
Magdy A. Alwany
Department of Marine Science, Faculty of Sciences, Suez Canal University, Egypt
Michael Stachowitsch
Department of Marine Biology, Faculty of Life Sciences, University of Vienna, Austria
Mahmoud H. Hanafy
Department of Marine Science, Faculty of Sciences, Suez Canal University, Egypt
Ali A.F.A. Gab-Alla
Department of Marine Science, Faculty of Sciences, Suez Canal University, Egypt
Journal of Fisheries and Aquatic Science
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 38-46







ABSTRACT
The food preferences of the highly specialized butterflyfish Chaetodon melannotus were investigated on the reef flat in the Northern Red Sea, Egypt. The food items include nine species of soft corals (belonging to 7 genera, Sinularia, Litophyton, Nephthea, Sarcophyton, Heteroxenia, Lobophytum and Parerythropodium), one species of stolonifera (Tubipora musica), hard substrate and other minor items (including the hard coral genus Pocillipora and soft coral genus Cladiella). In general, the total average feeding rate of C. melannotus was 40.6± 7.6 bites (5 min)‾1. The two most strongly selected genera, Sinularia and Litophyton, represented 78.0% of the total food items: Sinularia [22.6 bites (5 min)‾1, 55.7%], Litophyton [9.0 bites (5 min)‾1, 22.3%]. Based on Ivlev’s electivity index, C. melannotus prefers Heteroxenia fuscescens over other soft corals in the Northern Red Sea and tends to feed less on others such as Sarcophyton ehrenbergi and Lobophytum pauciflorum.
PDF Abstract XML References Citation
How to cite this article
Magdy A. Alwany, Michael Stachowitsch, Mahmoud H. Hanafy and Ali A.F.A. Gab-Alla, 2007. Food Preferences of the Butterflyfish Chaetodon melannotus on the Reef Flat in Sharm El-Sheikh, Egypt. Journal of Fisheries and Aquatic Science, 2: 38-46.
DOI: 10.3923/jfas.2007.38.46
URL: https://scialert.net/abstract/?doi=jfas.2007.38.46
DOI: 10.3923/jfas.2007.38.46
URL: https://scialert.net/abstract/?doi=jfas.2007.38.46
INTRODUCTION
Many fish groups are known to feed directly on living corals, either biting off the polyps, scraping the coral surface or cutting off the tips of coral branches (Harmelin-Vivien, 1979). The most important group is butterflyfishes (Chaetodontidae). This diverse, specialized group includes approximately 114 species (10 genera) distributed circumtropically (Burgess, 1978). While many butterflyfishes exhibit distinct preferences among their coral food resources, the causal factors behind these preference patterns are poorly understood (Hourigan et al., 1988). Considerable information is available on the feeding habits of butterflyfishes in the Indo-Pacific (Reese, 1975, 1977; Neudecker, 1977, 1979; Ralston, 1981; Harmelin-Vivien and Bouchon-Navaro, 1981, 1983; Sano et al., 1984; Tricas, 1985; Cox, 1986; Motta, 1988; Ferry-Graham et al., 2001a, b) and the Caribbean (Randall, 1967; Lasker, 1985; Neudecker, 1985). Relatively few feeding ecology studies are available for the Red Sea (Harmelin-Vivien and Bouchon-Navaro, 1981; Righton, 1997; Zekeria et al., 2002; Alwany, 1997, 2003).
The highly specialized predatory blackback butterflyfish Chaetodon melannotus (Bloch and Schneider, 1801) feeds on octocorals, which are toxic or unpalatable to many other predators and initially locates its prey using visual rather than chemical cues (Alino et al., 1992). It usually inhabits depths between 0.5-25 m and defends exclusive feeding territories against conspecifics. Territory size is related to group size and averages 500± 293 m2 (Righton, 1997). Because the diet of C. melannotus is very specialized, its habitat occupation may be more variable than in other butterflyfish species if required food resources are patchily distributed. C. melannotus is also the most aggressive butterflyfish species in the Northern Red Sea (M. Alwany, pers. observation).
Alwany et al. (2003) were the first to define the feeding selectivity of two corallivorous butterflyfishes (Chaetodon austriacus and C. trifascialis) in the Northern Red Sea. The aim of this complementary study is to focus in more detail on the food preferences and feeding behaviour of the more highly specialized congener C. melannotus on Northern Red Sea fringing reef flats, Egypt. Four sites were chosen to include the full range of structures on the reef flat and all potential food items; possible differences between the sites were tested with ANOVA.
MATERIALS AND METHODS
Preliminary observations to broadly determine the range of food items of Chaetodon melannotus were followed by a total of 120 h of detailed observations on 64 C. melannotus individuals by snorkeling on the reef flat. Data were collected over a four-week period in May and June 2004.
Study Area
The research was conducted at the fringing reefs off Sharm El-Sheikh city, Red Sea, Egypt (Fig. 1). Within this area, only the reef flat was examined; four sites were chosen to represent the full range of reef flat habitats and potential food items.
Site 1 (27° 53' N-34° 19' E): off Tower Hotel of Sharm El-Sheikh city, often calm. Reef flat between 10-30 m wide, only few patches of hard and soft corals, densely covered by algae at the reef edge. Site 2 (27° 47' N-34° 17' E): known as Faresh Ghzlany, in front of the Visitor Center of Ras Mohammed National Park, sheltered and calm.
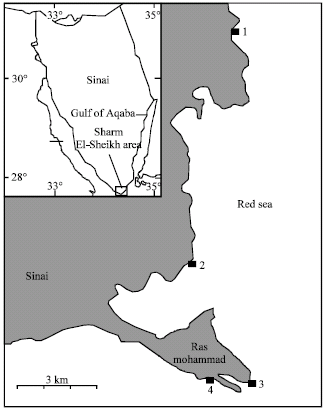 | |
| Fig. 1: | Map of Northern Red Sea showing location of the four study sites |
Reef flat relatively poor in coral communities, extending over a width of 40-60 m. Site 3 (27° 43' N-34° 15' E): Yolanda Reef, at the southern tip of the Ras Mohammed area, strong waver action year-round. Reef flat very broad (60 to 300 m) and rich in coral colonies. Site 4 (27° 44' N-34° 14' E): Old Quay, often calm, but more turbid. Reef flat relatively wide (35 m) and rich in coral colonies.
Feeding Rates and Selectivity
The feeding rates of 64 Chaetodon melannotus were measured as the biting action of fish per unit time (feeding rate = number of bites per minute). Randomly selected, mature fishes of the same size and condition were observed by snorkeling at a distance of 1-3 m. The observer swam slowly to minimize disturbance. Bites per food item were counted for six consecutive periods of 5 min for each individual (Reese, 1975, 1977). For the average density of C. melannotus and other species, see Alwany (1997). Shifting the observations along the reef every day avoided recording the same individual twice. Foraging behaviour was recorded during daytime from 1000 to 1500. Ivlev’s electivity index (Ivlev, 1961) was used to measure feeding selectivity, comparing the utilization of food with respect to its availability. This index has been successfully used to determine prey selectivity in a wide range of marine and freshwater fishes (Kim, 1991; Ushakumari and Aravindan, 1992). Ivlev’s electivity index is calculated as in Alwany et al. (2003).
Coral Percentage Cover and Identification
Coral cover was calculated within a 1 m2 quadrat divided into 16 small (25χ 25 cm) squares with a total of 25 points. Eight quadrats were laid down haphazardly on the reef flat at each site. The percentage coral cover for each species was calculated from its proportion of the total counts in each quadrat. When possible, species level was recorded directly in situ; in other cases, higher taxa were recorded and a small fragment of each soft coral was collected, labeled and kept in 70% alcohol for further identification in the lab. Soft coral specimens of uncertain identification were positively identified by comparison with Indo-Pacific shallow-water soft corals deposited at the Natural History Museum in London (NHML).
Data Analysis
The data were analyzed statistically using PRIMER (V. 5.0) and SPSS (V. 11.5). To compare coral diversity among different sites, three diversity indices were calculated based on the percentage cover of corals: species richness D (Margalef, 1968); Shannon-Wiener H� [Loge] (Shannon and Weaver, 1949); Pielou’s evenness J� (Pielou, 1969). ANOVAs were performed on raw data to compare feeding rates. For coral cover, data were log-transformed (x+1): the assumption of homogeneous variances was met (and ANOVAs calculated); for two species (Sarcophyton ehrenbergi and Lobophytum pauciflorum) non-parametric ANOVAs (Kruskal-Wallis test) were performed.
RESULTS
Feeding Activities
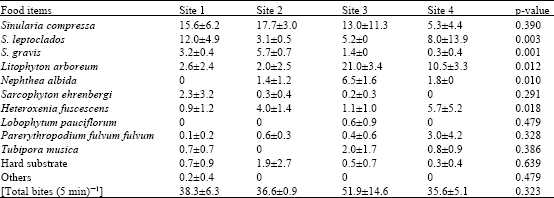
Overall, the list of the different food items (Table 1) includes nine species of soft corals (belonging to 7 genera: Sinularia, Litophyton, Nephthea, Sarcophyton, Heteroxenia, Lobophytum and Parerythropodium), one species of stolonifera (Tubipora musica), hard substrate and other minor items (includes the hard coral genus Pocillipora and the soft coral genus Cladiella). Overall, the average feeding rate of C. melannotus was 40.6± 7.6 bites (5 min)‾1. Two genera (Sinularia and Litophyton) were clearly eaten most and represented 78.0% of the total food items. Sinularia ranked first [22.6 bites (5 min)‾1, representing 55.7%] and Litophyton second [9.0 bites (5 min)‾1, representing 22.3%].
| Table 1: | Feeding rates (mean± SD) of C. melannotus at the four study sites |
 | |
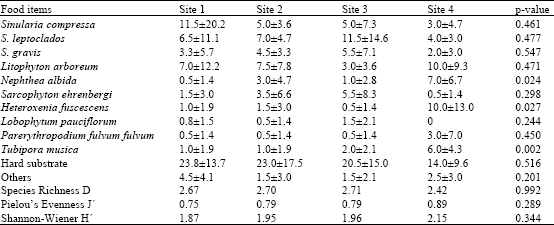
| Table 2: | Percentage cover of food items and diversity indices at the four study sites |
 | |
Heteroxenia fuscescens and Nephthea albida represented 7.2 and 6.0%, respectively. None of the other categories exceeded 2.5%, i.e., not more than one bite per 5 min. The feeding rates of C. melannotus did not differ significantly between sites (one-way ANOVA, p = 0.323).
Coral Cover
Sinularia and Litophyton are the most abundant soft coral genera at the study area, with a combined average percentage cover of 40.1% (28.6 and 11.5, respectively). In general, Sinularia leptoclados was the most abundant representative, followed by L. arboretum, S. compressa and S. gravis. The percentage of hard substrate amounted to 33.8% and this category encompassed a wide range of invertebrate communities and algae. Despite the different site conditions, one-way ANOVA showed that almost all coral and substrate categories did not differ significantly between sites. The exceptions were the two soft corals H. fuscescens (p = 0.027) and N. albida (p = 0.024) as well as T. musica (p = 0.002). The diversity indices (D, J' and H') at the four sites are given in Table 2. The highest richness was recorded at site 3 (2.71), the lowest at site 4 (2.42). Average evenness ranged from 0.75 at site 1 to 0.89 at site 4, while average Shannon-Wiener varied between 1.87 at site 1 and 2.15 at site 4. The three diversity indices did not differ significantly between sites.
Selectivity
Chaetodon melannotus fed on 12 food items, which can be divided into three broad categories ranging from preference to avoidance. The first category (>0.3) includes Sinularia compressa, Heteroxenia fuscescens, S. leptoclados and Litophyton arboretum, which are eaten more by C. melannotus. The second category (<0.3 to -0.3) includes Parerythropodium fulvum fulvum, S. gravis, Nephthea albida and Tubipora musica, which are characterized by no selection.
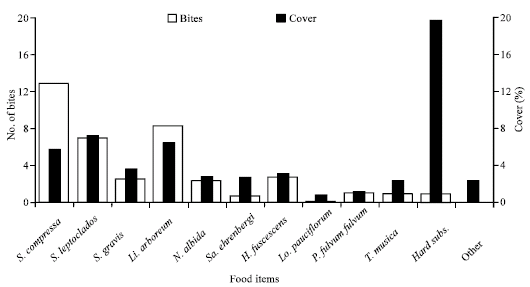 | |
| Fig. 2: | Average number of bites (5 min)‾1 on food items by C. melannotus and% cover of these items |
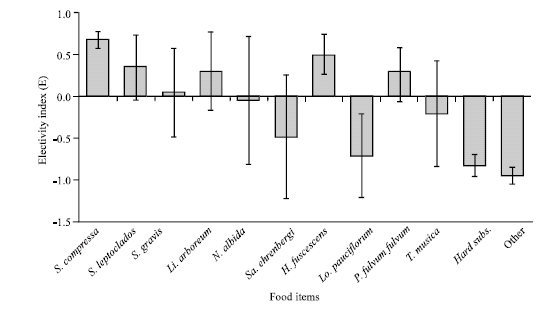 | |
| Fig. 3: | The average electivity indices (E) of C. melannotus on different food items in the study area |
The third category (<-0.3) includes Sarcophyton ehrenbergi, Lobophytum pauciflorum, hard substrate and other minor items (Pocillipora and Cladialla), which tend to be eaten less.
Sinularia compressa, L. arboreum and S. leptoclados had the highest number of bites (Fig. 2). Nonetheless, Chaetodon melannotus consumed more H. fuscescens than L. arboreum and S. leptoclados. This fish tended to bite Sarcophyton ehrenbergi less frequently, despite a relative cover nearly equal to that of H. fuscescens. C. melannotus showed clear patterns of preference and avoidance (Fig. 3). Although C. melannotus fed much more on S. compressa and H. fuscescens, the percentage cover of the former was high and that of the latter relatively low. Thus, the butterflyfish apparently prefers H. fuscescens over other soft corals in the Northern Red Sea. In contrast, it fed less on S. ehrenbergi and Lobophytum pauciflorum.
DISCUSSION
Reef octocorals are eaten by only a few species of specialized invertebrates and fishes (Sammarco and Coll, 1988). Such species may play an important role in overall coral community structure, considering that soft corals are key competitors with hard corals. Chaetodon melannotus is a well-known example along the East African coast (Talbot, 1965), in the Marshall Islands (Reese, 1977), in Madagascar (Harmelin-Vivien, 1979), in the Red Sea (Harmelin-Vivien and Bouchon-Navaro, 1981; Righton, 1997), in Japan (Sano et al., 1984b; Sano, 1989) and in Australia (Alino, 1989; Alino et al., 1988, 1992).
Feeding Rates
Alwany et al. (2003) examined the feeding rates of a generalist hard coral feeder, Chaetodon austriacus and a specialist hard coral feeder, C. trifascialis, in the Northern Red Sea, Egypt. The average feeding rates of C. austriacus (30.2 bites 5 min‾1) and of C. trifascialis (37.2 bites 5 min‾1) were similar to but slightly lower than that of C. melannotus in the present study (40.6 bites 5 min‾1). This would tend to support the interpretation that as fish become more specialized for a certain kind of food, their feeding rates increase (Alwany et al., 2003). One explanation would be that special or restricted food may not always contain all the nutritional components normally found in a broader variety of food items. C. melannotus appeared to be an energy maximizer because it fed at a relatively high rate (one bite every 7-8 sec).
Food Preferences
Sano (1989) reported that Chaetodon melannotus fed on alcyonarians (92%), unidentified animal material (5%), scleractinians (2%), sea anemones or hexcorallia (1%) and less than 1% on filamentous algae. Our results confirm Sano’s results rather than those of Harmelin-Vivien and Bouchon-Navaro (1981). In the present study, C. melannotus fed on alcyonarians (95.7%, including 7 genera), stoloniferans (0.9%, only a single species), hard substrate (0.8%, including filamentous algae) and other minor food items (0.1%, including the hard coral genus Pocillipora and the soft coral Cladiella). In the Northern Red Sea, Righton (1997) reported that C. melannotus preferred the genus Sinularia because this soft coral was the most common on the reef. Our results confirmed that Sinularia as most common on the Sharm El-Sheikh reef flats (28.6%), followed by Litophyton (11.5%), but more precisely identifies the actual preference for Heteroxenia.
Most butterflyfishes feed selectively on corals with high energy contents (Tricas, 1985; Alwany, 2003). The importance of lipids in coral mucus was examined by Benson and Muscatine (1974), who found wax esters and triglyceride to be the major lipid components exuded. Tricas (1989) confirmed that coral lipid content was very important for Hawaiian butterflyfishes, thus pointing to factors other than prey abundance to explain preference and avoidance. Ingestion of coral mucus by reef fishes is one route by which the energy-rich products of coral metabolism may be transferred to the reef fish population (Benson and Muscatine, 1974; Wild et al., 2004). Food selection, however, can depend on a number of different factors (such as the relative toxicities of many octocoral toxins; Coll et al., 1982). This may explain why C. melannotus preferred Sinularia over Sarcophyton: Coll et al. (1982) reported that Sarcophyton was the most toxic genus and Lobophytum, Sinularia and Nephthea ranged from highly toxic to non-toxic. The fecal analysis of the soft coral feeder Chaetodon unimaculatus on Guam (Pacific Ocean) indicated that the terpenoid metabolites in Sinularia extracts are passed through the fish digestive system without any apparent breakdown or conversion to other metabolites (Wylie and Paul, 1989). Alino (1989) demonstrated that C. melannotus metabolized diterpenes and was thus apparently capable of processing large quantities of these potentially toxic compounds. Our results therefore confirm Alino’s finding that food choice need not be based solely on avoidance of specific toxins.
CONCLUSIONS
Marine fishes, even so-called feeding specialists, can have a varied prey that does not necessarily reflect the composition of potentially available items. In the Egyptian Red Sea, the blackback butterflyfish Chaetodon melannotus mainly fed on 9 species of soft coral and the organ-pipe coral Tubipora musica. Although two soft coral genera Sinularia and Litophyton were most abundant in the reef flat and also represented most of the food items, this butterflyfish clearly preferred another, less abundant soft coral genus, Heteroxenia.
ACKNOWLEDGMENTS
This research would not have been possible without the kind assistance of the Department of Marine Science (Suez Canal University, Egypt) and the University of Vienna, Austria. The authors wish to thank Prof. Joerg Ott, Department of Marine Biology at the University of Vienna in Austria, for providing working space. The authors wish to thank Dr. Tarek Temraz, Department of Marine Science at Suez Canal University in Egypt, for identification of soft corals. Finally, the authors would like to thank the authorities of the Ras Mohamed National Park for logistical support during field work; special thanks are expressed to Mr. Belal Saleh (Ras Mohamed National Park, Sharm El-Sheikh, Egypt).
REFERENCES
- Alwany, M.A., E. Thaler and M. Stachowitsch, 2003. Food selection in two corallivorous butterflyfishes, Chaetodon austriacus and C. trifascialis, in the Northern Red Sea. Marine Ecol., 24: 165-177.
Direct Link - Ferry-Graham, L.A., P.C. Wainwright, C.D. Hulsey and D.R. Bellwood, 2001. Evolution and mechanics of long jaws in butterflyfishes (family Chaetodontidae). J. Morphol., 245: 120-143.
Direct Link - Ferry-Graham, L.A., P.C. Wainwright and D.R. Bellwood, 2001. Prey capture in long-jawed butterflyfishes (Chaetodontidae): The functional basis of novel feeding habits. J. Exp. Marine Biol. Ecol., 256: 167-184.
Direct Link - Pielou, E.C., 1969. An Introduction to Mathematical Ecology. 1st Edn., Wiley-Interscience, New York, USA, ISBN: 0-471-68918-1, Pages: 286.
Direct Link - Randall, J.E., 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr., 5: 665-847.
Direct Link - Wild, C., M. Huettel, A. Klueter, S.G. Kremb, M. Rasheed and B.B. Jorgensen, 2004. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature, 428: 66-70.
Direct Link - Zekeria, Z.A., Y. Dawit, S. Ghebremedhin, M. Naser and J.J. Videler, 2002. Resource partitioning among four butterflyfish species in the Red Sea. Marine Freshwater Res., 53: 163-168.
Direct Link