossRef - Fu, S.J., Z.D. Cao and J.L. Peng, 2007. Effect of purified macronutrients on specific dynamic action in carnivorous Southern catfish. Aquacult. Nutr., 13: 216-221.
CrossRef - Houlihan, D.F., B.H. Pedersen, J.F. Steffensen and J. Brechin, 1995. Protein synthesis, growth and energetics in larval herring (Clupeaharengus) at different feeding regimes. Fish Physiol. Biochem., 14: 195-208.
CrossRef - Kalbassi, M.R., S. Dorafshan, T. Tavakolian, M. Khazab and H. Abdolhay, 2006. Karyological analysis of endangered Caspian salmon, Salmo trutta caspius (Kessler, 1877). Aquacult. Res., 37: 1341-1347.
CrossRef - Kiabi, B.H., A. Abdoli and M. Naderi, 1999. Status of the fish fauna in the South Caspian Basin of Iran. Zool. Middle East, 18: 57-65.
CrossRefDirect Link - Lee, S.M. and K.D. Kim, 2001. Effects of dietary protein and energy levels on the growth, protein utilization and body composition of juvenile Masu salmon (Oncorhynchus masou Brevoort. Aquacult. Res., 32: 39-45.
Direct Link - Lanari, D. and E. D’Agaro, 2002. Effects of changes in dietary DP/DE ratio during the growing phase on growth performance and carcass composition of rainbow trout (Oncorhynchus mykiss, Walbaum). Aquacult. Int., 10: 421-432.
CrossRef - McGoogan, B.B. and D.M. Gatlin, 2000. Dietary manipulations affecting growth and nitrogenous waste production of red drum, Sciaenops ocellatus. II. Effects of energy level and nutrient density at various feeding rates. Aquaculture, 182: 271-285.
CrossRef - Meyer, G. and D.M. Fracalossi, 2004. Protein requirement of jundia fingerlings, Rhamdia quelen at two dietary energy concentrations. Aquaculture, 240: 331-343.
CrossRef - Quillet, E., A. Faure, B. Chevassus, F. Krieg and Y. Harache et al., 1992. The potential of brown trout (Salmo truttaL.) for mariculture in temperate waters. ICEL Agric. Sci., 6: 63-72.
Direct Link - Ruohonen, K., J. Vielma and D.J. Grove, 1999. Low-protein supplement increases protein retention and reduces the amounts of nitrogen and phosphorus wasted by rainbow trout fed on low-fat herring. Aquacult. Nutr., 5: 83-91.
CrossRef - Salhi, M., M. G. Bessonart,Chediak and M.D. Bellagamba, Carnevia, 2004. Growth, feed utilization and body composition of black catfish, Rhamdia quelen, fry fed diets containing different protein and energy levels. Aquaculture, 231: 435-444.
CrossRef - Santinha, P.J.M., F. Medale, G. Corraze and E.F.S. Gomes, 1999. Effects of the dietary protein: Lipid ratio on growth and nutrient utilization in gilthead seabream (Sparus aurata L.). Aquacult. Nutr., 5: 147-156.
CrossRef - Shiau, S.Y. and C.W. Lan, 1996. Optimal dietary protein level and protein to energy ratio for growth of grouper (Epinephelus malabaricus). Aquaculture, 145: 259-266.
CrossRef - Smith, R.R., G.L. Rumsey and A.L. Scott, 1978. Heat increment associated with dietary protein, fat, carbohydrate and complete diets in salmonids: Comparative energetic efficiency. J. Nutr., 108: 1025-1032.
PubMed - Wang, Y., J.L. Guo, K. Li and D.P. Bureau, 2006. Effects of dietary protein and energy levels on growth, feed utilization and body composition of cuneate drum (Nibea miichthioides). Aquaculture, 252: 421-428.
CrossRefDirect Link - Amirkolaie, A.K., J.I. Leenhouwers, J.A.J. Verreth and J.W. Schrama, 2005. Type of dietary fibre (soluble versus insoluble) influences digestion, faeces characteristics and faecal waste production in Nile tilapia (Oreochromis niloticus L.). Aquacult. Res., 36: 1157-1166.
CrossRefDirect Link