
K. Sivashanthini
Department of Zoology, University of Jaffna, Jaffna, Sri Lanka
G.A. Charles
Department of Zoology, University of Jaffna, Jaffna, Sri Lanka
S. Shutharshan
Department of Zoology, University of Jaffna, Jaffna, Sri Lanka
Journal of Fisheries and Aquatic Science
Year: 2008 | Volume: 3 | Issue: 5 | Page No.: 320-327







ABSTRACT
The present study on fecundity of Gerres abbreviatus, Bleeker, 1850 (Pisces: Gerreidae) occurring in Jaffna lagoon, Sri Lanka covers annual fecundity, batch fecundity, spawning frequency and relationship between fecundity and various lengths of G. abbreviatus. The composite histograms based on the diameters of all oocytes at successive stages of maturation showed three modes in ovaries at stages 4, 5 and 6 in G. abbreviatus. Fecundity varied from 167610 to 3162280 ova for an animal with a total length of 149 to 280 mm. Annual Fecundity (AF) varied significantly between various size groups and showed perfect positive correlation with total length (TL, mm) and total weight (W, g). The relationships between length or weight and fecundity were statistically significant and best described with the simple linear regressions: AF = 19.678 TL3.563 and AF = 218.2 TW1.226. Linear regressions for logarithmic transformed data of annual fecundity and batch fecundity versus total length were computed. Comparison of these two regressions confirmed that G. abbreviatus spawn around three batches of eggs during a season.
PDF Abstract XML References Citation
How to cite this article
K. Sivashanthini, G.A. Charles and S. Shutharshan, 2008. Fecundity Studies of Gerres abbreviatus (Bleeker, 1850) From the Jaffna Lagoon, Sri Lanka. Journal of Fisheries and Aquatic Science, 3: 320-327.
DOI: 10.3923/jfas.2008.320.327
URL: https://scialert.net/abstract/?doi=jfas.2008.320.327
DOI: 10.3923/jfas.2008.320.327
URL: https://scialert.net/abstract/?doi=jfas.2008.320.327
INTRODUCTION
Fecundity has been considered as the number of ripening eggs in the female prior to spawning (Bagenal and Braum, 1978; Hunter and Macewicz, 1985a, b). It varies from species to species, depending on age, length, weight, environmental conditions, length, age etc. Many workers have worked on the fecundity of different fish, viz., Almatar et al. (2004), Alam et al. (1994), Bhuiyan et al. (1993), Bhuiyan et al. (2000), Bhuiyan and Parveen (1998), Dadzie et al. (2000), Kiran and Puttaiah (2003), Kuddus et al. (1996) and Macchi et al. (2004). Studies on fecundity estimates are staples of fishery science. Inevitably, they will be estimated for every species of economic consequence because of their importance in the dynamics of the population (Bagenal, 1967; King, 1995). Another reason for studying fecundity is that when fecundity estimates are combined with estimates of the abundance of eggs in the sea, they can be used to estimate the biomass of a stock (Hunter et al., 1992).
Fecundity can be defined by several terms such as annual fecundity, total fecundity, potential annual fecundity, determinate fecundity, indeterminate fecundity, batch fecundity and relative fecundity (Hunter et al., 1992). Annual fecundity is the total number of eggs spawned by a female per year; Total fecundity signifies the standing stock of advanced yolk oocytes; Potential annual fecundity is the total advanced yolked oocytes matured per year, uncorrected for atretic losses (Hunter et al., 1992). In species, with determinate fecundity, potential annual fecundity is considered to be equivalent to the total fecundity prior to the onset of spawning. Determinate fecundity refers to annual fecundity when the potential annual fecundity becomes fixed prior to the onset of spawning. In fishes, with determinate fecundity total fecundity decreases with each spawning because the standing stock of advanced yolked oocytes is not replaced during the spawning season. Annual fecundity is indeterminate when the potential annual fecundity of a female is not fixed prior to the onset of spawning and unyolked oocytes continue to be matured and spawned during the spawning season. Batch fecundity is the number of hydrated oocytes released in one spawning, usually determined by counting the number of hydrated oocytes in the ovary. Relative fecundity can be obtained by dividing the fecundity by female weight (Hunter et al., 1992).
There is no information for studies on fecundity of G. abbreviatus. Consequently, the present study was undertaken to find out annual fecundity, batch fecundity, spawning frequency and relationship between fecundity and various lengths of G. abbreviatus.
MATERIALS AND METHODS
Random samples of G. abbreviatus were collected from Pasaioor, Kakkaitivu, Kurunagar and Navanthurai landing centres (Fig. 1) during March 2007-February 2008. At least one sample was obtained every week from each landing center. Since the sex of these fishes cannot be differentiated externally, the fishes were brought to the laboratory and the abdomens of the fishes were cut open to expose the gonads for the identification of the sexes. For each fish, total length TL (mm) was measured to the nearest 1 mm using a measuring board and total weight TW (g) was weighed to the nearest 1.0 g by an electronic balance. Nearly, all immature and mature gonads were examined macroscopically and microscopically.
Oocyte development pattern (synchronous, group synchronous or asynchronous) and type of annual fecundity (indeterminate or determinate) were assessed by oocyte size-frequency distributions (Hunter and Macewicz, 1985a) and histology (Hunter and Macewicz, 1985a; West, 1990). Six fishes were selected for analysis of oocyte size-frequency distributions; 3 fish in October and another three in February, representing gonad development early and late in the spawning season. Diameter of nearly 500 oocytes from each stage of ovary was measured using an ocular lens and stage micrometer.
 | |
| Fig. 1: | Map showing the collection sites of Jaffna lagoon, Sri Lanka, (a) Kakkaithevu, (b) Navanthrai, (c) Kurunagar, (d) Pasaioor, (e) Thondaimannar lagoon and (f) Jaffna lagoon |
All fecundity estimates were based on fish that had undamaged ovaries and showed no sign of previous spawning in that season (i.e., no loose, hydrated oocytes in the lumen of the ovary, Watson et al., 1992), no sign of post ovulatory follicles and no sign of major atresia. Initially, 1 g portions from five of these fish were dissected from the anterior, median and posterior regions of the gonad and weighed accurately (±0.001 g). Analysis of Variance (ANOVA) was used to compare the number of oocytes per gram between sub samples along the ovaries (in the anterior, median and posterior regions). Because, no significant differences (p>0.05) were observed between regions the medial gonad portions were weighed accurately and used to estimate fecundity by the gravimetric method (Hunter and Macewicz, 1985b) from the following relationship:
Annual fecundity was estimated from yolked oocytes (stages 4 and 5) from samples collected during spawning season. Batch fecundity, where fish may spawn more than one batch of eggs within a spawning season, was estimated from counts of hydrated oocytes in the nine suitable stage 6 ovaries. Only females that had stage 4 ovaries without post ovulatory follicles as confirmed through histological analysis were selected for batch fecundity analysis. The average relative fecundity was measured as the number of oocytes per gram body weight.
The annual fecundity was related to the total length, total weight, ovary weight or ovary length of fishes by using the following relationship (Bagenal, 1967):
F = a Xb |
where, F is the fecundity, a is a constant, b is the exponent derived from the data and X is the total length, total weight, age, ovary weight or ovary length of the fish. The following logarithmic transformation was used to obtain the regression lines of each relationship:
Log F = log a+b log X |
Predicted regression lines and 95% confidence limits for the regression lines for log fecundity against the log of each independent variable were obtained by simple linear regression residual analysis.
Simple linear regressions were used to describe relationships between log annual fecundity-log total length, log batch fecundity-log total length, log annual fecundity-log total weight, log annual fecundity-log ovary weight and log annual fecundity-log ovary length. The average relative fecundity was measured as the number of oocytes g-1 b.wt. and logarithm of it was regressed against log total length and log total weight. The correlation coefficient (r) was calculated (Snedecor and Cochran, 1967) to find out the relationship between each variable.
RESULTS
A total of 999 specimens of G. abbreviatus (442 males and 557 females) were collected from commercial catches of medium siragu valai, gillnet and beach seine from the Jaffna lagoon. The size (Total length-TL) of male G. abbreviatus ranged from 102 to 280 mm TL and females ranged from 97 to 279 mm TL.
Macroscopic and microscopic studies of female gonads revealed seven oocyte developmental stages such as chromatin nuclear stage, perinucleolar stage, cortical alveoli stage, yolk stage, nuclear migration stage, hydration stage and postovulatory follicle stage (POF) based on the occurrence of the most advanced type of oocytes present, regardless of their abundance.
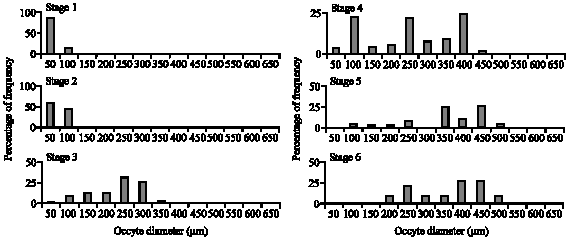 | |
| Fig. 2: | Frequency distribution of oocyte diameter (μm) of G. abbreviatus for stage 1 to 6 ovaries |
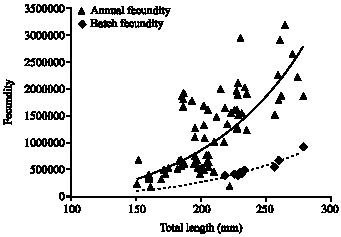 | |
| Fig. 3: | Relationship of annual fecundity and batch fecundity to total length of G. abbreviatus |
Size frequencies of the oocytes of G. abbreviatus in different stages of ovarian development are shown in Fig. 2. Ova diameter in G. abbreviatus ranged between 13.6 and 532.6 μm. The composite histograms (Fig. 2) based on the diameters of all oocytes found in the six females at successive stages of maturation showed three modes in ovarian stages 4, 5 and 6 of G. abbreviatus. These suggested that this fish spawn asynchronously in three batches. In the present study, a continuous oocyte distribution, i.e., lack of hiatus between advanced yolked oocytes and less mature oocytes and abundance of yolked oocytes not decreasing over the spawning season (Hunter and Macewicz, 1985a; Hunter et al., 1992) observed and so indeterminate fecundity is expected in gerreids.
Annual Fecundity (AF) increased exponentially with Total Length (TL) (Fig. 3), ranging from 167610 (149 mm TL) to 3162280 (280 mm TL). Annual fecundity varied considerably for a given length, linear regression of the log-transformed data (Fig. 3) was statistically significant (F1,73 = 83.1, p<0.001, R2 = 0.535) yielding the equation:
log[AF] = 3.563xlog TL+1.294 |
Similarly, Batch Fecundity (BF) ranged from 367818 (219 mm TL) to 837588 (279 mm TL) in 9 fishes and the regression for the log-transformed data was (Fig. 3), Comparison of these two regressions revealed that around 3 batches of eggs were spawned each year.
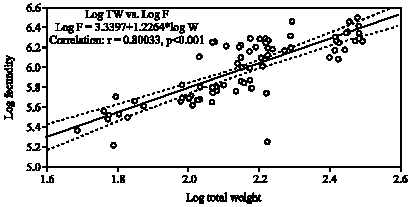 | |
| Fig. 4: | Relationship between log annual fecundity and log total weight of G. abbreviatus. *Solid line is the predicted log annual fecundity at log total weight, dotted lines show upper and lower 95% confidence estimates of the predicted line |
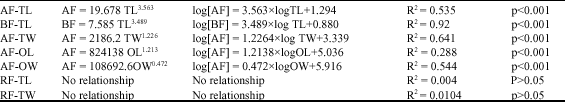
| Table 1: | Relationships between fecundity and other independent parameters of G. abbreviatus |
 | |
log[BF] = 3.489xlog TL + 0.880 (F1,8 = 83.76, p<0.001, R2 = 0.92) |
The linear regression of log-transformed fecundity versus log-weight (Fig. 4) was statistically significant (F1,73 = 128.29, p<0.001, R2 = 0.641) yielding the equation:
log[AF] = 1.226xlog TW+3.339 |
The relative fecundity varied between 601 and 15,570 oocytes g-1 and the average was 7272 oocytes (±3197 SD) g-1 (total weight). Relative fecundity (AF/TW) did not increase significantly with fish length (F1,73 = 0.2988, p>0.05, R2 = 0.004) or weight (F1,73 = 0.756, p>0.05, R2 = 0.0104). Parabolic and logarithmic equations of G. abbreviatus with R2 and p-values of above relationships are shown in Table 1. Equations for relationships of fecundity-ovary weight and fecundity-ovary length and the relevant parameters are also shown in Table 1.
DISCUSSION
The high mean relative fecundity of G. abbreviatus (7272 oocytes g-1 total weight) signifies the importance of managing this species along the coastal waters of Sri Lanka. The annual fecundity of G. abbreviatus increases exponentially (167610-3162280 oocytes per fish between 149 and 280 mm total length) with the increase in the length and age of females. These oocytes are released in about three large batches of 367818-837588 oocytes per fish, increasing with length of females.
As, the postovulatory follicles usually persist in fish ovaries for only a short period (Hunter and Goldberg, 1980; Macewicz and Hunter, 1993; Melo, 1994), the co-occurrence of postovulatory follicles and ripe oocytes in G. abbreviatus ovaries suggests that successive batches of oocytes may be released within a few days. More intensive sampling over short time periods within the spawning season would be required to prove this.
The exponential value is usually reported as 3 when fecundity is related to length and 1 when fecundity is related to weight. However the values may vary from 2.3 to 5.3 for a great variety of fishes (Bagenal and Braum, 1978). In the present study, the exponential value is greater than cube (b = 3.563, R2 = 0.535) when fecundity is related to total length and also greater than 1 (b = 1.2264, R2 = 0.641) when fecundity is related to weight. According to Bagenal and Braum (1978) changes in the environment such as temperature, salinity and oxygen may also result in significant changes in fecundity. Wootton (1973) suggested that food supply and nutrition affect the egg production and thereby the fecundity. He confirmed in his experiments with stickle back, that the low food intake led to fewer eggs, greater weight at maturity and shorter inter-spawning interval.
Prabhakara Rao (1970) observed that the fecundity of G. oyena varied from 104211 to 1443785 eggs in the size ranges of 148-282 mm. The exponent values obtained for length-fecundity and weight-fecundity relationships of G. oyena were 2.43 and 1.872, respectively. The correlation co-efficient between fecundity-length and fecundity-weight were reported as 0.5306 and 0.4421, respectively. Similarly, Patnaik (1971) observed that the fecundity of G. setifer varied from 17293 to 161505 eggs in the size ranges 88-193 mm. For G. filamentosus, Kurup and Samuel (1991) reported fecundity to vary from 64278 to 387576 oocytes per fish in the size ranges of 100-148 mm standard length and the exponential values for length-fecundity and weight-fecundity relationships of G. filamentosus were 3.2563 and 0.9896, respectively with correlation coefficients of 0.7178 and 0.6906. On comparison with other gerreids, it is clear that the fecundity obtained in the present study for G. abbreviatus is relatively higher than that of G. oyena, G. setifer and G. filamentosus.
Various authors have tried unsuccessfully to relate absolute fecundity and age, e.g., Simpson (1951) and Raitt and Hall (1967). Lear (1970) found that fecundity was proportional to age to the power 1.95 in the halibut, Reinhardtius hippoglossoides. Similarly May (1967) found fecundity to be nearly proportional to the square of the age in cod Gadus morhua.
The weight of the ovaries of a fish is mainly influenced by the number of ova contained in them. Many workers reported the increase in fecundity with increasing ovary weight (Bagenal, 1967).
CONCLUSION
High mean relative fecundity was obtained for G. abbreviatus. The annual fecundity of G. abbreviatus increased exponentially with the increase in the length. In the present study eggs are released in about three large batches of oocytes. Exponential value of 3.563 was obtained when fecundity is related to total length in the present study. Fecundity of different stocks of the same species can be used for racial discrimination studies. Different races have characteristic fecundities and egg size so that the racial origins of fishes can be determined from egg counts and a population can be identified as a homogenous unit or as a mixture of different stocks.
ACKNOWLEDGMENT
The authors are grateful for the authorities of University of Jaffna for the financial assistance and facilities provided.
REFERENCES
- Almatar, S.M., K.P. Lone, T.S. Abu-Rezq and A.A. Yousef, 2004. Spawning frequency, fecundity, egg weight and spawning type of silver pomfret, in Kuwait waters. J. Applied Ichthyol., 20: 176-188.
CrossRefDirect Link - Dadzie, S., F. Abou-Seedo and T. Al-Shallal, 2000. Reproductive biology of the silver pomfret, Pampus argenteus (Euphrasen), in Kuwait waters. J. Applied Ichthyol., 16: 247-253.
CrossRefDirect Link - Hunter, J.R. and S.R. Goldberg, 1980. Spawning incidence and batch fecundity in Northern Anchovy, Engraulis ordax. Fish. Bull., 77: 641-652.
Direct Link - Hunter, J.R. and B.J. Macewicz, 1985. Rates of atresia in the ovary of captive and wild Northern Anchovy, Engraulis mordax. Fish. Bull., 83: 119-136.
Direct Link - Hunter, J.R., B.J. Macewicz, N.C. Lo and C.A. Kimbrell, 1992. Fecundity, spawning and maturity of female Dover sole Microstomus pacificus, with an evaluation of assumptions and precision. Fish. Bull. US., 90: 101-128.
Direct Link - Macchi, G.J., P. Marcelo and E. Martín, 2004. Seasonal egg production pattern of the Patagonian stock of Argentine hake (Merluccius hubbsi). Fish. Res., 67: 25-38.
CrossRef - May, A.W., 1967. Fecundity of Atlantic cod. J. Fish. Res. Board Can., 24: 1531-1551.
CrossRefDirect Link - Melo, Y.C., 1994. Multiple spawning of the anchovy Engraulis capensis. South Afr. J. Mar. Sc., 14: 313-319.
Direct Link - Raitt, D.F.S. and W.B. Hall, 1967. On fecundity of the redfish Sebastes marinus. J. Cons. Perm. Int. Explor. Mer., 31: 237-245.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1967. Statistical Methods. 6th Edn., Oxford and IBH Publishing Co. Pvt. Ltd., New Delhi, India, Pages: 593.
Direct Link - Watson, J.J., I.G. Priede, P.R. Witthames and A. Owori-Wadunde, 1992. Batch fecundity of Atlantic mackerel, Scomber scomberus L. J. Fish Biol., 40: 591-598.
CrossRefDirect Link - West, G., 1990. Methods of assessing ovarian development in fishes: A review. Aust. J. Mar. Freshwater Res., 41: 199-222.
CrossRefDirect Link - Wootton, R.J., 1973. The effect of size of food ration on egg production in the female three spined-stickleback Gasterosteus aculeatus L. J. Fish. Biol., 5: 89-96.
CrossRefDirect Link