
S. Velmurugan
Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
P. Soundarapandian N. John Samuel
Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
S. Rajagopal
Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502, Tamil Nadu, India
Journal of Fisheries and Aquatic Science
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 77-81







ABSTRACT
The role of hyperglycemic hormones in the eyestalk ablated and intact control crabs of Portunus pelagicus were studied in relation to various external stimuli. The eyestalk ablated crabs showed lesser free and reducing sugar levels and also lesser haemolymph glucose levels than the intact control crabs. The ablated crabs showed considerably higher glucose levels than the normal crabs when injected with eyestalk extract (CHH). However, the ablated crabs showed lesser glucose levels than the intact control crabs when exposed to chloroform stress. The polyacrylamide gel electrophoresis bands of crude eyestalk extracts and purified extracts showed a much similar pattern of banding and protein nature.
PDF Abstract XML References Citation
How to cite this article
S. Velmurugan, P. Soundarapandian N. John Samuel and S. Rajagopal, 2008. Activity of Hyperglycemic Hormone in the Eyestalk of Commercially Important Crab Portunus pelagicus (Linnaeus). Journal of Fisheries and Aquatic Science, 3: 77-81.
DOI: 10.3923/jfas.2008.77.81
URL: https://scialert.net/abstract/?doi=jfas.2008.77.81
DOI: 10.3923/jfas.2008.77.81
URL: https://scialert.net/abstract/?doi=jfas.2008.77.81
INTRODUCTION
In crustaceans, the X-organ /sinus gland complex typically located in the eyestalk plays a central role in physiological regulation of biological activities (Fingerman, 1987; Kellar, 1992). Hormones produced by this system are known to regulate reproduction, metabolism, osmoregulation, chromatic adaptation and growth. One of the best-known crustacean hormones with a relatively fast response time (between l and 2 h for maximum response) is the crustacean hyperglycemic hormone (Santos and Colares, 1986, 1990; Keller and Sedlmeier, 1988). The crustacean hyperglycemic hormone is involved in the regulation of hemolymph glucose, lipids and hepatopancreatic enzyme secretion. The crab, Portunus pelagicus is potent source of food and it has rich amount of nutrients. It is commercially important crab along Parangipettai coast and abundant throughout the year (John Samuel et al., 2004). Very limited study was carried out on the role of eyestalk hyperglycemic hormone in the hemolymph glucose levels of crabs in general and the crab, P. pelagicus in particular. Hence, the present study is aimed to estimate the total free and reducing sugars of crab’s haemolymph before and after eyestalk ablation. Hemolymph glucose response and chloroform stress on hemolymph glucose level were studied before and after eyestalk ablation in the crabs. Eyestalk-sinus gland protein separation was also performed by using PAGE method.
MATERIALS AND METHODS
Healthy crabs were brought to the laboratory from the Annankoil landing center of Parangipettai (Lat. 11°29’ N and Long. 79°46’ E) and acclimatized in the laboratory conditions (Temp. 28±2°C; Salinity 35±2 ppt; Do 6.5 ppm; pH 8.2±0.5). Total free and reducing sugars were determined in intact control crabs. Subsequently unilateral eyestalk ablation was performed in the same animals by cutting the left eyestalk at its base with fine, clean scissors and the wound was cauterized immediately with a hot blunt needle in order to prevent the loss of hemolymph and mortality (Caillouet, 1973). Hemolymph was collected by hypodermic syringe (27 gauze needle) from the eyestalk-ablated animals every 2 h and total free and reducing sugars were determined.
Total free sugars estimated by anthrone method (Roe, 1955). Reducing sugars were determined as per Nelson (1944) and Somogyi (1945).Glucose was estimated in both groups of animals by oxidase peroxidase method.
Eyestalk extract was prepared by grinding 4 eyestalks with 0.5 mL of distilled water in a mortar and a small amount of clean sea sand. This extract was centrifuged for 5 min to settle the coarse detritus. The supernatant was separated and used for the experiment. The experimental animals were divided into two group’s one group with eyestalk and other group without eyestalk. The initial hemolymph was collected from the intact control and eyestalk ablated crabs and determines the glucose level by the glucose oxidase method. After that 0.1 mL of the crude eyestalk was injected to both set of experimental animals and allowed to stand for sometime. Every 2 h haemolymph was collected both intact control and eyestalk ablated crabs and the glucose was estimated as earlier.
The animals are divided into two groups. One group served as intact control and other group was eyestalk-ablated animals. Before exposing the animal to chloroform stress the hemolymph samples were taken and glucose was estimated. Subsequently animals were exposed to chloroform stress and haemolymph was collected every 2 h intervals for the estimation of glucose-by-glucose oxidase method.
Eyestalk sinus gland protein was separated by adopting one dimensional Sodium Dodecyl Sulphate (SDS) Poly Acrylamide Gel Electrophoresis (PAGE) (Laemmli, 1970).
RESULTS
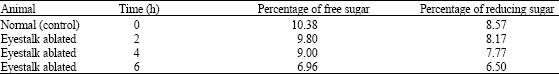
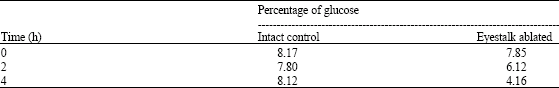
The results of the total free and reducing sugars are presented in Table 1. The percentage of total free sugar was maximum (10.38%) before eyestalk ablation and decreased gradually for the first 4 h. The value decreased drastically (6.96%) after 6 h. The percentage of total reducing sugar was maximum (8.57%) before the eyestalk ablation and decreased gradually for the first 4 h. The value decreased drastically (6.50%) after 6 h. The percentage of the glucose level decreased in eyestalk ablated animals rather than control animals (Table 2).
In both the groups, an increase in the glucose level was noticed for the first four hours after the eyestalk extract was injected. The next two hours showed a gradual decrease in the levels of glucose in animals of both the group. The eyestalk ablated animals showed higher variation in the glucose levels (maximum of 16% and minimum of 6.57%) when compared with the intact control animals (Table 3).
| Table 1: | The percentage of the total free and reducing sugars at various hours after eyestalk ablation |
 | |
| Table 2: | The percentage of glucose level in intact control and eyestalk ablated animals |
 | |
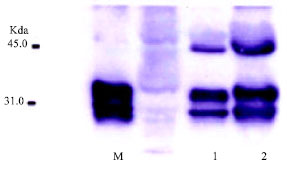 | |
| Fig. 1: | The polyacrylamide gel plate |
| Table 3: | The percentages of the glucose level in animals after eyestalk extract injection |
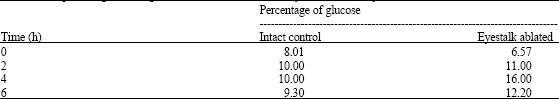 | |
| Table 4: | The percentage of the glucose level after chloroform stress |
 | |
When exposed to chloroform stress, the percentage of the glucose level decreased slightly in the eyestalk ablated animals (maximum 6.7% and minimum 6.41%) when compared with normal intact control animal (maximum 10.4% and minimum 6.57%) (Table 4).
Three bands appeared in both the samples, which were more or less similar by polyacrylamide gel electrophoresis (Fig. 1). The bands of the purified sample were thinner and clearer, while those of the crude extract were broader and not clearly demarcated.
DISCUSSION
Carbohydrates in the tissue of crustaceans exist as free sugars and as bound with proteins (Saravanan, 2000). The free sugars in hemolymph consist of mono, di and oligosaccharides. All monosaccharides, maltose and its oligosaccharides constitute total reducing sugars. Trehalose constitutes the non-reducing sugar fraction of the total free sugars.
In the present study the eyestalk ablation of the crab leads to decrease in the glucose level in the hemolymph compared to the intact control animals, which maintain their own glucose level. In this experiment every 2 h hemolymph was taken during the first 2 h interval glucose level slightly decreases, after 2 h interval hemolymph glucose level more rapidly decreases. The percentage of total reducing and total free sugars were also decreased in eyestalk-ablated animals. Eyestalk extirpation is a classical operation of crustacean endocrinology; it removes the X-organ sinus gland complex, which is the source of an array of hormones, including the hyperglycemic hormone. Removal of eyestalks eliminates the hyperglycemic hormone from circulation, which results in significant decrease in hemolymph sugar level in Oziotelphusa senex senex (Sreenivasulu Reedy, 1999). This clearly implies that the eyestalk sinus gland produce hyperglycemic hormones that maintain the levels of haemolymph sugar.
The eyestalk extract injection increases the level of glucose in haemolymph of both intact control and eyestalk-ablated animals. The eyestalk extract when injected into the eyestalk-ablated crab drastically increased the haemolymph sugar levels. This is due to the lack of CHH, which is essential in maintaining the hyperglycemic condition. When injected the eyestalk extract with CHH, these animals showed a sudden increase of haemolymph glucose levels. The possible reason for this sudden increase might be that the abrupt incorporation of CHH into the animal might have triggered the hepatopancreas to release larger amounts of glucose into the blood. Further, the eyestalk-ablation and the process of injection itself are stress factors on the crab, which would have caused the animal to secrete glucose when supplied with CHH. In the normal animals, since CHH was already maintaining the glucose level of the hemolymph, the excess addition of the eyestalk extract (CHH) might not have produced any significant effect. When injected with eyestalk extracts (CHH), the ablated crabs showed much higher blood glucose level, which could lead to decreased animal growth. This further substantiates the theory that eyestalk ablation of culture crabs could lead to accelerated growth and consequentially better yield.
Chloroform stress rapidly increases the glucose level of the hemolymph in intact control animals, but it decreases slightly in the eyestalk-ablated animals. When under stress, the metabolic activities of the animals increase rapidly, causing the hepatopancreas to release higher levels of glucose into the hemolymph. The sinus gland produces hyperglycemic hormone (CHH), which maintains the hyperglycemic level. An experiment conducted on manual and eyestalk crustaceans (Santos and Keller, 1993) confirmed the hormonal influence on glycemia. Webster (1996) made similar observations in the crab Cancer pagurus. He recorded significantly higher hemolymph glucose levels in the normal animals and it was decrease slightly in the eyestalk-ablated animals. In the normal animals, higher glucose levels were recorded. This may be due to hyperglycemia, which is maintained by CHH, while in the eyestalk-ablated animals, lack of CHH could be the reason, why the animal showed lesser glucose levels in the hemolymph. Similar results as in the present study have been obtained in C. pagurus by Santos and Colares (1990). They stated that the level of glucose was found to increase rapidly in the hemolymph following emersion stress in, there by rapidly changing the CHH level.
The polyacrylamide gel electrophoresis bands of the extracts clearly indicate a protein nature and since the extracts were taken from a neurohemal organ (eyestalk), one of the three bands maybe CHH.
REFERENCES
- Caillouet, C.W. Jr., 1973. Ovarian maturation by eyestalk ablation in pink shrimp Penaeus duorarum Burkenroad. Proc. World Maricult. Soc., 3: 205-211.
CrossRef - Samuel, J.N., N. Thirunavukkarasu, P. Soundarapandian, A. Shanmugam and Kannupandi, 2004. Fishery potential of commercially important portunid crabs along parangipettai coast. Proceedings of the International Conference and Exposition on Marine Living Resources of India for Food and Medicine, February 27-29, 2004, Aquaculture Foundation of India, Chennai, pp: 165-173.
- Kellar, R., 1992. Crustacean neuropeptides: Structures, functions and comparative aspects. Experientia, 48: 439-445.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Roe, J.H., 1955. The determination of sugar in blood and spinal fluid with anthrone reagent. J. Biol. Chem., 212: 335-343.
PubMedDirect Link - Santos, E.A. and E.P. Colares, 1986. Blood glucose regulation in an intertidal crab, Chasmagnathus granulata (Dana, 1851). Comp. Biochem. Physiol., 83: 673-675.
CrossRef - Santos, E.A. and E.P. Colares, 1990. Blood glucose changes in the blue crab Callinectes sapidus Rathbun on transfer from seawater to air. Brazil J. Med. Boil. Res., 23: 333-335.
PubMed - Santos, E.A. and R. Keller, 1993. Regulation of circulating levels of the Crustacean hyperglycemic hormone-evidence of a dual feedback control system. J. Com. Physiol., 163: 374-379.
CrossRef - Webster, S.G., 1996. Measurement of crustacean hyperglycemic hormone (CHH) level in the edible crab, Cancer pagurus, during emersion stress. J. Exp. Biol., 199: 1579-1785.
PubMedDirect Link